沉默β-catenin表達對胃癌MGC-803細胞生物學行為的影響與作用機制研究
2018-06-14 03:00:14潘安萍朱建偉王忻妍
浙江醫學 2018年9期
潘安萍 朱建偉 王忻妍
β-鏈蛋白(β-catenin)是一種多功能的蛋白質。在正常細胞內,β-catenin多數位于細胞膜,與E鈣黏蛋白及α-catenin一起在細胞間的黏附中發揮關鍵作用[1]。β-catenin若脫離細胞膜進入細胞質,易被降解,但當β-catenin在細胞內發生降解異常時,β-catenin會在細胞內堆積,進入細胞核促進下游靶基因的轉錄,即β-catenin的另一重要功能-參與Wnt信號通路[2]。β-catenin下游靶基因編碼許多調節細胞生物學行為的相關蛋白,如 c-Myc、cyclin D1、Bcl-2、Bax、基質金屬蛋白酶(MMP)-7等,即β-catenin作為轉錄因子有可能影響細胞增殖、分化、侵襲、轉移等多種生物學行為,且有研究發現β-catenin在多種腫瘤細胞中明顯高表達[3]。基于此,本研究應用干擾慢病毒感染β-catenin相對高表達的胃癌細胞株MGC-803,沉默其細胞內β-catenin表達,而后通過多種實驗方法觀察β-catenin表達下調對MGC-803細胞增殖、凋亡、侵襲、轉移等生物學行為的影響,同時檢測沉默β-catenin表達后的MGC-803細胞中Bcl-2、Bax、cyclin D1和MMP-7等蛋白表達水平的變化,初步探討β-catenin下調影響胃癌MGC-803細胞生物學行為可能的作用機制,現報道如下。
1 材料和方法
1.1 細胞與主要試劑 MGC-803細胞株購自上海博谷生物科技有限公司。β-catenin-RNAi慢病毒液、βcatenin-Neg慢病毒液購于上海吉瑪公司;凋亡檢測AnnexinV-PE/7-AAD試劑盒、細胞周期碘化丙啶(PI)染色試劑盒購于南京凱基生物有限公司;Matrigel膠購于美國BD公司;兔抗人β-catenin單克隆抗體購于美國Santan Cruz公司;鼠抗人β-actin、羊抗鼠IgG-HRP、羊抗兔IgG-HRP單克隆抗體均購于中杉金橋公司。
1.2 方法
1.2.1 實驗分組 分別將購買的β-catenin-RNAi慢病毒及β-catenin-Neg慢病毒感染MGC-803細胞,實驗分組:實驗組(感染β-catenin-RNAi慢病毒的MGC-803-RNAi細胞)、陰性對照組(感染空載慢病毒的MGC-803-Neg細胞)、空白對照組(未處理的MGC-803細胞)。
1.2.2 病毒感染與細胞培養 根據預實驗得知該病毒對MGC-803細胞的感染復數約為100,將MGC-803細胞接種于6孔板,待細胞貼壁后實驗組與陰性對照組細胞分別滴加相應慢病毒液,空白對照組加等體積培養基。24h后換液,3d后熒光倒置顯微鏡下觀察細胞熒光染色情況。感染效率=帶綠色熒光細胞數/總細胞數×100%。
1.2.3 Western blot檢測細胞 β-catenin、Bcl-2、Bax、cyclin D1及MMP-7表達水平 用蛋白提取試劑盒提取各組細胞內的總蛋白,分別取20μl進行聚丙烯凝膠電泳,結束后目的蛋白條帶轉移至硝酸纖維素(NC)膜,5%的脫脂奶粉封閉1h,一抗(1:1 000)4℃封閉過夜,二抗(1:2 500)室溫封閉1h。暗室內用發光液進行曝光、顯影、定影,得到目的蛋白條帶的膠片,用Image J軟件檢測各組細胞 β-catenin、Bcl-2、Bax、cyclin D1 及 MMP-7相對表達水平。實驗重復3次取平均值。
1.2.4 Annexin-V-PE/7-AAD雙染流式細胞術檢測各組細胞凋亡率 將各組細胞分別接種于6cm培養皿常規培養。待細胞長至約80%~90%融合度時收集細胞,PBS洗滌2次,加入500 μl的Binding Buffer懸浮細胞,加入1 μl Annexin V-PE染液,室溫、避光、反應15min,加入5 μl 7-AAD染液,室溫、避光、反應15min。過300目尼龍網后1h內進行流式細胞術檢測細胞凋亡率。實驗重復3次取平均值。
