過表達EuCuZnSOD提高巨尾桉的耐寒性研究
2018-05-30 14:42:42趙艷玲韓瑤靳玉蕾
廣西植物 2018年1期
趙艷玲 韓瑤 靳玉蕾
摘 要:? 銅鋅超氧化物歧化酶(CuZnSOD)是一種清除超氧陰離子自由基的金屬酶,與多種抗逆性相關。該研究利用CaMV35S啟動子融合巨尾桉EuCuZnSOD1基因通過根癌農桿菌介導的葉盤轉化法導入巨尾桉中,經卡那霉素初步篩選和PCR鑒定,得到10株轉基因陽性植株。結果表明:通過-13.1 ℃暗處理5 h的抗寒性實驗,發現40和41號植株在低溫下無葉片萎蔫現象,組培室培養100 d后全株存活無白化葉片,說明這兩株轉基因巨尾桉的抗低溫和凍害后恢復能力較強。在4 ℃低溫脅迫下跟蹤監測轉基因溫室苗的SOD酶活性發現,與對照相比轉基因植株的SOD酶比活力無規律性且變化不大,但是相同條件下實時定量PCR檢測結果表明抗寒性較強的40和41號植株在常溫下EuCuZnSOD1的表達量較對照增加了約9倍,4 ℃處理36 h后EuCuZnSOD1的表達量相比對照植株分別增加了16倍和36倍,說明EuCuZnSOD1在低溫脅迫下的表達量增加提高了巨尾桉的耐寒性。分析轉基因組培苗全株的硫酸木質素含量,表明EuCuZnSOD1高表達不影響甚至降低了巨尾桉的木質素含量,40號植株的木質素含量與對照相同,而41號整株植株的木質素含量相比對照降低了56%,其他檢測轉基因植株的木質素含量均有不同程度的降低。該研究結果表明巨尾桉中大量表達EuCuZnSOD1基因可以提高巨尾桉的抗寒能力以及寒害后的恢復能力,不影響甚至可能是負調控了轉基因巨尾桉的木質素生物合成。
關鍵詞: 巨尾桉, 銅鋅超氧化物歧化酶, 轉基因, 抗寒性, 硫酸木質素
中圖分類號:? Q945.78
文獻標識碼:? A
文章編號:? 1000-3142(2018)01-0101-08
Over-expression of cytosolic copper/zinc superoxide dismutase gene and increase cold-tolerance in Eucalyptus grandis × E. ophylla
ZHAO Yanling, HAN Yao, JIN Yulei
( College of Chemical Engineering, Huaqiao University, Xiamen 361021, Fujian, China )
Abstract:? Copper/zinc superoxide dismutases are important antioxidant enzymes to guard cells against superoxide toxicity. The over-expression of CuZnSOD can improve the resistance and recovery ability of plants to abiotic stresses such as salt, drought, cold and high temperature stress. The plant genome encodes three types of CuZnSOD, the CuZnSOD1 in cytoplasm, the CuZnSOD2 in chloroplast and the CuZnSOD3 in peroxisome or extracellular. The cytosolic EuCuZnSOD1 gene was cloned from Eucalyptus grandis × E. ophylla (GenBank Accession Number: JX138573) and the biological function was verified by prokaryotic expression system. The fusion vector of sense EuCuZnSOD1 gene derived by CaMV35S promoter was constructed and transformed to Eucalyptus grandis × E. ophylla by Agrobacterium. Using Kanamycin selection and PCR analysis, ten transgenic eucalyptus lines were obtained. Compared with the control plants the tissue culture transgenic eucalyptus No. 40 and No. 41 could be well tolerant in -13.1 ℃ for 5 h and recovery without obvious damage. There was no distinct increase in SOD enzyme activity in transgenic plant paralleled with wild plant under room temperature and 4 ℃ for 48 h. However, the real-time PCR analysis showed that the expression of? EuCuZnSOD1 was raised to about nine times higher than wild line under room temperature. After 4 ℃ for 36 h, the expression of EuCuZnSOD1 in No. 40 and No. 41 line was respectively increased about 16 and 36 times higher than the wild plant and then down to the normal level at 4 ℃ for 48 h. The data indicated that the increased expression of EuCuZnSOD1 improves the cold tolerance of Eucalyptus. The klason lignin content of transgenic Eucalyptus was reduced especially the No. 41 Eucalyptus was decreased to 56% of the control plant except for the No. 40 plant was no significantly different from the control plant. In conclusion, over-expression of the cytosolic EuCuZnSOD1 in Eucalyptus grandis × E. ophylla could enhance the cold-tolerance and improve the recovery ablity.
