致病性小腸結(jié)腸炎耶爾森菌外膜蛋白A對(duì)SD大鼠免疫保護(hù)作用研究
2018-06-21 09:00:50,,,,,,,
中國(guó)人獸共患病學(xué)報(bào) 2018年6期
關(guān)鍵詞:劑量
, ,, , ,,,
小腸結(jié)腸炎耶爾森菌(Yersiniaenterocolitica)是一種人獸共患病原菌,感染這種病原體可引起廣泛的臨床表現(xiàn),如急性腸炎、小腸結(jié)腸炎、末端回腸炎和腸系膜淋巴結(jié)炎,嚴(yán)重者還可引發(fā)一系列后遺癥如肝脾膿腫、關(guān)節(jié)炎、結(jié)節(jié)性紅斑、耶爾森肝炎和敗血癥等疾病[1]。OmpA是鑲嵌在革蘭陰性菌外膜上的一種產(chǎn)量豐富的β-桶裝蛋白,該蛋白不僅參與生物膜的形成,還可作為噬菌體和細(xì)菌素的受體,對(duì)于細(xì)菌具有重要的生理學(xué)功能[2-3]。近年來(lái),越來(lái)越多的研究表明OmpA在鼠疫耶爾森菌、沙門(mén)菌屬、溶血性曼氏桿菌和鮑曼不動(dòng)桿菌中都具有良好的免疫原性[4-7],提示OmpA有可能成為研制這些病原菌疫苗的候選蛋白。本課題組前期研究工作中已發(fā)現(xiàn)ompA核酸序列在三種致病性耶爾森菌中高度保守,并且在25 ℃和37 ℃下均是主要的免疫原性蛋白之一,體外免疫雜交實(shí)驗(yàn)證明該蛋白具有明顯的交叉免疫原性,具備作為致病性耶爾森菌的交叉免疫保護(hù)性抗原的潛力[8-10]。本研究使用純化的OmpA蛋白作為免疫原對(duì)大鼠進(jìn)行免疫,通過(guò)體內(nèi)實(shí)驗(yàn)對(duì)OmpA的免疫保護(hù)作用進(jìn)行分析。
1 材料與方法
1.1實(shí)驗(yàn)菌株和實(shí)驗(yàn)動(dòng)物 小腸結(jié)腸炎耶爾森菌Ye92010(1b/O∶8)和Ye92010-ompA重組克隆子均來(lái)自本實(shí)驗(yàn)室。SD雄性大鼠14只,SPF級(jí),體重280~320 g,購(gòu)自北京大學(xué)醫(yī)學(xué)部實(shí)驗(yàn)動(dòng)物科學(xué)部。
1.2主要實(shí)驗(yàn)試劑 His蛋白純化試劑盒購(gòu)于Novagen公司, BCA蛋白定量試劑盒購(gòu)自北京康為世紀(jì)生物科技有限公司,IPTG購(gòu)于Amresco公司,弗氏不完全佐劑購(gòu)于Sigma公司,細(xì)菌基因組提取試劑盒購(gòu)于Qiagen公司,PBS(pH=7.4)購(gòu)于Gibco公司,改良?jí)埦?Peptone Sorbitol Bile Broth)購(gòu)于Fluka公司,O:8血清型分型血清購(gòu)自日本生研株式會(huì)社,PCR擴(kuò)增相關(guān)試劑購(gòu)于TaKaRa公司,引物合成由上海生工生物工程有限公司完成。