1.2.5 PI單染流式細胞術檢測各組細胞周期 取各組細胞分別接種于6cm培養皿常規培養,待細胞長至約80%~90%融合度時收取細胞,每組細胞取約1×105個。用PBS清洗2次后用預冷的75%乙醇溶液1ml重懸細胞沉淀,4℃固定過夜。第2天1 000r/min離心5min后去固定液,PBS清洗2次,加入100μl的RNase A重懸細胞沉淀,置于37℃水浴鍋內30 min,加入PI染料400μl,4℃避光染色30min。過300目尼龍網后,流式細胞術檢測細胞周期。實驗重復3次取平均值。
1.2.6 Transwell法檢測細胞的侵襲能力 將融好的Mtrigel膠與含10%BSA的1640培養基按體積比1:6混合,后取50μl平鋪于小室的上室面。收集各組細胞,PBS清洗2次,用完全培養基稀釋成1×105/ml的細胞懸液。小室的下室面各加入含10%FBS的1640完全培養基500μl,上室面加入200μl各組細胞懸液,培養24h后取出小室,擦拭上室面未穿膜的細胞后結晶紫染色,顯微鏡下計數穿膜細胞數分析細胞侵襲能力。實驗重復3次取平均值。
1.3 觀察指標 觀察并比較MGC-803、MGC-803-Neg、MGC-803-RNAi細胞熒光染色情況、β-catenin 表達水平、細胞凋亡率、細胞周期、侵襲能力,以及Bcl-2、Bax、cyclin D1、MMP-7 蛋白表達水平。
1.4 統計學處理 應用SPSS 17.0統計軟件;計量資料以表示,3組比較采用單因素方差分析,兩兩比較采用LSD-t檢驗;P<0.05為差異有統計學意義。
2 結果
2.1 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞熒光染色情況比較 感染4d后,熒光顯微鏡下可見MGC-803-Neg、MGC-803-RNAi細胞顯示強綠色熒光,且帶綠色熒光的細胞數占總細胞數比例均超過80%,而MGC-803細胞無綠色熒光,即MGC-803-Neg、MGC-803-RNAi細胞慢病毒感染效率>80%。MGC-803-RNAi細胞熒光染色情況見圖1(插頁)。
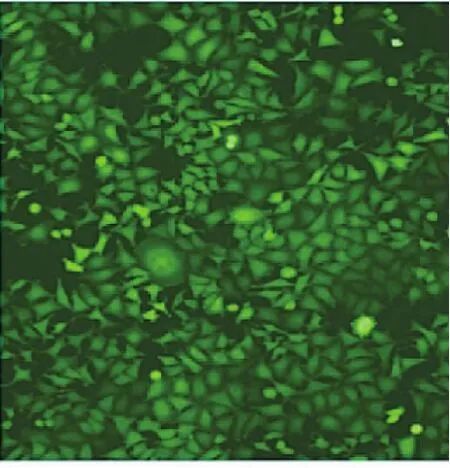
圖1 MGC-803-RNAi細胞熒光顯微鏡下所見(×100)
2.2 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞β-catenin表達水平比較 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞β-catenin相對表達水平分別為0.8725±0.0110、0.8593±0.0074、0.4220±0.0164,3 組比較差異有統計學意義(P>0.05),MGC-803-RNAi細胞β-catenin表達水平較MGC-803、MGC-803-Neg細胞下調(均P<0.05),而MGC-803-Neg細胞與MGC-803細胞間比較無統計學差異(P>0.05)。MGC-803、MGC-803-Neg、MGC-803-RNAi細胞 β-catenin蛋白電泳圖比較見圖2。

圖2 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞 β-catenin蛋白電泳圖比較(a:MGC-803 細胞;b:MGC-803-Neg細胞;c:MGC-803-RNAi細胞)
2.3 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞凋亡率比較 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞凋亡率分別為(1.20±0.26)%、(1.35±0.35)%、(8.52±0.82)%,3組比較差異有統計學意義(P>0.05),MGC-803-RNAi細胞凋亡率高于 MGC-803、MGC-803-Neg細胞(均P<0.05),而 MGC-803-Neg細胞與MGC-803細胞間比較無統計學差異(P>0.05)。流式細胞術檢測 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞凋亡率比較見圖3。
