落羽杉根系有機酸與NSC代謝對三峽消落帶水位變化的響應
2018-06-22 11:29:42陳紅純馬文超袁中勛
生態學報 2018年9期
王 婷,魏 虹,周 翠,陳紅純,李 瑞,馬文超,袁中勛
西南大學生命科學學院, 三峽庫區生態環境教育部重點實驗室,重慶市三峽庫區植物生態與資源重點實驗室,重慶 400715
三峽工程修建后,庫區“冬蓄夏排”的管理方式導致水位在海拔145—175 m間變動,庫區形成了垂直落差最高達30 m的消落帶[1]。消落帶水陸環境交替變化,大多數原有植物因不能適應周期性變化的水淹而消亡,導致植被退化、水土流失等環境問題,生態服務功能降低。為解決上述問題,在前期研究中,通過室內模擬試驗篩選出一批耐水淹植物,但將其應用于消落帶植被重建后發現,僅有部分植物能在消落帶生存,為消落帶植被恢復帶來新挑戰[2-3]。因此,在消落帶原位開展適生植物的水淹耐受機制研究極為重要,研究結果將能為庫區植被重建與管理提供有效的理論指導。
落羽杉(Taxodiumdistichum),杉科落羽杉屬落葉喬木,是三峽庫區消落帶植被重建的優良植物[4],有研究表明不同樹齡落羽杉對水淹的響應差異不明顯[5]。近年來,國內外學者就落羽杉在水分、鹽分脅迫下的生長特征[6]、光合生理[7-8]、營養元素變化與分配[3,9]、次生代謝[10]等展開了研究,但對落羽杉在水淹逆境下的物質代謝關注較少。在三峽消落帶反季節、高強度的水淹脅迫下,植物根系代謝途徑與其水淹耐受性密切相關[11-12]。根系初生代謝產物非結構性碳水化合物(non-structural carbohydrates,NSC)是植物在水淹生境中的能量來源,能保證植物生命活動所需的能量供應[13-14]。植物次生代謝在逆境防御中扮演著重要角色[15-16],其中根系次生代謝產物有機酸能提高水淹逆境下植物體內多種酶的活性,減輕對植物的毒害作用,增強植株耐水淹的能力[17]。
目前已有關于有機酸、NSC分別對水淹逆境的響應研究,主要集中于室內模擬試驗下的相應含量變化方面[12,17]。植物有機酸代謝是由其初生代謝衍生而來[16],與NSC代謝密切相關,但目前關于根系有機酸和NSC代謝對三峽消落帶大尺度水位變化的響應研究尚缺乏,而二者對消落帶原位環境的響應對策在其植被重建與管理工作中至關重要。因此,本研究基于三峽庫區消落帶原位環境,以適生木本植物落羽杉為對象,探究庫區水文變化對落羽杉根系有機酸和NSC的影響,明確落羽杉根系有機酸和NSC代謝響應對策,以期為消落帶生態恢復提供理論依據。
1 材料與方法
1.1 試驗材料與設計
2015年9月選取長勢基本一致的25株落羽杉兩年生盆栽苗(盆高21 cm,盆口直徑27 cm),每盆1株,盆中裝入三峽庫區消落帶紫色土壤12.5 kg。于2015-09- 15采集初始樣(5個重復),將剩余20株試驗用苗隨機分為3組(2∶1∶1),分別置于重慶市忠縣汝溪河消落帶(30°25′55.47″N,108°09′59.18″E)的175 m(對照,SS)、170 m(中度水淹處理組,MS)、165 m(深度水淹處理組,DS)3個不同海拔,處理強度如表1所示。試驗區屬亞熱帶東南季風氣候。全年≥10℃年積溫5787℃,年均溫18.2℃,無霜期341 d,日照時數1327.5 h,日照率29%,太陽總輻射能83.7×4.18 kJ/cm2,年降雨量1200 mm,相對濕度80%[3]。
