SIV感染猴外周血CD14+單核細(xì)胞CD169分子表達(dá)的變化
2018-05-31 08:45:52中國醫(yī)學(xué)科學(xué)院醫(yī)學(xué)實(shí)驗(yàn)動(dòng)物研究所北京協(xié)和醫(yī)學(xué)院比較醫(yī)學(xué)中心衛(wèi)計(jì)委人類疾病比較醫(yī)學(xué)重點(diǎn)實(shí)驗(yàn)室國家中醫(yī)藥管理局人類疾病動(dòng)物模型三級實(shí)驗(yàn)室新發(fā)再發(fā)傳染病動(dòng)物模型研究北京市重點(diǎn)實(shí)驗(yàn)室北京100021
中國比較醫(yī)學(xué)雜志 2018年5期
李 想,薛 婧,陳 霆,叢 喆,魏 強(qiáng)(中國醫(yī)學(xué)科學(xué)院醫(yī)學(xué)實(shí)驗(yàn)動(dòng)物研究所,北京協(xié)和醫(yī)學(xué)院比較醫(yī)學(xué)中心,衛(wèi)計(jì)委人類疾病比較醫(yī)學(xué)重點(diǎn)實(shí)驗(yàn)室,國家中醫(yī)藥管理局人類疾病動(dòng)物模型三級實(shí)驗(yàn)室,新發(fā)再發(fā)傳染病動(dòng)物模型研究北京市重點(diǎn)實(shí)驗(yàn)室,北京 100021)
唾液酸黏附素(sialic acid-binding immunoglobulin-type lectins, Siglecs)是宿主最大的唾液酸結(jié)合蛋白。Siglec-1,也被稱作CD169,是第一個(gè)被發(fā)現(xiàn)的Siglecs家族的成員,主要表達(dá)于單核巨噬細(xì)胞和樹突狀細(xì)胞[1 - 2]。CD169可與病毒表面的某些糖蛋白結(jié)合,增強(qiáng)病毒與細(xì)胞的結(jié)合,從而促進(jìn)病毒感染[3]。艾滋病又稱獲得性免疫缺陷綜合征(acquired immunodeficiency syndrome, AIDS),是由人類免疫缺陷病毒(human immunodeficiency virus,HIV)引起的,HIV主要分為兩種類型,即HIV-1和HIV-2,HIV-1是目前全球流行的主要病毒株。HIV-1研究中發(fā)現(xiàn),機(jī)體感染HIV-1后,組織巨噬細(xì)胞和外周血單核細(xì)胞表面高表達(dá)CD169[4]。猴免疫缺陷病毒(simian immunodeficiency virus,SIV)是從靈長類動(dòng)物身上分離得到的類似于HIV的病毒,SIV恒河猴模型被認(rèn)為是研究艾滋病最有效的模型,而SIVmac239是常用的SIV病毒。因此,本研究觀察了SIVmac239感染前后,恒河猴外周血單核細(xì)胞的比例及其表面CD169表達(dá)量的變化,分析了引起單核細(xì)胞CD169表達(dá)量變化的主要原因,以期為HIV-1/SIV感染單核巨噬細(xì)胞的研究提供新的思路。
1 材料和方法
1.1 實(shí)驗(yàn)材料
實(shí)驗(yàn)動(dòng)物:SPF級中國恒河猴40只,3~4歲,3~5 kg,雌雄各半,購自北京協(xié)爾鑫生物資源研究所[SCXK (京) 2015-0011]。動(dòng)物飼養(yǎng)及相關(guān)的實(shí)驗(yàn)在生物安全三級實(shí)驗(yàn)室中進(jìn)行[SYXK (京) 2015-0036]。實(shí)驗(yàn)前,經(jīng)血清學(xué)間接免疫熒光抗體檢查法(IFA)檢查排除猴B病毒(BV)、猴免疫缺陷病毒(SIV)、猴逆轉(zhuǎn)錄D型病毒(SRV-1)和猴T淋巴細(xì)胞性I型病毒(STLV-1)等相關(guān)病原體的感染。本實(shí)驗(yàn)通過本單位實(shí)驗(yàn)動(dòng)物管理和使用委員會的動(dòng)物倫理審查,批準(zhǔn)號為XJ16007。
