植物呼吸釋放CO2碳同位素變化研究進展
2018-06-07 03:07:46鐘尚志崔海瑩
生態學報 2018年8期
柴 華,鐘尚志,崔海瑩,李 杰,孫 偉
東北師范大學草地科學研究所植被生態科學教育部重點實驗室, 長春 130024
碳循環是最重要的生物地球化學循環之一[1- 2]。隨著CO2濃度升高引起的氣候變化,碳循環過程及其與氣候變化之間反饋調節機制成為生態學研究的一個熱點[3- 4]。穩定性碳同位素技術作為探究碳循環過程的重要手段,在不同時間和空間尺度生態與環境問題研究中得到廣泛應用[5- 7]。同時,探究碳循環不同過程穩定性碳同位素組成、變化趨勢及其控制機制是當前生態學研究的另一個熱點,有助于研究者更好地將碳同位素技術應用于生態系統碳循環過程研究[8]。
植物在生態系統與大氣CO2交換過程中起到非常重要的作用[9]。植物通過光合作用吸收大氣中的CO2,一部分光合產物儲存在生物體中以維持自身的生長、繁殖等生命活動,另一部分通過植物的呼吸作用返回到大氣中。在此過程中,植物通過光合作用和呼吸作用對近地面大氣中CO2的穩定性碳同位素組成(13CO2豐度)產生影響。其中,光合作用增加了近地面大氣中的13CO2豐度,而呼吸作用則趨于稀釋空氣中的13CO2豐度[10]。
植物呼吸釋放CO2的碳同位素組成(δ13CR)受環境變化導致的光合判別和光合產物同位素組成影響,與大氣中的13CO2存在顯著差異[11- 13]。光合產物后續代謝過程的同位素分餾(isotope fractionation)也能夠影響呼吸底物的碳同位素組成和植物不同器官呼吸釋放的CO2碳同位素組成,并通過改變碳同位素通量的時空變化影響生態系統呼吸CO2的碳同位素組成,最終影響基于穩定性同位素技術的碳通量估測[14-15]。因此,植物δ13CR的碳同位素信號不僅有助于探究生態系統與大氣的碳通量[16-17],區分生態系統呼吸[18-19],也能反映植物的生理過程[15]、碳分配方式[20]及植物應對環境變化的適應策略[21- 22]。
目前,國際上關于植物δ13CR的碳同位素組成、短期變化特征及其潛在生理機制已開展了一系列的研究工作[8,14,23],提出了底物同位素控制、呼吸中間產物分配、光照增強暗呼吸(LEDR, light-enhanced dark respiration)、呼吸過程同位素分餾變化等生理生態機制,但內在機制尚不清楚。本文通過概述國內外植物呼吸釋放13CO2短期動態變化的研究概況,分析影響植物δ13CR短期變化的潛在機制,旨在推動穩定性碳同位素技術在生態系統碳循環研究中的應用。
1 植物呼吸釋放13CO2短期動態變化研究概況
近年來,國內外學者針對植物δ13CR短期動態變化開展了一系列研究工作,發現植物δ13CR短期動態存在同種植物不同器官以及植物功能群差異。植物葉片暗呼吸釋放CO2碳同位素組成有較大的變異性,變化范圍為:(-31.9±0.3)‰—(-13.8±1.0)‰,晝夜變化幅度的最大值為11.5‰;樹干/莖暗呼吸釋放CO2碳同位素組成變化范圍為:(-32.1±0.8)‰—(-21.2±0.3)‰,晝夜變化幅度的最大值為4.0‰;植物根系暗呼吸釋放CO2碳同位素組成變化范圍為:(-33.3±0.5)‰—(-16.3±1.9)‰,晝夜變化幅度的最大值為5.4‰[23]。
不同功能群植物δ13CR晝夜變化幅度研究較少,且結果存在不一致性。Priault等[3]通過對16種植物葉片暗呼吸研究發現,不同功能群植物存在δ13CR晝夜變化差異:慢速生長和芳香類植物的δ13CR有晝夜變化,幅度范圍為1.4‰—7.9‰;草本、快速生長植物δ13CR則沒有顯著的晝夜變化。Cui等[24]通過對22種植物葉片暗呼吸研究發現,C3植物和C4植物δ13CR夜間變化幅度差異不顯著,并且δ13CR夜間變化幅度在木本植物、雜類草和禾草之間也沒有顯著的差異。植物δ13CR的功能群差異需要深入研究。
植物暗呼吸釋放CO2碳同位素組成與呼吸底物碳同位素組成的相關性研究結果并不一致:部分研究表明,植物δ13CR與呼吸底物碳同位素組成具有相關性,但植物δ13CR的變化幅度大于呼吸底物碳同位素組成的變化范圍[17, 25- 29];也有一些研究表明,植物呼吸底物碳同位素組成并沒有顯著的晝夜變化,但植物δ13CR卻有顯著的變化[28,30-33];此外,也有研究表明植物δ13CR的變化幅度與呼吸底物的碳同位素組成呈負相關關系[27,34]。植物自養呼吸底物十分復雜,可以分為快速周轉和慢速周轉碳水化合物兩類,其構成和穩定性碳同位素組成均有差異,很難在提取過程中將二者區分開,這也是導致植物δ13CR與底物碳同位素組成之間關系不確定的原因之一[35]。總體而言,目前對植物葉片δ13CR與呼吸底物碳同位素組成關系研究較多,但缺乏對植物其他特定組織的研究,如樹干、莖、根系呼吸釋放CO2碳同位素組成及其潛在生理機制的研究。
迄今為止,國內關于植物δ13CR短期動態變化的研究鮮有報道。近年來,植物δ13CR的動態變化及其調控機制是生態學的研究熱點之一,但國內關于此方面的研究較少。因此,亟需加強我國關于植物δ13CR碳同位素組成及其潛在呼吸代謝機制的研究。
2 植物δ13CR短期變化的驅動機制