1.3Ye92010-ompA的表達(dá)與純化[10]將重組克隆子Ye92010-ompA接種于具kan抗性的LB液體培養(yǎng)基中,置于37 ℃搖床培養(yǎng)至OD600約為0.6~0.8時(shí)加入終濃度1 mmol/L的IPTG,28 ℃培養(yǎng)誘導(dǎo)8 h。使用PBS(pH=7.4)將菌體清洗3次后重懸于binding buffer中,超聲裂解,5 500 r/min 離心10 min收集沉淀。將沉淀溶于含6 mol/L尿素的binding buffer中,冰浴1 h,再轉(zhuǎn)入His層析柱對(duì)蛋白進(jìn)行純化。純化后的蛋白通過(guò)SDS-PAGE電泳和考馬斯亮藍(lán)染色進(jìn)行驗(yàn)證,使用BCA蛋白定量試劑盒通過(guò)Bradford法對(duì)純化后蛋白進(jìn)行濃度測(cè)定。
1.4OmpA免疫保護(hù)試驗(yàn) 實(shí)驗(yàn)動(dòng)物分組情況見(jiàn)表1,SD大鼠被隨機(jī)分為OmpA免疫組(6只)、對(duì)照組(6只)和空白對(duì)照組(2只)。免疫組使用OmpA純化蛋白與弗氏不完全佐劑混合后(1.5∶1)通過(guò)腹股溝注射方式對(duì)大鼠進(jìn)行免疫,對(duì)照組由等體積的PBS代替蛋白。首次免疫(0.58 mg/只)后14 d對(duì)大鼠進(jìn)行加強(qiáng)免疫(1 mg/只)。攻毒試驗(yàn)使用致病性小腸結(jié)腸炎耶爾森菌Ye92010(低劑量組:106CFU、高劑量組:108CFU)通過(guò)腹腔注射方式進(jìn)行。攻毒后開(kāi)始為期3周的連續(xù)觀(guān)察,每日測(cè)量大鼠肛溫并采集糞便。
表1 OmpA免疫保護(hù)試驗(yàn)動(dòng)物分組
Tab.1 Animal grouping of OmpA immunoprotection test

動(dòng)物分組和攻毒劑量OmpA免疫組對(duì)照組空白對(duì)照組低劑量攻毒高劑量攻毒低劑量攻毒高劑量攻毒未做任何處理數(shù)量/只33332
1.5糞便排菌實(shí)驗(yàn)及PCR檢測(cè) 將大鼠糞便稱(chēng)重后按1∶2的比例接種于改良增菌液中,置于4 ℃培養(yǎng)7 d后對(duì)小腸結(jié)腸炎耶爾森菌進(jìn)行分離鑒定[11]。分別使用O∶8血清型診斷血清和PCR擴(kuò)增foxA、ail基因的方法對(duì)細(xì)菌進(jìn)行鑒定,引物序列及退火溫度參考文獻(xiàn)[12]。
1.6攻毒后大鼠肝、脾、腸病理切片制作與組織病理變化觀(guān)察 攻毒后21d將全部實(shí)驗(yàn)大鼠的肝、脾、回腸末端組織完整取下,觀(guān)察其大體形態(tài)。之后,使用4%多聚甲醛固定48 h,石蠟包埋切片,蘇木精-伊紅(HE)染色后,置于光鏡下觀(guān)察組織的病理變化。
1.7統(tǒng)計(jì)學(xué)分析 應(yīng)用SPSS 21.0軟件進(jìn)行統(tǒng)計(jì)分析,差異比較采用單因素方差分析,以P<0.05為差異有統(tǒng)計(jì)學(xué)意義。
2 結(jié) 果