病毒及動(dòng)物接種:SIVmac239,由美國Aaron Diamond艾滋病研究中心Preston Marx博士惠贈(zèng)。中國恒河猴PBMC擴(kuò)增制備,CEMx174細(xì)胞滴定TCID50為每毫升3 × 105。病毒使用劑量為500 TCID50,靜脈接種病毒。
樣本的收集:攻毒前和靜脈攻毒后第49天,采集恒河猴外周血2 mL,進(jìn)行全血流式;采集正常恒河猴外周血,分離PBMC,流式分選CD14+單核細(xì)胞,用含10%胎牛血清的RPMI-1640完全培養(yǎng)液,37℃、5% CO2孵箱中培養(yǎng)。
1.2 主要試劑與儀器
Recombinant Human IFN-alpha A protein(貨號:11100-1)購自R&D公司;流式抗體PE Mouse Anti-Human CD14(貨號:557154),F(xiàn)ITC Mouse Anti-Human CD14(貨號:555397),PE Mouse Anti-Human IFN-α(貨號:560097)和PerCP-CyTM5.5 Mouse Anti-Human CD16(貨號:560717)購自BD公司;流式抗體APC Anti-Human CD169(貨號:346008)購自BioLegend公司;Human M-CSF(貨號:300-25-10UG)和Human IL-13(貨號:200-13-10UG)購自Peprotech公司;Recombinant Rhesus Macaque IL-4 Protein(貨號:1577-IL-010/CF)購自R&D Systems公司;流式細(xì)胞洗液(PBS Ⅱ)為含2% FBS和2 mmol/L EDTA的PBS溶液。BD AccuriTMC6流式細(xì)胞儀,BD FACSAria Ⅱ流式細(xì)胞儀,日本Hitachi CF16RXII高速離心機(jī),Dynamica臺式離心機(jī),ESCO生物安全柜,Thermo Forma二氧化碳恒溫培養(yǎng)箱。
1.3 實(shí)驗(yàn)方法
1.3.1 流式細(xì)胞術(shù)分選恒河猴外周血CD14+單核細(xì)胞[5]
(1)恒河猴外周血PBMC分離:取40 mL正常恒河猴的外周血,2500 r/min離心10 min,棄去血漿,用RPMI-1640培養(yǎng)液等比稀釋血細(xì)胞,混勻后,每10 mL血細(xì)胞稀釋液緩慢加至5 mL Ficoll液面的上方(Ficoll與稀釋前血液的體積比為1∶1),2800 r/min離心30 min,用吸管將中間單個(gè)核細(xì)胞層吸出,轉(zhuǎn)移至另一新的離心管中,用不少于PBMC體積三倍的RPMI-1640培養(yǎng)液洗滌細(xì)胞兩次,2000 r/min離心10 min,適量的RPMI-1640培養(yǎng)液重懸后計(jì)數(shù)。
(2)流式分選單核細(xì)胞:按20 μL抗CD14抗體/1 × 106個(gè)細(xì)胞的濃度加入抗體,混勻,4℃孵育30 min后,用PBS Ⅱ洗2次,4℃、2000 r/min離心3 min,適量PBS Ⅱ重懸后,使用BD FACSAria Ⅱ流式儀進(jìn)行分選,得到CD14+單核細(xì)胞。
1.3.2 SIVmac239體外感染恒河猴外周血CD14+單核細(xì)胞[6]
取1 × 106個(gè)CD14+單核細(xì)胞置于24孔板中,1 mL完全培養(yǎng)液培養(yǎng)細(xì)胞,以100 μL的病毒用量感染細(xì)胞,混勻后置于37℃、5% CO2孵箱,培養(yǎng)48 h后,檢測細(xì)胞表面CD169的表達(dá)量。