碳從大氣經由植物葉片、莖、根系和土壤返回大氣過程中涉及眾多物理和生化過程,與之相關的同位素分餾和同位素判別(isotope discrimination)效應顯著改變13C豐度。
Farquhar等[36]對同位素分餾和同位素判別給出定義:“同位素分餾”是指某一反應過程中同位素以不同的比例分配到不同的物質中;而“同位素判別”是指某一反應中或某催化劑由于同位素在質量上的差異,使其對重同位素有識別和排斥的作用,致使產物的重同位素含量減少的現象。同位素分餾表示的是反應物同位素組成改變的效果,而同位素判別表示的是造成反應物同位素組成改變的一種過程或原因[37]。受生長環境、遺傳特性、功能群差異影響,植物光合、呼吸過程相關同位素效應存在變異,導致呼吸釋放CO2碳同位素組成和短期變化幅度存在差異。已有研究結果表明,這些因素對植物δ13CR的作用并不是單一進行的,而是共同影響并可能存在相互作用[3,23]。
2.1 光合作用及其產物的同位素效應
2.1.1 光合碳同位素效應
植物通過光合作用合成13C貧化的含碳化合物,這些光合產物是植物呼吸的主要底物。因此,光合碳固定過程中,碳同位素分餾能夠通過改變光合產物穩定性碳同位素組成,進而影響植物δ13CR及其變化幅度。植物葉片光合碳固定過程對于13C的同位素分餾主要包含兩個過程:(1)大氣中的CO2經由植物氣孔向葉片擴散時發生同位素分餾(4.4‰)。12C、13C在化學性質上沒有明顯的差別,但由于質量上的細微差異使得大氣中13CO2的擴散速率比12CO2慢,植物優先吸收12CO2而排斥13CO2。因此,進入葉肉細胞間隙的CO2其13C豐度低于大氣CO2;(2)CO2同化過程中,二磷酸核酮糖羧化酶(ribulose bisphosphate carboxylase, RuBPCase, 29‰)和磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPCase, 5.7‰)對穩定性碳同位素存在分餾作用,溶解在細胞質中的12CO2優先通過酶的作用結合為磷酸甘油酸,因此合成的光合產物13C貧化[10]。
光合碳同位素判別僅在白天發生,但其不僅影響白天δ13CR,也可能影響夜晚δ13CR。植物夜晚呼吸利用底物主要是日間光合所積累的淀粉,淀粉在合成和分解過程中存在一定的時間差異,清晨和上午合成的淀粉處于淀粉顆粒的核心,通常在凌晨時分被分解利用[32,38]。白天光照、溫度等環境條件變化導致的光合判別變化會通過影響淀粉顆粒不同部位穩定性碳同位素組成進而影響夜晚δ13CR。然而,δ13CR的晝夜變化并不能完全由光合碳同位素判別解釋[23]。光合產物后續代謝過程同樣存在潛在的同位素判別,例如,暗呼吸和光合產物由葉片向植物下游組織輸出過程亦伴隨同位素判別[39]。基于同位素模型預測研究發現,夜晚蓖麻(RicinuscommunisL.)的韌皮部和菜豆(PhaseolusvulgarisL.)、林煙草(NicotianasylvestrisSpeg.)、向日葵(HelianthusannuusL.)的葉片糖類δ13C值的變異度遠大于預計的僅受光合判別作用的值[29,40]。因此,在研究植物δ13CR的短期變化時,既要考慮光合產物形成過程的同位素判別,又不能忽視光合產物后續代謝過程同位素效應的作用。與其他器官相比,葉片暗呼吸釋放CO2呈13C富集態勢,而葉片的δ13C值卻低于植物其他器官,光合產物后續代謝過程同位素判別可以解釋這種普遍觀測到的植物器官間碳同位素差異現象[39]。
2.1.2 光合產物后續代謝過程同位素分餾
在卡爾文循環中,代謝分支點以及磷酸丙糖(triose phosphate, TP, 光合作用合成的最初糖類)向細胞溶質輸出或繼續在卡爾文循環中使用均存在碳同位素分餾[41]。在植物葉綠體中,卡爾文循環形成磷酸丙糖,經過各種酶的催化形成淀粉。在此過程中由于醛縮酶(aldolase)同位素效應的影響,使得淀粉中13C的豐度高于可溶性糖[42-43],而剩余的13C貧化的磷酸丙糖合成蔗糖從葉綠體中輸出,進而改變呼吸底物的13C組成。同時,白天光合作用積累的淀粉在夜晚分解用于植物呼吸代謝和向其他器官的光合產物輸出,造成葉片和植物韌皮部輸出糖類δ13C值的晝夜差異,白天蔗糖中13C貧化,夜晚蔗糖中13C富集[29,40]。植物韌皮部的蔗糖混合了具有不同代謝歷史和滯留時間的蔗糖分子[25],其δ13C值取決于植物最初積累淀粉的δ13C值,以及糖類在植物莖部由上至下的傳輸距離[23]。總體來講,糖類在植物由上往下的傳輸過程中,由于同位素分餾的作用,使得不同類型和不同部位的呼吸底物δ13C值及其日變化幅度存在差異,從而影響植物莖和根呼吸釋放CO2的δ13CR值及其日變化幅度。
2.2 呼吸碳同位素分餾
由于植物呼吸代謝過程中細胞、組織以及植物類型的不同,導致不同的代謝途徑呼吸釋放的CO2同位素組成存在差異。植物呼吸過程碳同位素分餾包括糖類分子結構上13C不均衡分布(non-statistical13C distribution)導致的裂解分餾、呼吸酶的同位素效應、次生代謝過程中的同位素分餾[10]。
2.2.1 裂解分餾