2.1OmpA蛋白表達(dá)與純化 將Ye92010-ompA重組克隆子表達(dá)后,采用His標(biāo)簽純化柱獲取目的蛋白OmpA,通過(guò)SDS-PAGE電泳對(duì)所獲取的蛋白進(jìn)行鑒定,結(jié)果顯示純化后的蛋白約38 kDa(圖1)。與預(yù)期結(jié)果相符[10],BCA法測(cè)定目的蛋白濃度為2.01 mg/mL。
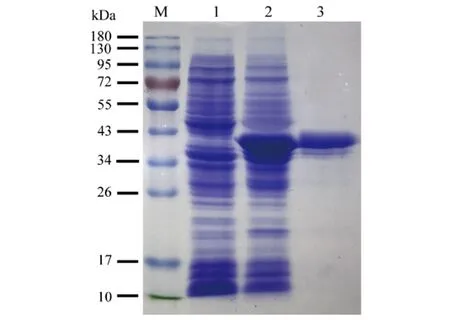
M:Marker;泳道1:未誘導(dǎo)的Ye92010-ompA重組克隆子全菌蛋白;泳道2:誘導(dǎo)后Ye92010-ompA重組克隆子全菌蛋白;泳道3:純化的Ye92010-OmpA圖1 小腸結(jié)腸炎耶爾森菌OmpA表達(dá)純化SDS-PAGE電泳圖Fig.1 SDS-PAGE electrophoretogram of purified Ye92010-OmpA
2.2大鼠攻毒試驗(yàn) 使用Ye92010攻毒4 h后,OmpA免疫組與對(duì)照組大鼠均表現(xiàn)出厭食、被毛雜亂、糞便呈松軟非顆粒狀等癥狀,至第2 d有部分恢復(fù)、第4 d全部恢復(fù)正常。連續(xù)觀(guān)察3周,OmpA免疫組和對(duì)照組大鼠均未出現(xiàn)死亡,各組大鼠肛溫?zé)o統(tǒng)計(jì)學(xué)差異(P>0.05)。
2.3糞便排菌檢測(cè) 使用Ye92010攻毒后,每日采集大鼠新鮮糞便,通過(guò)糞便分離培養(yǎng)和PCR對(duì)大鼠糞便排菌情況進(jìn)行檢測(cè)。結(jié)果顯示兩組大鼠糞便排菌起始日均為攻毒后第3 d,其中OmpA免疫組中,經(jīng)低劑量和高劑量攻毒的大鼠糞便排菌時(shí)間均為14 d,而對(duì)照組中經(jīng)低劑量和高劑量攻毒的大鼠糞便排菌時(shí)間分別為17 d和21 d。相對(duì)于對(duì)照組,OmpA免疫組糞便排菌時(shí)間明顯縮短,說(shuō)明OmpA蛋白免疫原性良好。
2.4攻毒后大鼠肝、脾、腸大體形態(tài)觀(guān)察 將攻毒3周后的實(shí)驗(yàn)大鼠進(jìn)行解剖并觀(guān)察各臟器的病理變化。由于Ye92010對(duì)大鼠具有較強(qiáng)的致病性,對(duì)照組大鼠出現(xiàn)了嚴(yán)重的病理改變,可觀(guān)察到明顯的組織變性及壞死灶,雙側(cè)腹股溝淋巴結(jié)腫大、肝臟黏連,甚至出現(xiàn)了腸膿腫和重度腸系膜液化等現(xiàn)象(圖2b1-b4)。相比較于對(duì)照組,OmpA免疫組大鼠癥狀較輕,僅出現(xiàn)輕度的皮下腫塊或脾腫大現(xiàn)象,腸管間界限模糊,腹股溝淋巴結(jié)腫大也僅局限于單側(cè)(圖2a1-a4)。空白對(duì)照組由于未經(jīng)攻毒而無(wú)病理學(xué)改變(圖2c1-c4)。
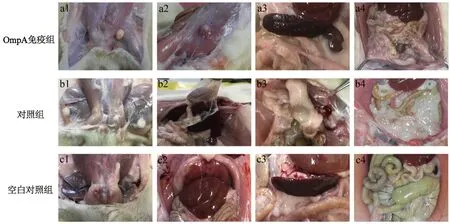