1.3.3 細(xì)胞因子刺激恒河猴外周血CD14+單核細(xì)胞[3]
(1)細(xì)胞因子IFN-α刺激單核細(xì)胞:取1 × 106個(gè)CD14+單核細(xì)胞置于24孔板中,1 mL完全培養(yǎng)液培養(yǎng)細(xì)胞,加入細(xì)胞因子IFN-α,使得IFN-α的終濃度為500 U/mL,48 h后檢測細(xì)胞表面CD169的表達(dá)量。
(2)細(xì)胞因子M-CSF、IL-4、IL-13刺激單核細(xì)胞:取1 × 106個(gè)CD14+單核細(xì)胞置于24孔板中,1 mL完全培養(yǎng)液培養(yǎng)細(xì)胞,加入細(xì)胞因子M-CSF、IL-4和IL-13,使得M-CSF、IL-4和IL-13的終濃度均為20 ng/mL,48 h后檢測細(xì)胞表面CD169的表達(dá)量。
1.3.4 流式細(xì)胞術(shù)檢測細(xì)胞表面CD169分子表達(dá)量[7 - 8]
(1)全血流式:取100 μL全血,依次加入抗體PE Mouse Anti-Human CD14,PerCP-CyTM5.5 Mouse Anti-Human CD16,APC Anti-Human CD169,室溫下避光孵育30 min,加入1 mL終濃度為10%的紅細(xì)胞裂解液,室溫作用8 min,加入1 mL PBS,2500 r/min離心5 min,棄去上清,加入2 mL PBS,2000 r/min離心10 min,適量PBS重懸過濾后BD AccuriTMC6流式細(xì)胞儀進(jìn)行樣本檢測。
(2)胞外染色:將CD14+單核細(xì)胞從培養(yǎng)板中取出后,用PBS Ⅱ洗2次,4℃、2000 r/min離心3 min,調(diào)整細(xì)胞濃度為每毫升1 × 107個(gè)細(xì)胞,輕輕混勻后加入流式管內(nèi),每管100 μL。各流式管中加入相應(yīng)抗體,4℃避光孵育30 min,PBS Ⅱ洗1次,PBS洗1次,PBS重懸后細(xì)胞篩過濾上機(jī)檢測。
(3)胞內(nèi)染色:每個(gè)流式管內(nèi)加入100 μL IC Fixation Buffer,4℃孵育20 min,加入2 mL 1 × Permeabilization Buffer洗2次,4℃、2000 r/min離心3 min。100 μL 1 × Permeabilization Buffer重懸細(xì)胞后加入相應(yīng)抗體,混勻后4℃避光孵育30 min,1 × Permeabilization Buffer洗2次,PBS洗1次,PBS重懸后細(xì)胞篩過濾上機(jī)檢測。
1.4 統(tǒng)計(jì)學(xué)方法
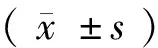
2 結(jié)果
2.1 SIVmac239感染猴CD14+單核細(xì)胞表面分子CD169表達(dá)量分析
正常恒河猴經(jīng)SIVmac239感染后,采集外周血,流式細(xì)胞術(shù)檢測CD14+單核細(xì)胞的比例及其表面CD169分子表達(dá)量。結(jié)果顯示,正常恒河猴外周血單核細(xì)胞低表達(dá)CD169,感染后,CD14+單核細(xì)胞的比例雖出現(xiàn)下降(P< 0.01)(圖1A),但CD14+單核細(xì)胞表面CD169的表達(dá)量卻顯著增加(P< 0.01)(圖1B)。
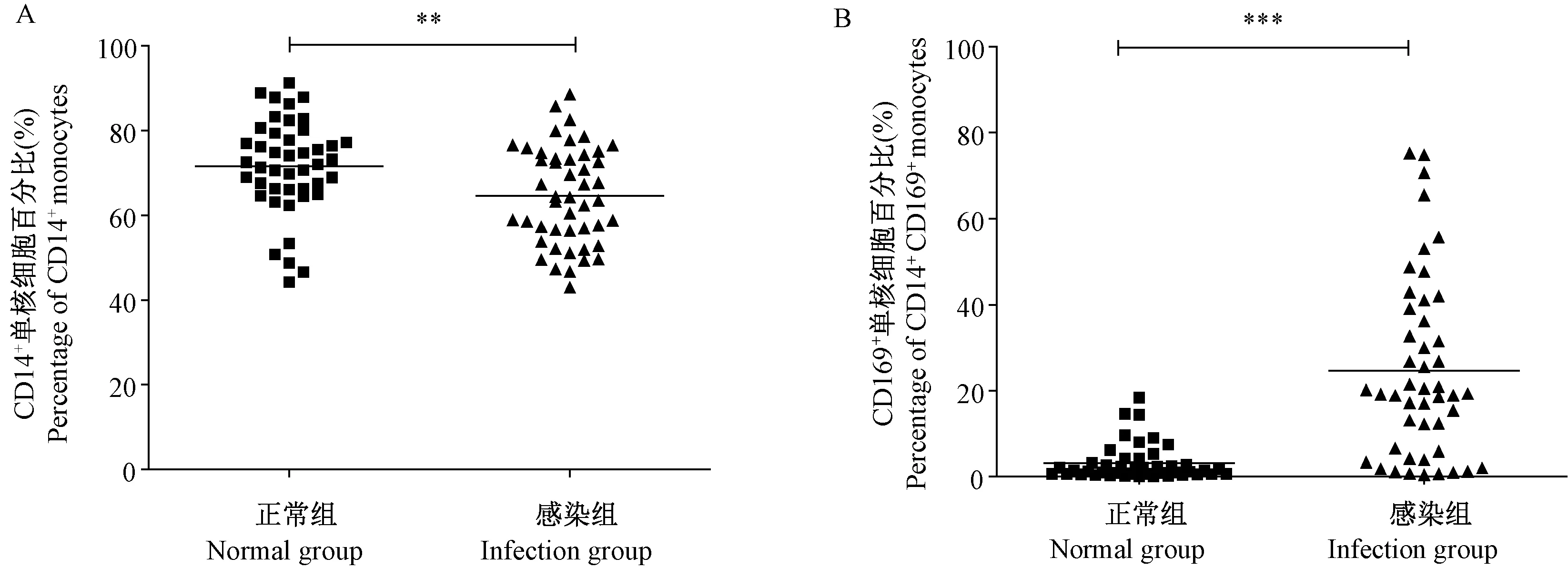