由于醛縮酶的同位素分餾效應,導致葡萄糖分子中13C的不均勻分布,與葡萄糖分子整體13C豐度相比,其中C- 3和C- 4富含13C,而其他位置的碳則呈13C貧化趨勢(表1)[43-45]。不同光合途徑植物在葡萄糖分子13C分布不均勻性上存在差異,主要是因為光呼吸導致二磷酸核酮糖(RuBP)分子內部結構存在差異,在光合作用下二磷酸核酮糖還原為磷酸丙糖,最終合成糖類物質[45]。

表1 葡萄糖分子內部13C的不均衡分布(不同位置C原子相對于葡萄糖分子δ13C平均值的差值)
C- 1:葡萄糖分子中第一位碳原子,C- 2:葡萄糖分子中第二位碳原子,后同,數據來源于Rossmann等[44]
在糖酵解過程中,葡萄糖首先活化為1,6-二磷酸果糖,然后裂解為3-磷酸甘油醛和磷酸二羥丙酮,接著3-磷酸甘油醛氧化釋放能量,并最終生成丙酮酸。在此過程中,從富含13C葡萄糖的C- 3和C- 4轉化為C- 1的丙酮酸,在丙酮酸脫氫酶(PDH)作用下釋放富含13C的CO2。含有相對13C貧化碳分子的乙酰輔酶A(Acetyl CoA)既可以進入檸檬酸循環,經過脫羧作用,分解產生ATP(adenosine triphosphate),又可以被用于合成次生代謝產物[3]。當所有乙酰輔酶A均用于呼吸作用時,所產生CO2的13C豐度與底物葡萄糖一致;當部分乙酰輔酶A用于合成次生代謝產物時,呼吸釋放CO2的13C豐度則高于底物葡萄糖[23](圖1)。很多研究發現植物暗呼吸釋放CO2的13C豐度高于其呼吸底物的13C豐度,并歸因于乙酰輔酶A未完全用于呼吸分解所導致的同位素分餾[46-50]。
Priault等[3]通過丙酮酸13C標記研究表明,在丙酮酸脫羧反應和檸檬酸循環中均存在碳同位素分餾。灌木植物歐洲半日花(HalimiumhalimifoliumL.)在丙酮酸脫氫酶的作用下,釋放富含13C的CO2,而剩余的丙酮酸進入次生代謝的碳通量超過進入檸檬酸循環的碳通量數倍,致使δ13CR的日變化幅度增加;而快速生長的草本植物三角紫葉酢漿草(OxalistriangularisA. St-Hil.)的次生代謝活動和檸檬酸循環都保持低的活性,因此δ13CR的日變化幅度不明顯(圖1)。
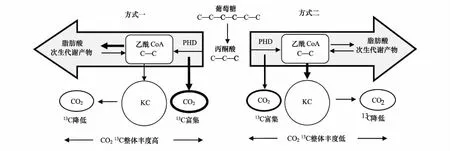
圖1 植物呼吸代謝中間產物利用對暗呼吸δ13CR的影響Fig.1 Major expected fluxes of respiratory substrates explaining δ13CR of dark-respired CO2 depending on the respiratory energy demand PDH: 丙酮酸脫氫酶 pyruvate dehydrogenase; KC: 檸檬酸循環 Krebs Cycle; 粗體C代表含有高豐度的13C,陰影部分表示主要的呼吸底物的通量,黑色箭頭代表碳的通量,改繪自Priault等[3]
2.2.2PEPCase對植物釋放CO2的再固定
PEPCase能夠催化植物體內呼吸釋放CO2的再固定(CO2refixation),該過程伴隨碳同位素判別效應(5.7‰)[36],對植物非光合組織呼吸釋放CO2碳同位素組成有重要影響。所有植物器官中均存在PEPCase[51-52],不同器官PEPCase酶活性的差異會導致同位素呼吸分餾上存在差異[53]。由于PEPCase的作用使得生成的有機物質中13C豐度高于未被固定CO2的豐度,同時降低了由植物向大氣的CO2呼吸釋放通量。理論上PEPCase酶活性與植物δ13CR值呈正相關關系,但研究表明植物根、莖的PEPCase與δ13CR值并沒有相關性[23]。因此,PEPCase可能并不是導致植物δ13CR變化的關鍵因素。
2.3 光照增強暗呼吸
當植物葉片從光照條件立即轉變為黑暗條件時,呼吸釋放CO2量呈顯著增加趨勢,這種現象被稱為光照增強暗呼吸(LEDR)[54]。LEDR不僅導致植物葉片呼吸釋放CO2量的增加,也影響呼吸釋放CO2的碳同位素組成,δ13CR值表現為先增加后降低的趨勢,植物葉片δ13CR的日變化幅度增加,δ13CR值增加的時間一般會持續5—20分鐘[55-56]。LEDR現象對葉片暗呼吸釋放CO2碳同位素組成影響程度依賴于光照強度,一般隨著光強的增加而增加[54, 57-59]。在光照條件下,植物自養器官中與糖酵解和檸檬酸循環相關酶的活性受到抑制[60-61],導致檸檬酸循環處于非閉合的狀態[62-63],而蘋果酸則在PEPCase的作用下得到積累。當植物葉片由光照條件轉入黑暗條件后,呼吸酶抑制作用解除,催化蘋果酸分解,釋放富含13C的C- 4,導致呼吸釋放CO2的碳同位素組成改變,LEDR的瞬時效應在自然條件下不僅出現在白天,晴天日落后也會持續約90分鐘,顯著影響夜晚初期生態系統呼吸釋放CO2同位素組成[64]。因此,采用容器收集植物葉片呼吸時需要慎重確定培養時間[3]。
2.4 環境因素對植物δ13CR短期變化的影響