注:OmpA免疫組:經(jīng)OmpA免疫后,用Ye92010攻毒的大鼠大體形態(tài);對(duì)照組:等體積PBS代替OmpA免疫后,用Ye92010攻毒的大鼠大體形態(tài);空白對(duì)照組:未經(jīng)任何處理,正常飼養(yǎng)的大鼠大體形態(tài)a1:?jiǎn)蝹?cè)腹股溝淋巴結(jié)腫大;a2:皮下腫塊;a3:輕度脾水腫 a4:腸管間界限模糊;b1:雙側(cè)腹股溝淋巴結(jié)腫大;b2:肝臟黏連;b3:腸膿腫;b4:重度腸系膜液化;c1-c4:正常大鼠大體形態(tài)圖2 各組SD大鼠大體形態(tài)觀(guān)察Fig.2 Observation on general morphology of SD rats in each group
2.5細(xì)菌攻毒后大鼠肝組織病理學(xué)分析 在攻毒試驗(yàn)中,本研究同時(shí)進(jìn)行高、低菌量的攻毒方案。低劑量攻毒組中,OmpA免疫組大鼠肝組織局部可見(jiàn)炎性細(xì)胞(淋巴細(xì)胞,嗜酸性粒細(xì)胞)灶性浸潤(rùn),肝組織輕度水腫,細(xì)胞胞漿染色變淺(圖3B);對(duì)照組大鼠肝組織局部可見(jiàn)炎性細(xì)胞(淋巴細(xì)胞,嗜酸性粒細(xì)胞)灶性浸潤(rùn),肝組織水腫,細(xì)胞胞漿染色變淺,肝竇間隙水腫增寬(圖3D)。高劑量攻毒組中,OmpA免疫組大鼠肝組織局部可見(jiàn)炎性細(xì)胞(淋巴細(xì)胞為主)灶性浸潤(rùn),肝組織其他結(jié)構(gòu)均正常(圖3C);對(duì)照組大鼠肝組織局部可見(jiàn)炎性細(xì)胞(淋巴細(xì)胞,嗜酸性粒細(xì)胞)灶性浸潤(rùn),肝組織水腫,肝竇間隙水腫增寬(圖3E)。
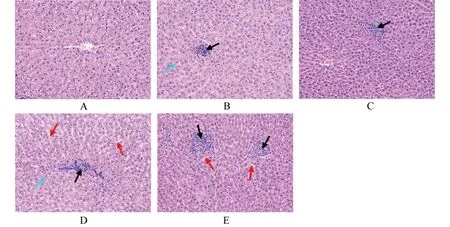
A:正常肝組織;B:低劑量攻毒OmpA免疫組大鼠肝組織;C:高劑量攻毒OmpA免疫組大鼠肝組織;D:低劑量攻毒對(duì)照組大鼠肝組織;E:高劑量攻毒對(duì)照組大鼠肝組織黑色箭頭所示為炎性細(xì)胞灶性浸潤(rùn),藍(lán)色箭頭所示為細(xì)胞胞漿染色變淺,紅色箭頭所示為肝竇間隙水腫增寬圖3 小腸結(jié)腸炎耶爾森菌Ye92010攻毒后各組SD大鼠肝組織病理學(xué)改變(×200)Fig.3 Histopathologic change of liver of SD rats infected with Ye92010 in each group(×200)
2.6細(xì)菌攻毒后大鼠脾組織病理學(xué)分析 低劑量攻毒組中,OmpA免疫組大鼠脾臟外膜完整覆蓋,脾竇大小正常,脾小結(jié)局部區(qū)域偶見(jiàn)濾泡中心變性壞死(圖4B);對(duì)照組大鼠脾臟外膜完整覆蓋,紅髓區(qū)域可見(jiàn)較多中性粒細(xì)胞散在浸潤(rùn),脾竇結(jié)構(gòu)疏松伴輕度擴(kuò)張和紅髓腫脹,濾泡中心變性壞死(圖4D)。高劑量攻毒組中,OmpA免疫組大鼠脾臟外膜完整覆蓋,紅髓區(qū)域可見(jiàn)組織結(jié)構(gòu)疏松、水腫,伴少量中性粒細(xì)胞浸潤(rùn)(圖4C);對(duì)照組大鼠脾臟外膜完整覆蓋,局部區(qū)域脾臟結(jié)構(gòu)紊亂,脾竇結(jié)構(gòu)疏松伴輕度擴(kuò)張,紅髓腫脹且局部區(qū)域可見(jiàn)少量中性粒細(xì)胞浸潤(rùn)(圖4E)。