注:A:SIVmac239感染前后外周血單核細(xì)胞比例的變化;B:SIVmac239感染前后外周血單核細(xì)胞表面分子CD169表達(dá)量的變化。與正常組相比,** P< 0.01,*** P< 0.001;n=40。圖1 CD14+單核細(xì)胞及其表面分子CD169在SIVmac239感染后表達(dá)量的變化Note.A: Changes in the percentage of peripheral blood CD14+ monocytes before and after SIVmac239 infection. B: Changes in the expression of CD169 on the surface of peripheral blood CD14+ monocytes before and after SIVmac239 infection. Compared with the normal group,**P< 0.01,***P< 0.001. n =40.Fig.1 Changes in the percentage of CD14+ monocytes in the peripheral blood and the expression of CD169 on their surface before and after SIVmac239 infection
2.2 SIVmac239感染前后恒河猴外周血不同單核細(xì)胞亞群CD169分子表達(dá)量分析
根據(jù)外周血單核細(xì)胞CD14和CD16表達(dá)量的差異,可將單核細(xì)胞分為三種亞型,經(jīng)典型CD14++CD16-、中間過渡型CD14++CD16+和非經(jīng)典型CD14+CD16++單核細(xì)胞。SIVmac239感染恒河猴前后,流式檢測不同亞群單核細(xì)胞比例及其表面分子CD169表達(dá)量的變化。結(jié)果顯示,感染后經(jīng)典型CD14++CD16-和非經(jīng)典型CD14+CD16++單核細(xì)胞的比例未發(fā)生明顯的變化,中間過渡型CD14++CD16+單核細(xì)胞的比例出現(xiàn)下降(P< 0.01)(圖2 A)。三種亞型單核細(xì)胞表面CD169的表達(dá)量均出現(xiàn)不同程度的增高(P< 0.01)(圖2B)。其中,經(jīng)典型CD14++CD16-和非經(jīng)典型CD14+CD16++單核細(xì)胞表面CD169表達(dá)量升高較為明顯,分別為感染前的10倍和11倍(圖2C)。進(jìn)一步分析可知,SIVmac239感染后,非經(jīng)典型CD14+CD16++單核細(xì)胞表面分子CD169的表達(dá)量明顯高于經(jīng)典型CD14++CD16-單核細(xì)胞(P< 0.01)(圖2D)。

注:A:SIVmac239感染前后外周血不同亞群單核細(xì)胞比例的變化;B:SIVmac239感染前后外周血不同亞群單核細(xì)胞表面分子CD169表達(dá)量的變化;C:SIVmac239感染前后不同亞群單核細(xì)胞表面分子CD169表達(dá)量的比較;D:SIVmac239感染后經(jīng)典型和非經(jīng)典型單核細(xì)胞表面分子CD169表達(dá)量的比較。與正常組相比,*** P< 0.001,ns:差異無顯著性;與CD14++CD16-CD169+相比,△△P< 0.01;n=40。圖2 不同單核細(xì)胞亞群比例及其表面分子CD169在SIVmac239感染后表達(dá)量的變化Note.A: Changes in the percentage of different subsets of peripheral blood monocytes. B: Changes in the expression of CD169 on the surface of different subsets of peripheral blood monocytes. C: Comparison of the relative expression of CD169 