環境因素能夠影響植物δ13CR及其變化幅度。DeNiro和Epstein[65]的研究發現夜間溫度的變化會通過影響呼吸酶的活性改變13C/12C分餾,促使葉片δ13CR發生變化。隨著葉片溫度的升高,呼吸底物由碳水化合物轉為脂肪(13C相對貧化),導致植物葉片呼吸CO2的δ13CR值持續下降[47]。Schnyder和Lattanzi[66]的研究也表明生長在高溫環境中(25℃/23℃,白天/夜晚)的黑麥草(LoliumperenneL.)其根系呼吸底物和呼吸釋放CO2的δ13C值(-21.7‰、-24.9‰)均高于生長在低溫環境中的δ13C值(15℃/14℃,白天/夜晚)(-22.8‰、-28.3‰)。
水分條件會通過影響光合碳同位素判別,改變呼吸底物的δ13C值,使植物呼吸CO2的δ13CR發生改變[67]。葉片內外水蒸汽壓差的增加通常會降低氣孔導度[68],使大氣CO2經氣孔進入葉片的量減少,導致胞間CO2和空氣CO2分壓比(Pi/Pa)降低,同位素判別會隨著Pi/Pa的降低而降低,同時增加了13C的同化,因此植物呼吸CO2的δ13CR值和日變化幅度均會發生改變[36]。在干旱脅迫下,水分條件的限制降低了氣孔導度和Pi/Pa的日變化幅度,導致光合碳同位素判別日變化幅度不明顯[69],并且干旱脅迫會顯著的降低呼吸分餾,影響與呼吸底物相關聯的植物呼吸δ13CR值[47-48]。例如,在美國亞利桑那州沙漠生態系統的研究結果表明,干旱季節葉片呼吸δ13CR值顯著高于雨季[32]。
不同的生長環境也會影響植物δ13CR值,Klumpp等[50]研究發現,生長在低密度下的向日葵δ13CR值高于生長在高密度下的δ13CR值。環境因素對植物呼吸δ13CR值的影響往往表現為綜合作用,不同時間尺度(晝夜、季節性)環境變化會改變與呼吸底物相關聯的葉片暗呼吸δ13CR的數值和日變化程度[19,33]。例如,Sun等[70-71]通過探究不同植被類型(C3和C4)δ13CR對于環境變化的響應發現,季節性的環境變化不僅影響呼吸代謝底物碳同位素值,也能夠通過影響底物量進而影響呼吸代謝中間產物乙酰輔酶A的利用,最終改變δ13CR值和日變化幅度。此外,植物在受到來自自然條件的脅迫時(例如凋萎或衰老、長期處于黑暗條件),會發生呼吸底物的轉變[19],呼吸底物類型(可溶性糖、淀粉、脂類、氨基酸)間在碳同位素組成上存在差異,進而對δ13CR產生影響[35,49]。
3 結論與展望
植物δ13CR的短期動態變化能夠反映植物碳分配方式、碳代謝生理過程、植物與環境相互作用等重要生理生態過程,探究植物呼吸釋放碳同位素組成變化有助于推進陸地生態系統碳循環的研究。不同植物類型以及植物的不同器官δ13CR值均存在差異。多數研究表明植物不同部位δ13CR值差異及變化幅度趨勢一致,表現為:葉片δ13CR>根系δ13CR>樹干/莖δ13CR,但植物暗呼吸釋放的δ13CR與呼吸底物的變化趨勢并不一致。同位素效應、呼吸底物的供給和消耗、糖類分子13C的不均勻分布、碳代謝相關酶的活性、LEDR、植物的遺傳特性及外部環境等因素均可以改變植物δ13CR值及其變化幅度。總體上說,導致植物δ13CR發生變化的原因可以歸納為以下幾點:(1)植物呼吸底物的δ13C值發生變化;(2)植物在不同時期利用的呼吸底物不同,而這些呼吸底物中δ13C值存在差異;(3)植物呼吸代謝中對中間產物利用方式的變化,導致植物δ13CR值發生變化。
近年來,隨著科學技術的發展,為研究植物暗呼吸碳同位素組成的動態變化及內在控制機制提供了廣闊的前景。隨著穩定性碳同位素標記技術的成熟,利用13C標記進而追蹤植物同化和釋放的碳同位素,可直接、有效的定期監測植物不同器官及其暗呼吸碳同位素組成變化趨勢及對環境變化的響應。穩定性同位素質譜儀(Isotope Ratio Mass Spectrometer, IRMS)是測定植物暗呼吸釋放CO2碳同位素組成的較為成熟的方法,具有樣品用量少、測量精確性高等優點。但設備存在結構復雜,體積大,造價較高,且測試樣品容易受到污染等缺陷。激光吸收光譜法(Laser Absorption Spectroscopy, LAS)、特定化合物同位素分析(Compound Specific Isotope Analysis, CSIA)、核磁共振技術(Nuclear Magnetic Resonance, NMR)、納米二次離子質譜技術(Nano-scale Secondary Ion Mass Spectrometer, Nano SIMS)具有快速測定、高敏感度、高精確度、增加可重復性等優點,為研究植物暗呼吸和內在控制機制提供了更為多樣化的手段。
關于導致植物暗呼吸發生變化的原因已有了較多的研究,但內在調控機理尚不明晰。在未來的研究中,研究者可關注以下幾方面的研究:(1)植物暗呼吸δ13CR值對外部環境變化及植物特性(例如:葉肉細胞導度)的響應;(2)植物同化產物δ13C值與呼吸釋放δ13CR的比較及其在不同時空尺度的動態變化;(3)不同功能群植物暗呼吸δ13CR的變化及其與呼吸代謝活動的關系;(4)不同生態系統碳同位素通量以及對生態系統呼吸δ13CR的貢獻率。
研究植物δ13CR有助于我們更好地了解植物尺度碳的流動,以及植物與生態系統的碳交換。目前,國際上已有較多關于δ13CR值短期變化的研究,但我國關于此方面的研究鮮有報道,以期通過本文增進國內關于植物呼吸碳同位素領域研究的了解,推動相關研究工作的深入開展。