2.7細(xì)菌攻毒后大鼠回腸組織病理學(xué)分析 低劑量攻毒組中,OmpA免疫組大鼠回腸組織粘膜層可見(jiàn)較多炎癥細(xì)胞(中性粒細(xì)胞,單核細(xì)胞,淋巴細(xì)胞)散在浸潤(rùn),伴有粘膜層及粘膜下層水腫(圖5B)。對(duì)照組大鼠回腸組織粘膜層可見(jiàn)大量炎癥細(xì)胞(中性粒細(xì)胞,單核細(xì)胞,淋巴細(xì)胞)廣泛浸潤(rùn),粘膜下層可見(jiàn)淋巴結(jié)結(jié)構(gòu),組織整體水腫(圖5D)。高劑量攻毒組中,OmpA免疫組大鼠回腸組織粘膜層局部可見(jiàn)炎癥細(xì)胞(淋巴細(xì)胞)散在浸潤(rùn),伴有組織局部區(qū)域壞死,粘膜下層及肌層結(jié)構(gòu)正常(圖5C)。對(duì)照組大鼠回腸組織粘膜層可見(jiàn)大量炎癥細(xì)胞(嗜酸性粒細(xì)胞,淋巴細(xì)胞)廣泛浸潤(rùn),粘膜下層組織水腫(圖5E)。
3 討 論
小腸結(jié)腸炎耶爾森菌屬于腸桿菌科,是一種革蘭陰性無(wú)芽孢桿菌。在歐洲,小腸結(jié)腸炎耶爾森菌是僅次于空腸彎曲菌和沙門(mén)氏菌的第三大人獸共患病病原菌;在我國(guó),已知的小腸結(jié)腸炎耶爾森菌暴發(fā)感染共2次,總計(jì)500多人[13-14]。但由于對(duì)小腸結(jié)腸炎耶爾森菌的監(jiān)測(cè)起步較晚,很多由該菌感染造成的疾病沒(méi)有得到很好的鑒定,故該菌的發(fā)病率在我國(guó)一直被嚴(yán)重低估,研制對(duì)抗致病性耶爾森菌的疫苗對(duì)于腸道疾病的預(yù)防和控制具有重大意義。近幾年,有大量研究表明OmpA是一種很好的免疫原性蛋白,可作為研制疫苗的候選蛋白。

A:正常脾組織;B:低劑量攻毒OmpA免疫組大鼠脾組織;C:高劑量攻毒OmpA免疫組大鼠脾組織;D:低劑量攻毒對(duì)照組大鼠脾組織;E:高劑量攻毒對(duì)照組大鼠脾組織黑色箭頭所示為濾泡中心變性壞死,紅色箭頭所示為中性粒細(xì)胞浸潤(rùn),藍(lán)色箭頭所示為紅髓腫脹,黃色箭頭所示為脾臟結(jié)構(gòu)紊亂圖4 小腸結(jié)腸炎耶爾森菌Ye92010攻毒后各組SD大鼠脾組織病理學(xué)改變(×200)Fig.4 Histopathologic change of spleen of SD rats infected with Ye92010 in each group(×200)

A:正常回腸組織;B:低劑量攻毒OmpA免疫組大鼠回腸組織;C:高劑量攻毒OmpA免疫組大鼠回腸組織; D:低劑量攻毒對(duì)照組大鼠回腸組織;E:高劑量攻毒對(duì)照組大鼠回腸組織紅色箭頭所示為粘膜層炎癥及水腫,藍(lán)色箭頭所示為粘膜下層炎癥及水腫,黃色箭頭所示為淋巴結(jié)結(jié)構(gòu)圖5 小腸結(jié)腸炎耶爾森菌Ye92010攻毒后各組SD大鼠回腸組織病理學(xué)改變(×200)Fig.5 Histopathologic change of ileum of SD rats infected with Ye92010 in each group(×200)