on the surface of different subsets of monocytes. D: Comparison of the expression of CD169 on the surface of classical and non-classical monocytes after SIVmac239 infection. Compared with the normal group,***P< 0.001, ns: not significant. Compared with CD14++CD16-CD169+,△△P< 0.01. n=40.Fig.2 Changes in the percentage of different subsets of monocytes and the expression of CD169 on their surface after SIVmac239 infection
2.3 SIVmac239直接感染和不同細(xì)胞因子刺激CD14+單核細(xì)胞后CD169分子表達(dá)量的變化
正常恒河猴外周血流式分選CD14+單核細(xì)胞,細(xì)胞因子M-CSF、IL-4和IL-13等不能刺激CD14+單核細(xì)胞表面表達(dá)CD169(圖3A),但經(jīng)細(xì)胞因子IFN-α刺激48 h后,細(xì)胞表面高表達(dá)CD169,陽性率達(dá)到(98.9±0.89)%(圖3B)。SIVmac239體外直接感染CD14+單核細(xì)胞,未能檢測到CD169表達(dá)的變化(圖3C),同時(shí)細(xì)胞內(nèi)也未能檢測到細(xì)胞因子IFN-α的表達(dá)(圖3D)。
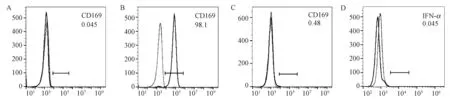
注:A:M-CSF、IL-4和IL-13誘導(dǎo)的CD14+單核細(xì)胞表面分子CD169表達(dá)量的變化;B:IFN-α誘導(dǎo)的CD14+單核細(xì)胞表面分子CD169表達(dá)量的變化;C:SIVmac239感染CD14+單核細(xì)胞后CD169表達(dá)量的變化;D:SIVmac239感染CD14+單核細(xì)胞后胞內(nèi)細(xì)胞因子IFN-α表達(dá)量的變化。“……”:同型對照;“——”:CD169/IFN-α。圖3 SIVmac239直接感染和不同細(xì)胞因子刺激的CD14+單核細(xì)胞表面分子CD169的變化Note.A: Changes in the expression of CD169 on the surface of CD14+ monocytes induced by M-CSF, IL-4 and IL-13. B: Changes in the expression of CD169 on the surface of CD14+ monocytes induced by IFN-α. C: Changes in the expression of CD169 on the surface of CD14+ monocytes after direct infection with SIVmac239. D: Changes in the expression of intracellular cytokine IFN-α of CD14+ monocytes after direct infection with SIVmac239. “……”: Isotype; “——”: CD169/IFN-α.Fig.3 Changes in the expression of CD169 on the surface of CD14+ monocytes after stimulated by different cytokines or directly infected with SIVmac239