參考文獻(References):
[1] Falkowski P, Scholes R J, Boyle E, Canadell J, Canfield D, Elser J, Gruber N, Hibbard K, H?gberg P, Linder S, Mackenzie F T, Moore III B, Pedersen T, Rosenthal Y, Seitzinger S, Smetacek V, Steffen W. The global carbon cycle: a test of our knowledge of earth as a system. Science, 2000, 290(5490): 291- 296.
[2] Scholze M, Kaminski T, Knorr W, Blessing S, Vossbeck M, Grant J P, Scipal K. Simultaneous assimilation of SMOS soil moisture and atmospheric CO2in-situ observations to constrain the global terrestrial carbon cycle. Remote Sensing of Environment, 2016, 180: 334- 345.
[3] Priault P, Wegener F, Werner C. Pronounced differences in diurnal variation of carbon isotope composition of leaf respired CO2among functional groups. New Phytologist, 2009, 181(2): 400- 412.
[4] IPCC. Climate Change 2013: the Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press, 2013.
[5] Yakir D, da Silveira Lobo Sternberg L. The use of stable isotopes to study ecosystem gas exchange. Oecologia, 2000, 123(3): 297- 311.
[6] Ostle N, Ineson P, Benham D, Sleep D. Carbon assimilation and turnover in grassland vegetation using aninsitu13CO2pulse labelling system. Rapid Communications in Mass Spectrometry, 2000, 14(15): 1345- 1350.
[7] 易現峰, 張曉愛. 穩定性同位素技術在生態學上的應用. 生態學雜志, 2005, 24(3): 306- 314.
[8] Bowling D R, Pataki D E, Randerson J T. Carbon isotopes in terrestrial ecosystem pools and CO2fluxes. New Phytologist, 2008, 178(1): 24- 40.
[9] Beer C, Reichstein M, Tomelleri E, Ciais P, Jung M, Carvalhais N, R?denbeck C, Arain M A, Baldocchi D, Bonan G B, Bondeau A, Cescatti A, Lasslop G, Lindroth A, Lomas M, Luyssaert S, Margolis H, Oleson K W, Roupsard O, Veenendaal E, Viovy N, Williams C, Woodward F I, Papale D. Terrestrial gross carbon dioxide uptake: global distribution and covariation with climate. Science, 2010, 329(5993): 834- 838.
[10] 林光輝. 穩定同位素生態學. 北京: 高等教育出版社, 2013.
[11] Fung I, Field C B, Berry J A, Thompson M V, Randerson J T, Malmstr?m C M, Vitousek P M, Collatz G J, Sellers P J, Randall D A, Denning A S, Badeck F, John J. Carbon 13 exchanges between the atmosphere and biosphere. Global Biogeochemical Cycles, 1997, 11(4): 507- 533.