Key words: Eucalyptus grandis × E. ophylla, copper/zinc-superoxide dismutases, transgenosis, cold-tolerance, klason lignin
桉樹是世界三大用材樹種之一,其種植面積已經占世界人工林面積的三分之一,是一種極具價值的經濟林。但是,低溫、其它環境脅迫、人工除草等高成本因素嚴重影響了桉樹人工林的發展,而選育良種、培育新品種則是解決這一問題的有效途徑。基因工程育種具有高效性和目標性強等特點,可以加速優質、高抗桉樹新品種的選育進程。目前,已經有澳大利亞、日本、巴西和中國等獲得了轉基因桉樹。
目前,桉樹抗寒分子機理研究較少,主要集中在抗寒轉錄因子CBF的功能研究,首先,法國圖盧茲大學的Teulieres課題組克隆了巨桉的CBF基因(Kayal et al,2006),研究了CBF基因與抗寒性的關系(Navarro et al,2009),將巨桉CBF1基因轉入雜交桉樹中發現轉基因桉樹除了抗寒能力增強之外,還表現出生長緩慢、保水能力增強等常青闊葉樹所具備的生理特點(Navarro et al,2011);隨后分析了AP2/ERF基因家族的功能,發現DREB1/CBF與桉樹的寒害逆境相關(Cao et al,2015),后通過高通量的qPCR技術研究了巨桉的CBF和DREB2與桉樹冷、熱和干旱逆境的關系,發現CBF調節桉樹的抗逆與生長(Nguyen et al,2017)。將巨桉的低溫脅迫響應基因EgrCR(徐鳳華等,2016)轉化擬南芥,提高了擬南芥的抗凍性,證明了與桉樹的抗寒性相關。此外,還開展了低溫脅迫下的尾巨桉和鄧恩桉轉錄組學研究(Liu et al,2014)和巨桉的莖在短時寒害下的蛋白質組學研究(Leonardi et al,2015)。
SOD活性可作為篩選桉樹抗寒性的一個生理指標,研究發現不同種類的桉樹應對低溫逆境時其SOD的活性變化差異明顯(劉友全等,2000)。例如尾巨桉和鄧恩桉對低溫具有不同的響應特性,低溫脅迫下SOD的活性均有升高,且耐寒的鄧恩桉的SOD活性高于尾巨桉(劉奕清,2015)。綜上所述,研究表明SOD的活性與桉樹對低溫脅迫的抵抗能力成正相關。
本實驗室成功克隆了巨尾桉的EuCuZnSOD基因(GenBank Accession Number: JX138573),通過原核表達,證明該基因具有生物學功能(趙艷玲和周利建,2015)。本研究通過基因工程手段獲得過量表達EuCuZnSOD的巨尾桉轉基因植株,研究超表達EuCuZnSOD與桉樹抗寒能力的關系,為定向培育耐低溫桉樹新品種提供直接的實驗基礎。
1 材料與方法
1.1 材料
EuCuZnSOD 為本研究實驗室克隆并保存(GenBank Accession Number: JX138573)。巨尾桉無菌苗、pBI121載體、JM109菌株、農桿菌LBA4404為本實驗室保存。PCR體系、內切酶等均購自Takara公司,其他試劑購自上海生工。引物由廈門精聚公司合成。
1.2 方法
1.2.1構建植物二元表達載體與農桿菌介導的葉盤法轉基因桉樹 pBI121和pMD-CSD質粒BamH I/Sac I雙酶切,雙酶切產物由TaKaRa MiniBEST Agarose Gel DNA Extraction Kit回收,T4 DNA ligase連接片段,雙酶切驗證和測序檢測陽性克隆載體。將pBIEuCSD轉化根癌農桿菌LBA4404感受態細胞,參照(周利建和趙艷玲,2012)葉盤法轉化巨尾桉,50 mg·L-1 Cef,5 mg·L-1 Kan,初步篩選轉基因桉樹。