3 討論
單核巨噬細(xì)胞可以被HIV-1感染,成為HIV-1的潛伏感染細(xì)胞,其感染機(jī)制目前尚不完全明確。表達(dá)于單核巨噬細(xì)胞表面的CD169由于可以與病毒表面某些糖蛋白結(jié)合而促進(jìn)病毒感染,從而成為HIV-1感染機(jī)制研究中的一個(gè)新的關(guān)注點(diǎn)。
本研究中發(fā)現(xiàn),恒河猴外周血CD14+單核細(xì)胞在SIVmac239感染后比例略降低,這可能與外周血單核細(xì)胞向不同的組織遷徙分化為巨噬細(xì)胞有關(guān)[9]。同時(shí),感染猴外周血CD14+單核細(xì)胞表面CD169的表達(dá)量顯著增加,這與HIV-1感染后,組織巨噬細(xì)胞和外周血單核細(xì)胞高表達(dá)CD169的結(jié)果一致[4]。由此可見,單核巨噬細(xì)胞上表達(dá)的CD169分子確實(shí)和病毒的感染有關(guān)。
Ziegler-Heitbrock[10]根據(jù)外周血單核細(xì)胞CD14和CD16表達(dá)量的差異,將單核細(xì)胞分為三種亞型,經(jīng)典型CD14++CD16-,中間過渡型CD14++CD16+和非經(jīng)典型CD14+CD16++單核細(xì)胞。外周血單核細(xì)胞來源于骨髓,剛進(jìn)入外周血時(shí)為經(jīng)典型CD14++CD16-單核細(xì)胞,其中的一小部分分化為中間過渡型CD14++CD16+單核細(xì)胞,這類細(xì)胞最終可能分化為非經(jīng)典型CD14+CD16++單核細(xì)胞,進(jìn)入不同的組織,產(chǎn)生不同類型的終末分化巨噬細(xì)胞[11]。研究發(fā)現(xiàn),非經(jīng)典型CD14+CD16++單核細(xì)胞相比經(jīng)典型CD14++CD16-單核細(xì)胞具有更強(qiáng)的吞噬能力和抗原提呈能力[9, 12],是發(fā)育更為成熟的細(xì)胞。恒河猴經(jīng)SIVmac239感染后,非經(jīng)典型CD14+CD16++單核細(xì)胞中CD169表達(dá)量的升高最為明顯,其表達(dá)量明顯高于經(jīng)典型CD14++CD16-單核細(xì)胞,推測CD14+CD16++單核細(xì)胞更強(qiáng)的吞噬能力和抗原提呈能力可能與CD169具有吞噬和介導(dǎo)抗原提呈的作用[13 - 14]有關(guān)。
本研究還發(fā)現(xiàn),SIVmac239體外直接感染正常恒河猴外周血CD14+單核細(xì)胞并不能引起CD169表達(dá)的增加,而使用IFN-α刺激可誘導(dǎo)單核細(xì)胞高表達(dá)CD169分子。但是,SIVmac239直接感染單核細(xì)胞并不能引起其本身分泌IFN-α,而使用其它細(xì)胞因子也不能引起單核細(xì)胞表達(dá)CD169。說明恒河猴外周血單核細(xì)胞CD169表達(dá)的增加不是病毒感染直接引起的,而是體內(nèi)其它免疫細(xì)胞分泌的細(xì)胞因子IFN-α引起的。
綜上所述,在HIV-1/SIV感染過程中,單核巨噬細(xì)胞表面CD169的表達(dá)量發(fā)生明顯變化,其表達(dá)與病毒感染機(jī)體后其它細(xì)胞釋放的細(xì)胞因子IFN-α相關(guān),因此,對CD169的進(jìn)一步研究可能為HIV-1感染機(jī)制的研究提供新的思路。
參考文獻(xiàn):
[1] Crocker PR, Paulson JC, Varki A. Siglecs and their roles in the immune system [J]. Nat Rev Immunol, 2007, 7(4): 255-266.