[12] Pataki D E, Ehleringer J R, Flanagan L B, Yakir D, Bowling D R, Still C J, Buchmann N, Kaplan J O, Berry J A. The application and interpretation of Keeling plots in terrestrial carbon cycle research. Global Biogeochemical Cycles, 2003, 17(1): 1022.
[13] Mortazavi B, Chanton J P, Smith M C. Influence of13C-enriched foliage respired CO2onδ13C of ecosystem-respired CO2. Global Biogeochemical Cycles, 2006, 20(3): GB3029.
[14] Ghashghaie J, Badeck F W. Opposite carbon isotope discrimination during dark respiration in leaves versus roots-a review. New Phytologist, 2014, 201(3): 751- 769.
[15] Ghashghaie J, Badeck F W, Lanigan G, Nogués S, Tcherkez G, Deléens E, Cornic G, Griffiths H. Carbon isotope fractionation during dark respiration and photorespiration in C3plants. Phytochemistry Reviews, 2003, 2(1/2): 145- 161.
[16] Knohl A, Werner R A, Brand W A, Buchmann N. Short-term variations inδ13C of ecosystem respiration reveals link between assimilation and respiration in a deciduous forest. Oecologia, 2005, 142(1): 70- 82.
[17] Kodama N, Barnard R L, Salmon Y, Weston C, Ferrio J P, Holst J, Werner R A, Saurer M, Rennenberg H, Buchmann N, Gessler A. Temporal dynamics of the carbon isotope composition in aPinussylvestrisstand: from newly assimilated organic carbon to respired carbon dioxide. Oecologia, 2008, 156(4): 737- 750.
[18] Bowling D R, Tans P P, Monson R K. Partitioning net ecosystem carbon exchange with isotopic fluxes of CO2. Global Change Biology, 2001, 7(2): 127- 145.
[19] Unger S, Máguas C, Pereira J S, Aires L M, David T S, Werner C. Disentangling drought-induced variation in ecosystem and soil respiration using stable carbon isotopes. Oecologia, 2010, 163(4): 1043- 1057.
[20] Damesin C, Lelarge C. Carbon isotope composition of current-year shoots fromFagussylvaticain relation to growth, respiration and use of reserves. Plant, Cell & Environment, 2003, 26(2): 207- 219.
[21] Keitel C, Adams M A, Holst T, Matzarakis A, Mayer H, Rennenberg H, Ge?ler A. Carbon and oxygen isotope composition of organic compounds in the phloem sap provides a short-term measure for stomatal conductance of European beech (FagussylvaticaL.). Plant, Cell & Environment, 2003, 26(7): 1157- 1168.
[22] Pate J, Arthur D.δ13C analysis of phloem sap carbon: novel means of evaluating seasonal water stress and interpreting carbon isotope signatures of foliage and trunk wood ofEucalyptusglobulus. Oecologia, 1998, 117(3): 301- 311.
[23] Werner C, Gessler A. Diel variations in the carbon isotope composition of respired CO2and associated carbon sources: a review of dynamics and mechanisms. Biogeosciences, 2011, 8(9): 2437- 2459.
[24] Cui H Y, Wang Y B, Jiang Q, Chen S P, Ma J Y, Sun W. Carbon Isotope composition of nighttime leaf-respired CO2in the agricultural-pastoral zone of the Songnen Plain, Northeast China. PLoS One, 2015, 10(9): e0137575.
[25] Brandes E, Kodama N, Whittaker K, Weston C, Rennenberg H, Keitel C, Adams M A, Gessler A. Short-term variation in the isotopic composition of organic matter allocated from the leaves to the stem ofPinussylvestris: effects of photosynthetic and postphotosynthetic carbon isotope fractionation. Global Change Biology, 2006, 12(10): 1922- 1939.
[26] Brandes E, Wenninger J, Koeniger P, Schindler D, Rennenberg H, Leibundgut C, Mayer H, Gessler A. Assessing environmental and physiological controls over water relations in a Scots pine (PinussylvestrisL.) stand through analyses of stable isotope composition of water and organic matter. Plant, Cell & Environment, 2007, 30(1): 113- 127.
[27] Gessler A, Keitel C, Kodama N, Weston C, Winters A J, Keith H, Grice K, Leuning R, Farquhar G D.δ13C of organic matter transported from the leaves to the roots inEucalyptusdelegatensis: short-term variations and relation to respired CO2. Functional Plant Biology, 2007, 34(8): 692- 706.
[28] Gessler A, Tcherkez G, Peuke A D, Ghashghaie J, Farquhar G D. Experimental evidence for diel variations of the carbon isotope composition in leaf, stem and phloem sap organic matter inRicinuscommunis. Plant, Cell & Environment, 2008, 31(7): 941- 953.
[29] Werner C, Wegener F, Unger S, Nogués S, Priault P. Short-term dynamics of isotopic composition of leaf-respired CO2upon darkening: measurements and implications. Rapid Communications in Mass Spectrometry, 2009, 23(16): 2428- 2438.
[30] Wegener F, Beyschlag W, Werner C. The magnitude of diurnal variation in carbon isotopic composition of leaf dark respired CO2correlates with the difference betweenδ13C of leaf and root material. Functional Plant Biology, 2010, 37(9): 849- 858.