1.2.2 轉基因巨尾桉的PCR檢測 CTAB法提取轉基因桉樹基因組DNA,引物1:5′-ATGGTGAAGGCCGTTGCCGTCC-3′;引物2:5′-TTAGCCTTGCAGACCAATAATAC-3′ 進行EuCuZnSOD基因的PCR擴增。以CaMV35s啟動子部分序列設計的引物進行二次驗證,引物3:5′-CAGGTCCCCAGATTAG CCTT-3′,引物4:5′-CGTGTTCTCT-CCAAATGA-3′,PCR反應程序如下:預變性94 ℃,5 min;94 ℃變性30 s,60 ℃退火30 s,72 ℃延伸90 s,30個循環;72 ℃延伸10 min,瓊脂糖凝膠電泳檢測。
1.2.3 轉基因巨尾桉的抗寒能力實驗 將轉基因組培苗-13.1 ℃冰箱暗處理,逆境處理3.5 h觀察植株情況,直到5 h,取出拍照后置于組培室進行恢復培養, 3 d和100 d后觀察植株生長狀況。
1.2.4 NBT法檢測SOD酶活性 SOD酶活性測定方法參照趙艷玲和周利建(2015)。植物材料取盆栽轉基因巨尾桉植株葉片,測定的條件分別是25 ℃、4 ℃恒溫氣候箱中培養36 h和48 h。
1.2.5 實時定量PCR技術分析轉基因植株EuCuZnSOD1表達量 巨尾桉葉片總RNA用Takara公司的RNA提取試劑盒,利用Takara SYBR ExScript 試劑和羅氏LightCycler 480儀器,以桉樹4個樣本的cDNA為模板,內參基因為巨尾桉18S rRNA(171 bp),引物設計分別為EuCSDF:5′-GGAAATGTCACTGTTGGTGC-3′;EuCSDR:5′-TCATCCGGATCAGCATGGAC-3′;Eu18SF:5′-CGCGCTACACTGATGTATTC-3′;Eu18SR:5′-GTACAAAGGGCAGGG ACGTA-3′,反應程序為預變性95 ℃,30 s;變性95 ℃,5 s;退火60 ℃,20 s,40個循環,最后延伸65 ℃,15 s,每個反應包括3個重復。
1.2.6 硫酸木質素含量測定 取巨尾桉轉基因組培苗全株做硫酸木質素含量測定,參照趙艷玲等(2012)的方法。
2 結果與分析
2.1 轉pBIEuCSD巨尾桉植株的獲得
pBIEuCSD植物的二元表達載體質粒通過BamHⅠ和SacⅠ雙酶切驗證連接成功,農桿菌介導的葉盤法轉基因巨尾桉,圖1所示為整個轉化過程的部分圖片,芽分化在培養基中培養60 d后,不定芽從愈傷組織中分化(圖1:A),芽長至2 cm移至生根培養基中繼續抗性篩選,抗Kan的轉基因巨尾桉正常生長(圖1:B)并生根(圖1:C),經初步篩選共得到46株抗性植株。
2.2 PCR檢測pBIEuCSD轉基因巨尾桉
對初步篩選所得到的46株轉基因植株進行PCR檢測。EuCuZnSOD1克隆自巨尾桉,以該基因作為陽性指標檢測到31株陽性植株,存在假陽性問題。以CaMV35S啟動子上的部分序列為模版設計引物,對得到的31株檢測陽性植株進行二次PCR驗證,若為陽性植株應有約900 bp的目的片段。從圖2可以看出,有10株轉基因植株在900 bp左右有條帶,通過雙PCR驗證,最終得到pBIEuCSD轉基因巨尾桉陽性植株10棵。
2.3 pBIEuCSD轉基因巨尾桉抗寒性分析
通過不同的低溫設置研究組培苗的最低耐受溫度, 經預實驗得到-13.1 ℃暗處理3.5 h后普通組培苗萎蔫現象嚴重甚至死亡。本實驗將生長狀況相同的轉基因陽性和陰性巨尾桉放置在同一組培瓶中培養至生根,于-13.1 ℃放置3.5 h,觀察發現36和47號轉基因植株有明顯凍傷,停止低溫處理,其余的轉基因陽性植株無萎蔫現象,重新放回-13.1 ℃放置5 h,結果發現49、52、53、55、44號轉基因陽性植株在凍害后表現為莖尖萎蔫,組培室培養72 h后植株莖上部1/3處萎蔫。40、41和42號巨尾桉凍害后植株無明顯凍傷,組培室培養 72 h后40號植株無萎蔫現象,41和42號植株莖尖有所萎蔫,不換瓶培養100 d后,40和41號巨尾桉全株成活,而42號植株底部葉片白化,重新發芽,如圖3所示。