OmpA在革蘭陰性菌中廣泛存在,是最主要的外膜蛋白之一。有研究表明該蛋白主要通過(guò)粘附于粘膜表面、血清抵抗、侵襲、抗菌肽抵抗和使宿主細(xì)胞活化使細(xì)菌發(fā)揮致病作用,其高拷貝數(shù)和暴露于細(xì)菌表面的特性使其能促發(fā)固有免疫和適應(yīng)性免疫[15-16]。此外,OmpA可被中性粒細(xì)胞彈性蛋白酶特異性識(shí)別,并且可誘導(dǎo)實(shí)驗(yàn)動(dòng)物產(chǎn)生較高水平的抗體,但其對(duì)被免疫動(dòng)物的免疫保護(hù)能力目前尚不清楚[17-18]。
在本課題組前期工作中已經(jīng)通過(guò)體外實(shí)驗(yàn),將三種序列性ompA克隆表達(dá)蛋白純化后分別免疫小鼠所得的血清與三個(gè)型的OmpA進(jìn)行免疫雜交,結(jié)果證實(shí)OmpA良好的交叉免疫原性。因此本研究的目的在于證實(shí)OmpA的這種免疫原性是否能給機(jī)體帶來(lái)免疫保護(hù)。本研究通過(guò)腹股溝注射的方式對(duì)大鼠進(jìn)行OmpA免疫,攻毒后通過(guò)糞便分離培養(yǎng)和PCR鑒定菌株。結(jié)果顯示經(jīng)高劑量攻毒的對(duì)照組排菌時(shí)間最長(zhǎng),長(zhǎng)達(dá)21 d,且長(zhǎng)于低劑量攻毒對(duì)照組。而經(jīng)低劑量和高劑量攻毒的OmpA免疫組排菌時(shí)間相同,都在攻毒后14 d檢測(cè)不到排菌。排菌實(shí)驗(yàn)結(jié)果表明大鼠經(jīng)OmpA免疫后,Ye92010在腸道的定植受到阻礙并被快速地從腸道清除。
小腸結(jié)腸炎耶爾森菌具有嗜淋巴組織特征,感染小鼠后,在腸粘膜通過(guò)M細(xì)胞進(jìn)入下層的淋巴組織,即Peyer結(jié),這種侵襲可致大量白細(xì)胞增生,并與細(xì)胞外小腸結(jié)腸炎耶爾森菌形成微小膿腫,還可通過(guò)血流或淋巴管播散到肝、脾等臟器,引發(fā)腸以外的其他臟器感染[19-20]。本研究需收集大鼠糞便進(jìn)行排菌檢測(cè),故并未在急性感染期對(duì)大鼠進(jìn)行組織病理分析,而在其停止排菌后對(duì)所有大鼠進(jìn)行解剖,摘取肝、脾、腸,觀(guān)察組織病理學(xué)改變,研究致病性小腸結(jié)腸炎耶爾森菌OmpA對(duì)大鼠的免疫保護(hù)作用。大體觀(guān)察結(jié)果顯示OmpA免疫組出現(xiàn)腹股溝淋巴結(jié)腫大、皮下腫塊、脾結(jié)節(jié)、輕度腸系膜液化,對(duì)照組出現(xiàn)腹股溝淋巴結(jié)腫大、脾結(jié)節(jié)、腸膿腫、脾膿腫、重度腸系膜液化,其中OmpA免疫組中,低劑量攻毒組和高劑量攻毒組各有一只大鼠未觀(guān)察到明顯的組織病變及壞死,我們認(rèn)為可能是由于大鼠個(gè)體差異。組織病理切片結(jié)果顯示OmpA免疫組和對(duì)照組所有大鼠均出現(xiàn)不同程度的組織病變,肝病變出現(xiàn)炎癥細(xì)胞浸潤(rùn)和輕度水腫,脾病變出現(xiàn)濾泡中心變性壞死、水腫、炎癥細(xì)胞浸潤(rùn),腸病變出現(xiàn)粘膜層和粘膜下層炎癥細(xì)胞浸潤(rùn)和水腫。與大體觀(guān)察結(jié)果中出現(xiàn)的臟器膿腫、液化較為一致,且均表現(xiàn)為OmpA免疫組的病變程度輕于對(duì)照組,說(shuō)明OmpA對(duì)大鼠產(chǎn)生免疫保護(hù)。