[31] Rascher K G, Máguas C, Werner C. On the use of phloem sapδ13C as an indicator of canopy carbon discrimination. Tree Physiology, 2010, 30(12): 1499- 1514.
[32] Sun W, Resco V, Williams D G. Diurnal and seasonal variation in the carbon isotope composition of leaf dark-respired CO2in velvet mesquite (Prosopisvelutina). Plant, Cell & Environment, 2009, 32(10): 1390- 1400.
[33] Hymus G J, Maseyk K, Valentini R, Yakir D. Large daily variation in13C-enrichment of leaf-respired CO2in twoQuercusforest canopies. New Phytologist, 2005, 167(2): 377- 384.
[34] Gessler A, Tcherkez G, Karyanto O, Keitel C, Ferrio J P, Ghashghaie J, Kreuzwieser J, Farquhar G D. On the metabolic origin of the carbon isotope composition of CO2evolved from darkened light-acclimated leaves inRicinuscommunis. New Phytologist, 2009, 181(2): 374- 386.
[35] Nogués S, Tcherkez G, Cornic G, Ghashghaie J. Respiratory carbon metabolism following illumination in intact French bean leaves using13C/12C isotope labeling. Plant Physiology, 2004, 136(2): 3245- 3254.
[36] Farquhar G D, Ehleringer J R, Hubick K T. Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology, 1989, 40: 503- 537.
[37] 林光輝, 柯淵. 穩定同位素技術與全球變化研究//李博. 現代生態學講座. 北京: 科學出版社, 1995.
[38] Zeeman S C, Smith S M, Smith A M. The diurnal metabolism of leaf starch. Biochemical Journal, 2007, 401(1): 13- 28.
[39] Cernusak L A, Tcherkez G, Keitel C, Cornwell W K, Santiago L S, Knohl A, Barbour M M, Williams D G, Reich P B, Ellsworth D S, Dawson T E, Griffiths H G, Farquhar G D, Wright I J. Why are non-photosynthetic tissues generally13C enriched compared with leaves in C3plants? Review and synthesis of current hypotheses. Functional Plant Biology, 2009, 36(3): 199- 213.
[40] Tcherkez G, Farquhar G, Badeck F, Ghashghaie J. Theoretical considerations about carbon isotope distribution in glucose of C3plants. Functional Plant Biology, 2004, 31(9): 857- 877.
[41] Werner C, Schnyder H, Cuntz M, Keitel C, Zeeman M J, Dawson T E, Badeck F W, Brugnoli E, Ghashghaie J, Grams T E E, Kayler Z E, Lakatos M, Lee X, Máguas C, Ogée J, Rascher K G, Siegwolf R T W, Unger S, Welker J, Wingate L, Gessler A. Progress and challenges in using stable isotopes to trace plant carbon and water relations across scales. Biogeosciences, 2012, 9(8): 3083- 3111.
[42] Gleixner G, Scrimgeour C, Schmidt H L, Viola R. Stable isotope distribution in the major metabolites of source and sink organs ofSolanumtuberosumL.: a powerful tool in the study of metabolic partitioning in intact plants. Planta, 1998, 207(2): 241- 245.
[43] Gleixner G, Schmidt H L. Carbon isotope effects on the fructose- 1,6-bisphosphate aldolase reaction, origin for non-statistical13C distributions in carbohydrates. Journal of Biological Chemistry, 1997, 272(9): 5382- 5387.
[44] Rossmann A, Butzenlechner M, Schmidt H L. Evidence for a nonstatistical carbon isotope distribution in natural glucose. Plant Physiology, 1991, 96(2): 609- 614.
[45] Hobbie E A, Werner R A. Intramolecular, compound-specific, and bulk carbon isotope patterns in C3and C4plants: a review and synthesis. New Phytologist, 2004, 161(2): 371- 385.
[46] Park R, Epstein S. Metabolic fractionation of C13& C12in plants. Plant Physiology, 1961, 36(2): 133- 138.
[47] Duranceau M, Ghashghaie J, Badeck F, Deleens E, Cornic G.δ13C of CO2respired in the dark in relation toδ13C of leaf carbohydrates inPhaseolusvulgarisL. under progressive drought. Plant, Cell & Environment, 1999, 22(5): 515- 523.
[48] Ghashghaie J, Duranceau M, Badeck F W, Cornic G, Adeline M T, Deleens E.δ13C of CO2respired in the dark in relation toδ13C of leaf metabolites: comparison betweenNicotianasylvestrisandHelianthusannuusunder drought. Plant, Cell & Environment, 2001, 24(5): 505- 515.
[49] Tcherkez G, Nogués S, Bleton J, Cornic G, Badeck F, Ghashghaie J. Metabolic origin of carbon isotope composition of leaf dark-respired CO2in French bean. Plant Physiology, 2003, 131(1): 237- 244.
[50] Klumpp K, Sch?ufele R, L?tscher M, Lattanzi F A, Feneis W, Schnyder H. C-isotope composition of CO2respired by shoots and roots: fractionation during dark respiration? Plant, Cell & Environment, 2005, 28(2): 241- 250.
[51] Hibberd J M, Quick W P. Characteristics of C4photosynthesis in stems and petioles of C3flowering plants. Nature, 2002, 415(6870): 451- 454.