表1統計了凍害后轉基因植株的恢復程度,經-13.1 ℃凍害實驗的植株光照培養100 d后,轉基因陽性植株只有47號全株白化死亡,對照植株有4株完全死亡;底部葉片白化但重新發芽的轉基因植株有4株,陰性對照有7株;有5株轉基因植株完全沒有白化現象,生長良好,但是完全存活的陰性植株為0株,說明EuCuZnSOD1基因轉入巨尾桉后,確能增強植株的抗寒能力且低溫處理后的復壯能力強。
2.4 pBIEuSOD陽性轉基因植株SOD酶活檢測
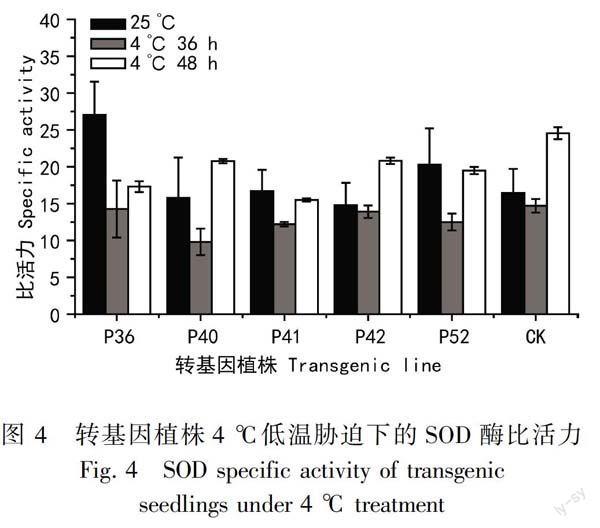
參考劉奕清(2015)的方法,本研究在4 ℃氣候箱中對盆栽轉基因巨尾桉植株低溫脅迫處理,測定SOD酶活性。從圖4結果可以看出,通過組培獲得的盆栽苗的預試驗發現在48 h的冷脅迫過程中,前36 h低溫脅迫下SOD酶的比活力呈現出下降趨勢,36 h左右SOD酶比活力達到最低,之后開始升高。
常溫環境下,轉基因植株的SOD比活力最高的是36號植株,但是36號植株的抗凍能力較差。抗凍能力最強的40和41號植株常溫下的SOD比活力和對照差異較小。4 ℃低溫脅迫下,巨尾桉抗凍能力較強的轉基因植株40對低溫脅迫較敏感,36 h脅迫后SOD比活力相比室溫下的比活力降低了38%,48 h比活力比36 h的上升了2.1倍,對照植株比活力分別是降低了11%和上升了1.6倍。從酶比活力的分析發現巨尾桉轉基因植株的SOD酶活性與抗凍能力的關聯性不強。
2.5 qPCR分析轉基因植株的EuCuZnSOD表達量
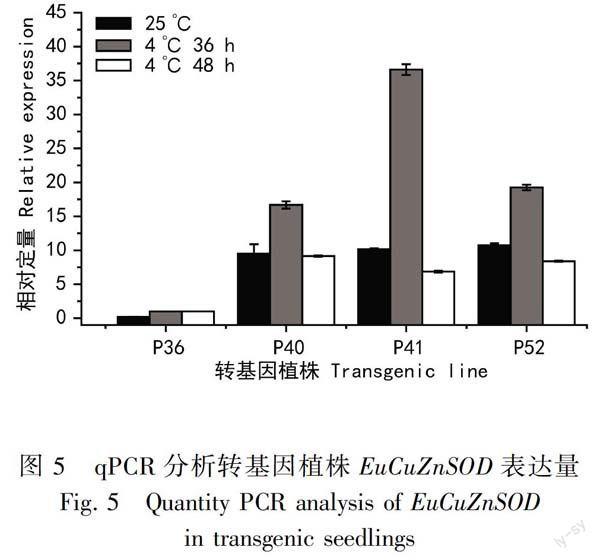
綜合抗寒性實驗和酶活性測定的實驗結果,選取4株轉基因巨尾桉進行實時定量PCR分析,包括酶活性最高但不耐凍的36號植株,最耐凍且酶活性對溫度敏感的40和41號植株,酶活性較高且較耐凍的52號植株,以盆栽苗的葉片為研究對象,選擇18S RNA作為內參基因對所有樣品進行歸一化處理,對不同樣品之間的EuCuZnSOD基因表達量進行比較如圖5。常溫下耐寒能力強的40、41和52號轉基因巨尾桉的EuCuZnSOD表達量是對照植株的9~10倍,在4 ℃低溫脅迫下EuCuZnSOD表達量具有先升高后降低的特點,在4 ℃低溫脅迫36 h時抗凍能力較強的40和41號轉基因植株EuCuZnSOD的表達量分別升高到對照的16倍和36倍,較高的EuCuZnSOD的表達有利于植株抵御凍害。
2.6 轉基因植株的硫酸木質素含量
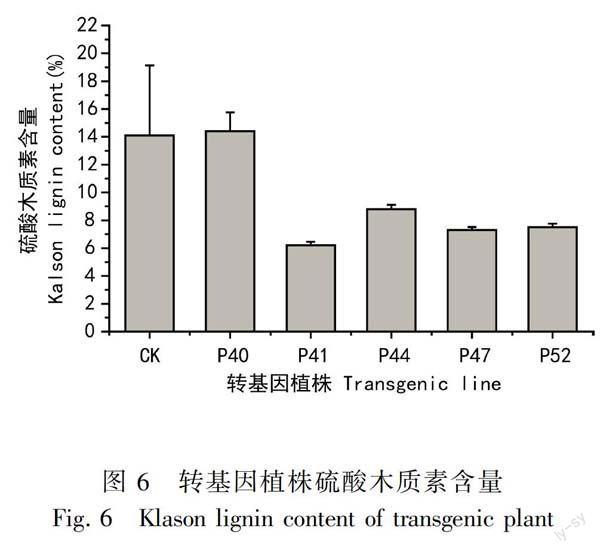
將轉基因植株組培苗的全株,包括根,莖,葉一起磨粉,硫酸木質素的含量測定結果見圖6,轉基因桉樹的硫酸木質素含量除了40號和對照相比變化較小外,其它植株的木質素含量都降低了,其中41號轉基因植株木質素含量比對照植株降低了56%。
3 討論與結論
桉樹是重要的紙漿林木,在林業生產中有非常明顯的區域性。目前,低溫、鹽堿、重金屬等環境因素嚴重限制了桉樹人工林的推廣及發展。CuZnSOD與植物的抗逆性有著密切關系。對CuZnSOD的轉基因研究主要集中在耐鹽、耐旱、耐儲存、耐凍等能力的研究。研究發現克隆多抗花生的AhCuZnSOD基因,獲得的轉基因煙草的抗鹽和抗旱能力明顯提高(Negi et al,2015)。小麥CuZnSOD基因TaSOD1.1和TaSOD1.2轉化煙草,轉基因植株抵御鹽脅迫能力增強(張海娜等,2008)。橡膠樹中過表達細胞質CuZnSOD基因,獲得耐旱的轉基因橡膠樹(Leclercq et al,2012)。李子的胞質CuZnSOD和APX轉入李子組培苗后,轉基因李子的快繁能力增強 (Faiza et al,2013)。