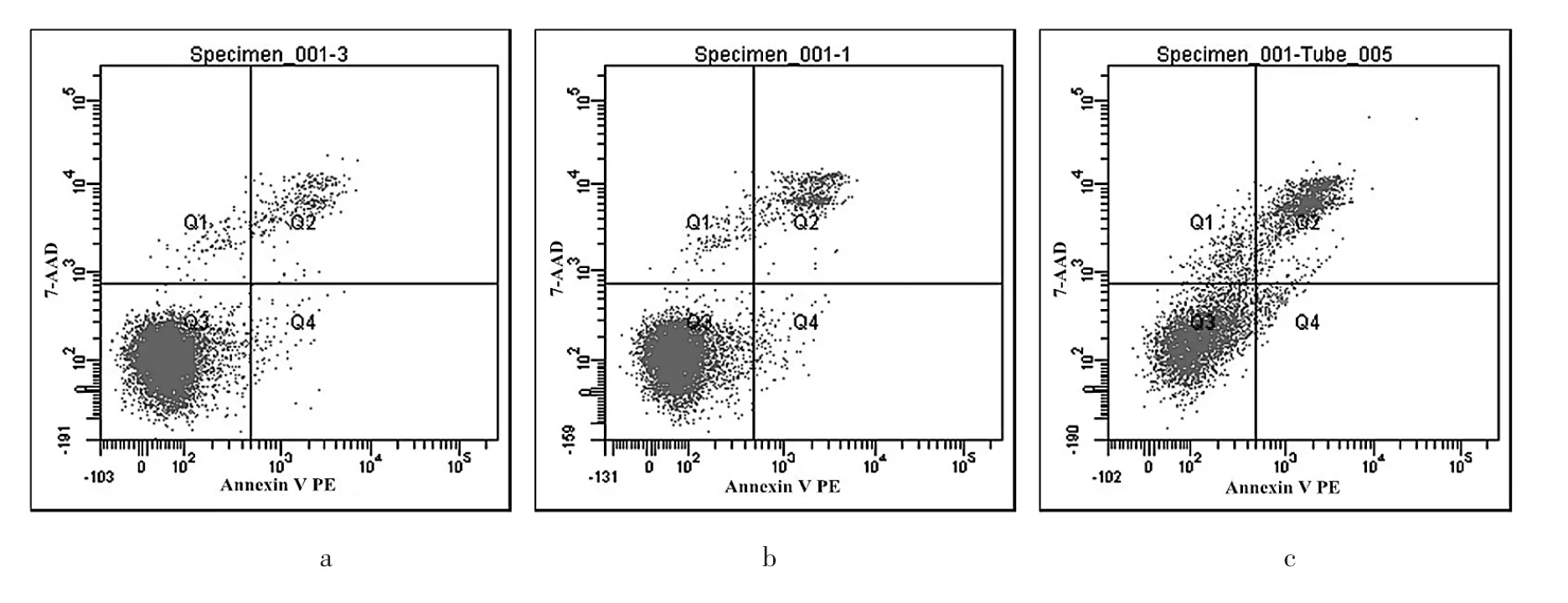
圖3 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞凋亡率比較(a:MGC-803 細胞;b:MGC-803-Neg細胞;c:MGC-803-RNAi細胞)
2.4 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞周期比較 以S/G1期細胞比例為觀察指標,MGC-803、MGC-803-Neg、MGC-803-RNAi細胞分別為 (1.11±0.05)%、(1.09±0.10)%、(0.78±0.08)%,3 組比較差異有統計學意義(P >0.05),MGC-803-RNAi細胞 S/G1期比例低于 MGC-803、MGC-803-Neg細胞(均P<0.05),而MGC-803-Neg細胞與MGC-803細胞間比較無統計學差異(P>0.05)。流式細胞術檢測MGC-803、MGC-803-Neg、MGC-803-RNAi細胞周期結果比較見圖4(插頁)。

圖4 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞周期結果比較(a:MGC803 細胞;b:MGC803-Neg細胞;c:MGC803-RNAi細胞)
2.5 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞侵襲能力比較 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞穿膜染色數分別為(109.80±5.36)、(104.40±7.47)、(54.00±6.28)個,3 組比較差異有統計學意義(P>0.05),MGC-803-RNAi細胞侵襲能力低于 MGC-803、MGC-803-Neg細胞(均P<0.05),而 MGC-803-Neg細胞與MGC-803細胞間比較無統計學差異(P>0.05)。Transwell法檢測 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞侵襲能力實驗結果見圖5(插頁)。

圖5 MGC-803、MGC-803-Neg、MGC-803-RNAi穿膜細胞顯微鏡下所見比較(a:MGC803 細胞;b:MGC803-Neg 細胞;c:MGC803-RNAi細胞;結晶紫染色,×200)
2.6 MGC-803、MGC-803-Neg、MGC-803-RNAi 細胞 Bcl-2、Bax、cyclin D1及MMP-7蛋白表達水平比較 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞Bcl-2、Bax、cyclin D1及 MMP-7蛋白表達水平比較差異均有統計學意義(均P>0.05),MGC-803-RNAi細胞Bcl-2、cyclin D1、MMP-7蛋白表達水平均低于MGC-803、MGC-803-Neg細胞(均P<0.05),Bax蛋白表達水平高于 MGC-803、MGC-803-Neg細胞(均P<0.05),而MGC-803-Neg細胞與MGC-803細胞間比較無統計學差異(P >0.05)。MGC-803、MGC-803-Neg、MGC-803-RNAi細胞 Bcl-2、Bax、cyclin D1 及 MMP-7 蛋白電泳圖比較見圖6。
3 討論