綜上所述,致病性小腸結(jié)腸炎耶爾森菌OmpA免疫大鼠后,可使大鼠抵抗致病性小腸結(jié)腸炎耶爾森菌的能力大大增強(qiáng),排菌時(shí)間變短,組織病變變輕,OmpA對(duì)大鼠的免疫保護(hù)作用顯著。本研究表明OmpA不僅在體外水平表現(xiàn)出良好的免疫原性,更能在體內(nèi)水平給機(jī)體帶來(lái)免疫保護(hù),結(jié)合OmpA在3種致病性耶爾森菌中高度保守的特點(diǎn),推測(cè)OmpA可成為包括鼠疫在內(nèi)的致病性耶爾森菌疫苗的優(yōu)質(zhì)候選蛋白。
[1] Bottone EJ.Yersiniaenterocolitica: overview and epidemiologic correlates[J]. Microbes Infect, 1999, 1: 323-333.
[2] Koebnik R. Structural and functional roles of the surface-exposed loops of the beta-barrel membrane protein OmpA fromEscherichiacoli[J]. J Bacteriol, 1999, 181:3688-3694.
[3] Smajs D, Pilsl H, Braun V. Colicin U, a novel colicin produced byShigellaboydii[J]. J Bacteriol, 1997, 179:4919-4928.
[4] Erova TE, Rosenzweig JA, Sha J, et al. Evaluation of protective potential ofYersiniapestisouter membrane protein antigens as possible candidates for a new-generation recombinant plague vaccine[J]. Clin Vaccine Immunol, 2013, 20:227-238. DOI:10.1128/CVI.00597-12
[5] Singh SP, Williams YU, Miller S, et al. The C-terminal domain ofSalmonellaentericaserovar typhimurium OmpA is an immunodominant antigen in mice but appears to be only partially exposed on the bacterial cell surface[J]. Infect Immun, 2003, 71:3937-3946.