CuZnSOD基因和CAT1在木薯中過表達,木薯的采后生理惡化現象最少延遲10 d(Xu et al,2013)。實驗證明,將葉綠體CuZnSOD和APX基因轉入的甘薯具有較高的耐旱性且旱后恢復速度較快(Lu et al,2010),在鹽脅迫下CuZnSOD相比對照植株表達量增加了13.3倍,可耐受100 mmol·L-1 NaCl(Yan et al,2016),耐寒能力和耐受SO2的能力也有所增強(Kim et al,2015)。西伯利亞蓼銅伴侶蛋白基因(PsATX)和CuZnSOD共轉化的煙草耐鹽,且比轉單價基因植株具有更強的耐NaCl能力(馬靜等,2015)。本文的研究目標是通過CuZnSOD的過表達提高巨尾桉的抗寒能力,同時CuZnSOD的過表達可能也會增強巨尾桉對其它逆境環境的耐受能力,這方面的工作需要進一步的實驗確定。
通過實時定量PCR技術,可以研究CuZnSOD基因的表達量與低溫生理響應的關系。qPCR結果表明,雖然36號植株的SOD酶活性很高,但是CuZnSOD的表達量很低。抗寒能力較強的40、41和52號植株的CuZnSOD表達量較大,意味著CuZnSOD的過表達與轉基因巨尾桉的抗寒能力正相關。轉基因40和41號凍害后的恢復能力較強,而36和52號植株出現了寒害后白化并重生的現象,從48 h內的qPCR數據不能得出CuZnSOD表達量大與恢復能力有關,在組培苗-13.1 ℃凍害實驗后的觀察數據中,72 h的觀察數據與最終的100 d的成活程度也是不同的,說明CuZnSOD的表達量與低溫脅迫后的恢復能力可能需要更長時間的跟蹤監測數據。
通過硫酸木質素含量的分析,發現過表達細胞質EuCuZnSOD基因沒有導致木質素含量的升高,除了抗寒能力強的40號植株的木質素含量與對照差別較小外,其他轉基因植株的木質素含量都降低了,尤其是41號巨尾桉的組培苗整株的硫酸木質素含量只有6.2%,這與擬南芥(Gill et al, 2010)中大量表達CuZnSOD增加木質化的研究結果差異較大,具體原因還有待進一步的研究確定。通過對巨尾桉CuZnSOD功能的研究,篩選得到了抗寒且低木質素的轉基因巨尾桉,獲得既速生優質又多抗的桉樹新品系,使桉樹在冷涼的溫帶地區以及多種復雜逆境環境中的推廣種植成為可能。
參考文獻:
CAO PB, AZAR S, SAN CH, et al, 2015. Genome-wide analysis of the AP2/ERF family in Eucalyptus grandis: an intriguing over-representation of stress-responsive DREB1/CBF genes [J]. PLoS ONE, 10(4):e0121041.
FAIZE M, FAIZE L, PETRI C, et al, 2013. Cu/Zn superoxide dismutase and ascorbate peroxidase enhance in vitro shoot multiplication in transgenic plum [J]. J Plant Physiol, 170(7): 625-632.