β-catenin是一種多功能、進化保守的蛋白質分子,在多細胞生物的胚胎發育及機體穩態中發揮至關重要的作用。研究發現,在腫瘤的發生、發展過程中,一方面β-catenin作為一種轉錄活化因子,能介導多種原癌基因與細胞周期調控基因的轉錄翻譯,促進細胞增殖、侵襲及轉移,抑制細胞分化、凋亡,還能誘導腫瘤耐藥,另一方面,腫瘤細胞中β-catenin在細胞質內的異常定位導致細胞膜上β-catenin含量下降,易使癌細胞間黏附功能下降,致腫瘤細胞更易于脫離原發部位而發生遠處轉移[3]。β-catenin參與調控上述生物學行為的靶基因編碼蛋白包含 c-Myc、cyclin D1、Bcl-2、Bax、MMP-7、P-gp等[3]。本研究選取β-catenin相對高表達的胃癌MGC-803細胞進行實驗,通過干擾慢病毒感染MGC-803細胞,經感染成功的MGC-803細胞在熒光顯微鏡下有明顯的熒光顯示;并通過Western blot法證實MGC-803-RNAi細胞β-catenin表達水平明顯下調。本研究再將此穩定穿代的3株細胞(MGC-803-RNAi、MGC-803-Neg及MGC-803細胞)進行后續實驗,以了解βcatenin表達下調對MGC-803細胞生物學行為的影響及其可能的作用機制。

圖6 MGC-803、MGC-803-Neg、MGC-803-RNAi細胞 Bcl-2、Bax、cyclin D1及 MMP-7蛋白電泳圖比較(a:MGC-803細胞;b:MGC-803-Neg細胞;c:MGC-803-RNAi細胞)
本研究采用Annexin-V-PE/7-AAD雙染流式細胞術證實β-catenin表達的下調可促進MGC-803細胞凋亡。這與在腸癌細胞中得到的結果一致[4]。細胞凋亡過程主要受Bcl-2蛋白家族所調控,其中最具代表性的為Bax及Bcl-2蛋白,Bax促進細胞凋亡,而Bcl-2抑制細胞凋亡[5]。本研究檢測到沉默β-catenin表達之后的MGC-803-RNAi細胞Bcl-2表達水平明顯下調,而Bax表達水平則相對上調,結果同預期。筆者推測,β-catenin表達異常對MGC-803細胞凋亡的影響,其機制與影響Wnt/β-catenin信號通路下游靶蛋白Bax及Bcl-2的表達有關。cyclin是一類在細胞周期中發揮關鍵調節作用的蛋白,其中cyclin D1能調細胞周期由G1期順利進入到S期[6]。本研究干擾MGC-803細胞β-catenin的表達后發現MGC-803-RNAi細胞cyclin D1表達水平也下調,且出現明顯的G1期阻滯,細胞增殖減弱。這與Shtutman等[7]研究結果相符。筆者推論,沉默β-catenin表達使MGC-803細胞阻滯在G1期,其作用機制與Wnt/β-catenin信號通路下游靶蛋白cyclin D1表達下調有關。
胃癌細胞轉移是胃癌患者治療失敗的主要原因。β-catenin作為細胞黏附中的關鍵因子,當其在細胞膜中減少時,細胞間黏附功能下降,導致腫瘤細胞易脫離原發部位。但是游離出來的腫瘤細胞要在細胞間質中進行遷徙,必須能夠分解細胞外基質。在細胞外基質的眾酶中,MMP起著關鍵的作用[8]。研究顯示,MMP家族蛋白酶的表達受Wnt/β-catenin信號通路調控,其中MMP-7蛋白與腫瘤關系最密切,MMP-7普遍被發現在腫瘤組織及細胞中明顯高表達,且MMP-7在相對晚期的胃癌組織中陽性率明顯高于相對早期的胃癌組織[9]。本研究通過Transwell實驗方法發現,沉默β-catenin表達之后,MGC-803-RNAi細胞的侵襲力顯著下降,且MGC-803-RNAi細胞內MMP-7的表達水平也明顯下降。因此推論β-catenin沉默抑制MGC-803細胞侵襲能力與Wnt/β-catenin信號通路下游靶蛋白MMP-7表達水平下調有關。這一結果與在腸癌、食管癌中的研究相似[10]。
綜上所述,本研究結果顯示,β-catenin表達下調可影響胃癌MGC-803細胞的增殖、凋亡、侵襲等多種生物學行為,其作用機制可能與影響Wnt/β-catenin信號通路下游多種靶蛋白表達有關。
[1]Archbold HC,Yang YX,Chen L,et al.How do they do Wnt they do?:regulation of transcription by the Wnt/β-catenin pathway[J].Acta Physiologica,2012,204(1):74.