[6] Bhowmick R, Pore D, Chakrabarti MK. Outer membrane protein A (OmpA) ofShigellaflexneri2a induces TLR2-mediated activation of B cells: involvement of protein tyrosine kinase, ERK and NF-kappaB[J]. PLoS One, 2014, 9(10):e109107. DOI:10.1371/journal.pone.0109107
[7] Ayalew S, Shrestha B, Montelongo M, et al. Immunogenicity ofMannheimiahaemolyticarecombinant outer membrane proteins serotype 1-specific antigen, OmpA, OmpP2, and OmpD15[J]. Clin Vaccine Immunol, 2011, 18:2067-2074. DOI:10.1128/CVI.05332-11
[8] Gu W, Wang X, Qiu H, et al. Comparative antigenic proteins and proteomics of pathogenicYersiniaenterocoliticabio-serotypes 1B/O: 8 and 2/O: 9 cultured at 25 degrees C and 37 degrees C[J]. Microbiol Immunol, 2012, 56:583-594. DOI:10.1111/j.1348-0421.2012.00478.x
[9] Li K, Gu W, Liang J, et al. Gene polymorphism analysis ofYersiniaenterocoliticaouter membrane protein A and putative outer membrane protein A family protein[J]. BMC Genomics, 2014, 15:201. DOI:10.1186/1471-2164-15-201
[10] Chen Y, Duan R, Li X, et al. Homology analysis and cross-immunogenicity of OmpA from pathogenicYersiniaenterocolitica,YersiniapseudotuberculosisandYersiniapestis[J]. Mol Immunol, 2015, 68:290-299. DOI:10.1016/j.molimm.2015.09.016
[11] Wang X, Cui Z, Jin D, et al. Distribution of pathogenicYersiniaenterocoliticain China[J]. Eur J Clin Microbiol Infect Dis, 2009, 28:1237-1244. DOI:10.1007/s10096-009-0773-x
[12] Liang J, Wang X, Xiao Y, et al. Prevalence ofYersiniaenterocoliticain pigs slaughtered in Chinese abattoirs[J]. Appl Environ Microbiol, 2012, 78:2949-2956. DOI:10.1128/AEM.07893-11
[13] Backhans A, Fellstrom C, Lambertz ST. Occurrence of pathogenicYersiniaenterocoliticaandYersiniapseudotuberculosisin small wild rodents[J]. Epidemiol Infect, 2011, 139:1230-1238. DOI:10.1017/S0950268810002463
[14] Wang X, Qiu H, Jin D, et al. O:8 serotypeYersiniaenterocoliticastrains in China[J]. Int J Food Microbiol, 2008, 125:259-266. DOI:10.1016/j.ijfoodmicro.2008.04.016
[15] Smith SG, Mahon V, Lambert MA, et al. A molecular Swiss army knife: OmpA structure, function and expression[J]. FEMS Microbiol Lett, 2007, 273:1-11. DOI:10.1111/j.1574-6968.2007.00778.x
[16] Confer AW, Ayalew S. The OmpA family of proteins: roles in bacterial pathogenesis and immunity[J]. Vet Microbiol, 2013, 163:207-222.DOI:10.1016/j.vetmic.2012.08.019
[17] Lower BH, Yongsunthon R, Vellano FP 3rd, et al. Simultaneous force and fluorescence measurements of a protein that forms a bond between a living bacterium and a solid surface[J]. J Bacteriol, 2005, 187:2127-2137. DOI:10.1128/JB.187.6.2127-2137.2005
[18] Gatto NT, Dabo SM, Hancock RE, et al. Characterization of, and immune responses of mice to, the purified OmpA-equivalent outer membrane protein ofPasteurellamultocidaserotype A:3 (Omp28)[J]. Vet Microbiol, 2002, 87:221-235.
[19] Autenrieth IB, Firsching R. Penetration of M cells and destruction of Peyer’s patches byYersiniaenterocolitica: an ultrastructural and histological study[J]. J Med Microbiol, 1996, 44:285-294. DOI:10.1099/00222615-44-4-285
[20] Bottone EJ.Yersiniaenterocolitica: the charisma continues[J]. Clin Microbiol Rev, 1997, 10:257-276
猜你喜歡
全科護(hù)理(2022年10期)2022-12-26 21:19:15
中國(guó)合理用藥探索(2022年1期)2022-11-26 00:22:32
今日農(nóng)業(yè)(2022年4期)2022-11-16 19:42:02
鄉(xiāng)村科技(2021年33期)2021-03-16 02:26:54
國(guó)際放射醫(yī)學(xué)核醫(yī)學(xué)雜志(2021年10期)2021-02-28 08:41:58
藥學(xué)與臨床研究(2015年4期)2015-06-05 11:35:54
衛(wèi)生職業(yè)教育(2014年24期)2014-05-20 09:05:38
同位素(2014年2期)2014-04-16 04:57:20
中國(guó)合理用藥探索(2014年11期)2014-03-11 20:30:20