GILL T, SREENIVASULU Y, KUMAR S, et al, 2010. Over-expression of superoxide dismutase exhibits lignification of vascular structures in Arabidopsis thaliana [J]. J Plant Phy-siol, 167(9) : 757-760.
KIM YH, LIM S, HAN SH, et al, 2015. Expression of both CuZnSOD and APX in chloroplasts enhances tolerance to sulfur dioxide in transgenic sweet potato plants [J]. C R Biol, 338(5):307-313.
LECLERCQ J, MARTIN F, SANIER C, et al, 2012. Over-expression of a cytosolic isoform of the HbCuZnSOD gene in Hevea brasiliensis changes its response to a water deficit [J]. Plant Mol Biol, 80(3): 255-272.
LEONARDI GA, CARLOS NA, MAZZAFERA P, et al, 2015. Eucalyptus urograndis stem proteome is responsive to short-term cold stress [J]. Genet Mol Biol, 38(2):191-198.
LIU YQ, LIU JL, PAN TL, 2000. Cold resistant of Eucalyptus camaldulensis with reference to growth study in Hunan [J]. J Centr S For Univ, 20(3): 86-89. [劉友全,劉加林,潘天玲, 2000. 赤桉在湖南的抗寒與生長適應性 [J]. 中南林學院學報,20(3): 86-89.]
LIU YQ, JIANG YS, LAN JB, et al,2014. Comparative transcriptomic analysis of the response to cold acclimation in Eucalyptus dunnii [J]. PLoS ONE, 9(ll):ell3091.
LIU RQ,2015. Physiological response characteristics and transcriptomic comparsion of two Eucalyptus trees under low temperature stress [M]. Beijing: China Agricultural University:15-18? [劉奕清,2015. 低溫脅迫下兩種桉樹的生理響應特征及轉錄表達差異研究 [M]. 北京: 中國農業大學:15-18.]
LU YY, DENG XP, SANGSOO K, 2010. Over expression of Cu, Zn superoxide dimutase (Cu, Zn-SOD) and ascorbate peroxidase (APX) in transgenic sweet potato enhances tolerance and recovery from drought stress [J]. Afr J Biotechnol, 9(49): 8378-8391.