[52] Berveiller D, Damesin C. Carbon assimilation by tree stems: potential involvement of phosphoenolpyruvate carboxylase. Trees, 2008, 22(2): 149- 157.
[53] Badeck F W, Tcherkez G, Nogués S, Piel C, Ghashghaie J. Post-photosynthetic fractionation of stable carbon isotopes between plant organs-a widespread phenomenon. Rapid Communications in Mass Spectrometry, 2005, 19(11): 1381- 1391.
[54] Atkin O K, Evans J R, Ball M C, Siebke K, Pons T L, Lambers H. Light inhibition of leaf respiration: the role of irradiance and temperature//Moller I M, Gardestrom P, Gliminius K, Glaser E, eds. Plant Mitochondria: from Gene to Function. Leiden: Backhuys Publishers, 1998: 567- 574.
[55] Barbour M M, McDowell N G, Tcherkez G, Bickford C P, Hanson D T. A new measurement technique reveals rapid post-illumination changes in the carbon isotope composition of leaf-respired CO2. Plant, Cell & Environment, 2007, 30(4): 469- 482.
[56] Werner C, Hasenbein N, Maia R, Beyschlag W, Máguas C. Evaluating high time-resolved changes in carbon isotope ratio of respired CO2by a rapid in-tube incubation technique. Rapid Communications in Mass Spectrometry, 2007, 21(8): 1352- 1360.
[57] Reddy M M, Vani T, Raghavendra A S. Light-enhanced dark respiration in mesophyll protoplasts from leaves of pea. Plant Physiology, 1991, 96(4): 1368- 1371.
[58] Xue X, Gauthier D A, Turpin D H, Weger H G. Interactions between photosynthesis and respiration in the green algaChlamydomonasreinhardtii(characterization of light-enhanced dark respiration). Plant Physiology, 1996, 112(3): 1005- 1014.
[59] Atkin O K, Evans J R, Siebke K. Relationship between the inhibition of leaf respiration by light and enhancement of leaf dark respiration following light treatment. Australian Journal of Plant Physiology, 1998, 25(4): 437- 443.
[60] Tcherkez G, Farquhar G D. Carbon isotope effect predictions for enzymes involved in the primary carbon metabolism of plant leaves. Functional Plant Biology, 2005, 32(4): 277- 291.
[61] Nunes-Nesi A, Sweetlove L J, Fernie A R. Operation and function of the tricarboxylic acid cycle in the illuminated leaf. Physiologia Plantarum, 2007, 129(1): 45- 56.
[62] Tcherkez G, Mahé A, Gauthier P, Mauve C, Gout E, Bligny R, Cornic G, Hodges M. In folio respiratory fluxomics revealed by13C isotopic labeling and H/D isotope effects highlight the noncyclic nature of the tricarboxylic acid “cycle” in illuminated leaves. Plant Physiology, 2009, 151(2): 620- 630.
[63] Sweetlove L J, Beard K F M, Nunes-Nesi A, Fernie A R, Ratcliffe R G. Not just a circle: flux modes in the plant TCA cycle. Trends in Plant Science, 2010, 15(8): 462- 470.
[64] Barbour M M, Hunt J E, Kodama N, Laubach J, McSeveny T M, Rogers G N D, Tcherkez G, Wingate L. Rapid changes inδ13C of ecosystem-respired CO2after sunset are consistent with transient13C enrichment of leaf respired CO2. New Phytologist, 2011, 190(4): 990- 1002.
[65] DeNiro M J, Epstein S. Mechanism of carbon isotope fractionation associated with lipid synthesis. Science, 1977, 197(4300): 261- 263.
[66] Schnyder H, Lattanzi F A. Partitioning respiration of C3-C4 mixed communities using the natural abundance13C approach-testing assumptions in a controlled environment. Plant Biology, 2005, 7(6): 592- 600.
[67] Brugnoli E, Farquhar G D. Photosynthetic fractionation of carbon isotopes//Leegood R C, Sharkey T D, von Caemmerer S, eds. Photosynthesis: Physiology and Metabolism. Berlin: Springer, 2000: 399- 434.
[68] Oren R, Sperry J S, Katul G G, Pataki D E, Ewers B E, Phillips N, Sch?fer K V R. Survey and synthesis of intra-and interspecific variation in stomatal sensitivity to vapour pressure deficit. Plant, Cell & Environment, 1999, 22(12): 1515- 1526.
[69] Valladares F, Pearcy R W. Interactions between water stress, sun-shade acclimation, heat tolerance and photoinhibition in the sclerophyllHeteromelesarbutifolia. Plant, Cell & Environment, 1997, 20(1): 25- 36.
[70] Sun W, Resco V, Williams D G. Environmental and physiological controls on the carbon isotope composition of CO2respired by leaves and roots of a C3woody legume (Prosopisvelutina) and a C4perennial grass (Sporoboluswrightii). Plant, Cell & Environment, 2012, 35(3): 567- 577.
[71] Sun W, Resco V, Williams D G. Nocturnal and seasonal patterns of carbon isotope composition of leaf dark-respired carbon dioxide differ among dominant species in a semiarid savanna. Oecologia, 2010, 164(2): 297- 310.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