[2] Crocker PR, Mucklow S, Bouckson V, et al. Sialoadhesin, a macrophage sialic acid binding receptor for haemopoietic cells with 17 immunoglobulin-like domains [J]. EMBO J, 1994, 13(19): 4490-4503.
[3] Akiyama H, Ramirez NP, Gibson G, et al. Interferon-inducible CD169/Siglec1 attenuates anti-HIV-1 effects of IFN-α [J]. J Virol, 2017, 91(21): JVI.00972-17.
[4] van der Kuyl AC, van den Burg R, Zorgdrager F, et al. Sialoadhesin (CD169) expression in CD14+cells is upregulated early after HIV-1 infection and increases during disease progression [J]. PLoS One, 2007, 2(2): e257.
[5] 趙文秀, 張正奇, 許雅蘋, 等. 不同流式抗體分選小鼠原位肝癌模型中髓系來源抑制性細(xì)胞的比較 [J]. 中國實(shí)驗(yàn)動(dòng)物學(xué)報(bào), 2013, 21(4): 42-46.
[6] Zheng Q, Hou J, Zhou Y, et al. Siglec1 suppresses antiviral innate immune response by inducing TBK1 degradation via the ubiquitin ligase TRIM27 [J]. Cell Res, 2015, 25(10): 1121-1136.
[7] 趙長城, 高錫強(qiáng), 薛婧, 等. IL-21對恒河猴SHIV特異性CD8+T細(xì)胞毒性效應(yīng)的影響 [J]. 中國實(shí)驗(yàn)動(dòng)物學(xué)報(bào), 2012, 20(5): 29-34.
[8] 彭卓穎, 叢喆, 李想, 等. 不同方法誘導(dǎo)THP-1細(xì)胞分化效果比較 [J]. 中國比較醫(yī)學(xué)雜志, 2017, 27(9): 1-7.
[9] 王鑫, 康富標(biāo), 王桂平, 等. CD14+CD16+亞型單核細(xì)胞的研究進(jìn)展 [J]. 免疫學(xué)雜志, 2012, 28(10): 910-913.
[10] Ziegler-Heitbrock L. Monocyte subsets in man and other species [J]. Cell Immunol, 2014, 289(1-2): 135-139.
[11] Moniuszko M, Liyanage NP, Doster MN, et al. Glucocorticoid treatment at moderate doses of SIVmac251-infected rhesus macaques decreases the frequency of circulating CD14+CD16++monocytes but does not alter the tissue virus reservoir [J]. AIDS Res Hum Retroviruses, 2015, 31(1): 115-126.
[12] Boudjeltia KZ, Brohee D, Piro P, et al. Monocyte-platelet complexes on CD14/CD16 monocyte subsets: relationship with ApoA-I levels. A preliminary study [J]. Cardiovasc Pathol, 2008, 17(5): 285-288.
[13] Izquierdo-Useros N, Lorizate M, McLaren PJ, et al. HIV-1 capture and transmission by dendritic cells: the role of viral glycolipids and the cellular receptor Siglec-1 [J]. PLoS Pathog, 2014, 10(7): e1004146.
[14] De Schryver M, Leemans A, Pintelon I, et al. Comparative analysis of the internalization of the macrophage receptor sialoadhesin in human and mouse primary macrophages and cell lines [J]. Immunobiology, 2017, 222(6): 797-806.