KAYAL WE, NAVARRO M, MARQUE G, et al,2006. Expression profile of CBF-like transcriptional factor genes from Eucalyptus in response to cold [J]. J Exp Bot, 57(10): 2455-2469.
MA J, QU CP, XU ZR, et al, 2015. Salt tolerance of transgenic tobacco containing PsATX and PsSOD genes of polygonum sibiricum Laxm [J]. Bull Bot Res, 35(2):208-217.? [馬靜,曲春浦,許志茹,等, 2015. 西伯利亞蓼銅伴侶蛋白與銅鋅超氧化物歧化酶基因共轉化煙草的耐鹽性分析 [J]. 植物研究, 35( 2):208-217.]
NAVARRO M, AYAX C, MARTINEZ Y, et al, 2011. Two EguCBF1 genes overexpressed in Eucalyptus display a different impact on stress tolerance and plant development [J]. Plant Biotechnol J, 9(1):50-63.
NAVARRO M, MARQUE G, AYAX C, et al,2009. Complementary regulation of four Eucalyptus CBF genes under various cold conditions [J]. J Exp Bot, 60(9):2713-24.
NEGI NP, SHRIVASTAVA DC, SHARMA V, et al, 2015. Overexpression of CuZnSOD from Arachis hypogaea alleviates salinity and drought stress in tobacco [J]. Plant Cell Rep, 34(7): 1109-1126.
NGUYEN HC, CAO PB, SAN CH, et al, 2017. Special trends in CBF and DREB2 groups in Eucalyptus gunnii vs Eucalyptus grandis suggest that CBF are master players in the trade-off between growth and stress resistance [J]. Physiol Plant, doi: 10.1111/ppl.12529.
XU J, DUAN X, YANG J, et al, 2013. Enhanced reactive oxygen species scavenging by overproduction of superoxide dismutase and catalase delays postharvest physiological deterioration of cassava storage roots [J]. Plant Physiol, 161(3): 1517-1528.
XU FH, CHENG LJ, WEI XL, et al, 2016. Expression and function of EgrCR gene responding to cold stress in Eucalyptus grandis [J]. Sci Silv Sin, 52(3):59-67. [徐鳳華,程龍軍,魏曉玲等,2016. 巨桉低溫脅迫響應基因EgrCR的表達與功能 [J]. 林業科學,52(3):59-67.]
YAN H, LI Q, PARK SC, et al, 2016. Overexpression of CuZnSOD and APX enhance salt stress tolerance in sweet potato [J]. Plant Physiol Biochem, 109(2016):20-27.
ZHANG HN, LI XJ, LI CD, et al, 2008. Effects of overexpression of wheat superoxide dismutase (SOD) genes on salt tolerant capability in tobacco? [J]. Acta Agron Sin, 34(8): 1403-1408. [張海娜,李小娟,李存東,等,2008. 過量表達小麥超氧化物歧化酶(SOD)基因對煙草耐鹽能力的影響 [J].作物學報,34(8): 1403-1408.]
ZHAO YL, LU H, JIANG XN, 2012. Regulation of lignin biosynthesis in Populus tomentosa with GRP1.8 promoter and 4CL1 gene constructs [J]. J Chengdu Univ, 31(2):99-102.? [趙艷玲,陸海,蔣湘寧, 2012. GRP1.8融合4CL1基因調控楊樹木質素生物合成 [J]. 成都大學學報,31(2):99-102.]
ZHAO YL, ZHOU LJ, 2015. Cloning and expression of the cytosolic Copper/Zinc superoxide dismutase gene in Eucalyptus grandis × E. ophylla [J]. J Huaqiao Univ (Nat Sci Ed), 36(6):693-697. [趙艷玲,周利建, 2015. 巨尾桉胞質EuCuZnSOD基因的克隆與原核表達 [J]. 華僑大學學報(自然科學版),36(6):693-697.]
ZHOU LJ, ZHAO YL, 2012. Optimization of Eucalyptus grandis × E. ophylla tissue culture and regeneration system on genetic transformation [J]. Guangdong Agric Sci, 39(22):51-54.? [周利建, 趙艷玲, 2012. 巨尾桉組培快繁與轉基因再生體系的優化 [J]. 廣東農業科學, 39(22):51-54.]