[2]Hecht A,Litterst CM,Huber O,et al.Functional characterization of multiple transactivating elements in beta-catenin,some of which interact with the 1'ATA-binding protein in vitro[J].J Biol Chem,1999,274(25):18017-18025.
[3]Polakis P.Wnt signaling and cancer[J].Genes&Development,2000,14(15):1837-1851.
[4]Lu W,Jia G,Meng X,et al.Beta-catenin mediates the apoptosis induction effect of celastrol in HT29 cells[J].Life sciences,2012,91(7):279-283.
[5]Hancock JT,Desikan R,Neill SJ.Role of reactive oxygen species in cell signalling pathways[J].Biochemical Society Transactions,2001,29(2):345-349.
[6]Brunelle JK,Letai A.Control of mitochondrial apoptosis by the Bcl-2 family[J].Journal of cell science,2009,122(4):437-441.
[7]Shtutman M,Zhurinsky J,Simcha I,et al.The cyclin D1 gene is a target of the β-catenin/LEF-1 pathway[J].Proceedings of the National A-cademyof Sciences,1999,96(10):5522-5527.
[8]Deryugina EI,Quigley JP.Matrix metalloproteinases and tumor metastasis[J].Cancer and Metastasis Reviews,2006,25(1):9-34.
[9]Brabletz T,Jung A,Dag S,et al.β-catenin regulates the expression of the matrix metalloproteinase-7 in human colorectal cancer[J].The American journal of pathology,1999,155(4):1033-1038.
[10]Minamoto T,Ougolkov A,Yamashita K,et al.Oncogenic beta-catenin and MMP-7(matrilysin)cosegregate in late stage clinical colon cancer[J].Proceedings of the American Association for Cancer Research,2004,2004(1):1170.
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
人大建設(2019年12期)2019-05-21 02:55:32
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
中國衛生標準管理(2015年3期)2016-01-14 03:41:46
醫學研究雜志(2015年9期)2015-07-01 17:28:27
中國當代醫藥(2015年20期)2015-03-01 02:04:29
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:52