根據三峽庫區消落帶水位變動規律,于2016-02- 17(海拔170 m退水)采集海拔175、170 m的植物樣品(第一批樣品),2016-04- 16(海拔165 m退水)采集海拔175、165 m的植物樣品(第二批樣品),并分別用測高桿、游標卡尺測量株高和基徑。每個處理5個重復,試驗處理從2015-09- 15蓄水期開始,至2016-04- 16結束。試驗期間落羽杉的存活率為100%。

表1 三峽庫區原位試驗處理水平
SS:對照control;MS:中度水淹moderate submergence;DS:深度水淹deep submergence
1.2 根系代謝產物的測定
將試驗苗主根、側根分開取樣,立即裝入冰盒帶回實驗室,依次用自來水和去離子水清洗干凈,置于110℃烘箱內殺青15 min,再調至80℃烘干至恒重;將烘干的樣品采用MM400球磨儀研磨成小于2 mm的粉末,封裝待測。
1.2.1 根系有機酸的測定
在預備試驗和前人研究基礎上[18-19],采用日本島津LC高效液相色譜儀,以95% 的20 mmol/L KH2PO4緩沖液(用磷酸調pH=2.5)與5% 的甲醇分別作為流動相的水相和有機相,在Sepax Sapphire C18色譜柱(4.6×250 mm,5 μm)上進行測定。檢測器為Agilent 1100二極管陣列多波長檢測器,流動相流速為0.9 mL/min;檢測波長為210 nm;柱溫30℃,進樣量20 μL。98%的蘋果酸、檸檬酸、酒石酸、草酸標準品由成都普菲德生物技術有限公司提供;97%的分析純莽草酸由梯希愛(上海)化成工業發展有限公司提供;85%的色譜純甲醇、85%的分析純磷酸、99.5%的色譜純磷酸二氫鉀由成都市科龍化工試劑廠提供。
溶液制備時,準確稱取落羽杉粉末0.1 g置于10 mL離心管中,加入5 mL超純水后超聲提取1 h,冷卻至室溫,在8000 r/min下離心10 min,取上清液后用孔徑為0.45 μm的注射式過濾器(美國Millpore)過濾,濾液即為供試液[18,20]。
主根、側根有機酸分別以每克主根、側根干物質含有的有機酸毫克數計(mg/g, DW);總根有機酸為主根、側根組成的根系整體含有的有機酸平均含量(mg/g, DW)。
1.2.2 根系NSC的測定
本文中植物NSC=可溶性糖+淀粉。準確稱取落羽杉粉末0.01 g置于10 mL離心管中,加入5 mL 80%乙醇,80℃水浴加熱40 min,冷卻至室溫,7000 r/min離心12 min后取上清液。重復提取2次后合并上清液并定容至50 mL,所得液體即為可溶性糖待測液。在625 nm波長下采用紫外可見光分光光度計(UV- 2550,Japan)測定待測液中的可溶性糖含量[21]。將淀粉用酸水解為可溶性糖后用上述蒽酮比色法進行測定。
主根、側根NSC分別以每克主根、側根干物質含有的NSC毫克數計(mg/g, DW);總根NSC為主根、側根組成的根系整體含有的NSC平均含量(mg/g, DW)。
1.3 數據分析
本研究采用SPSS 22.0、Origin 8.5對數據進行統計分析和繪圖。用獨立樣本T檢驗分析不同水淹對落羽杉根系有機酸、NSC的影響;采用配對樣本T檢驗分析水淹前初始值與水淹后處理值之間的差異顯著性;用Pearson相關系數法評價落羽杉根系代謝產物間的相關性。
2 結果與分析
2.1 落羽杉生長的變化
三峽庫區消落帶原位水淹對落羽杉生長的影響如表2所示。由表可知,除第二批樣品SS組株高顯著高于初始值外,其余均無明顯差異;MS組和DS組株高均在SS組之下,且DS組顯著低于SS組(P<0.05),說明消落帶原位水淹抑制了落羽杉的高生長(表2)。與初始值相比,落羽杉基徑在消落帶原位水淹下雖表現為增長的趨勢,但均無顯著差異;基徑在不同水淹處理組與SS組間也無明顯差異。
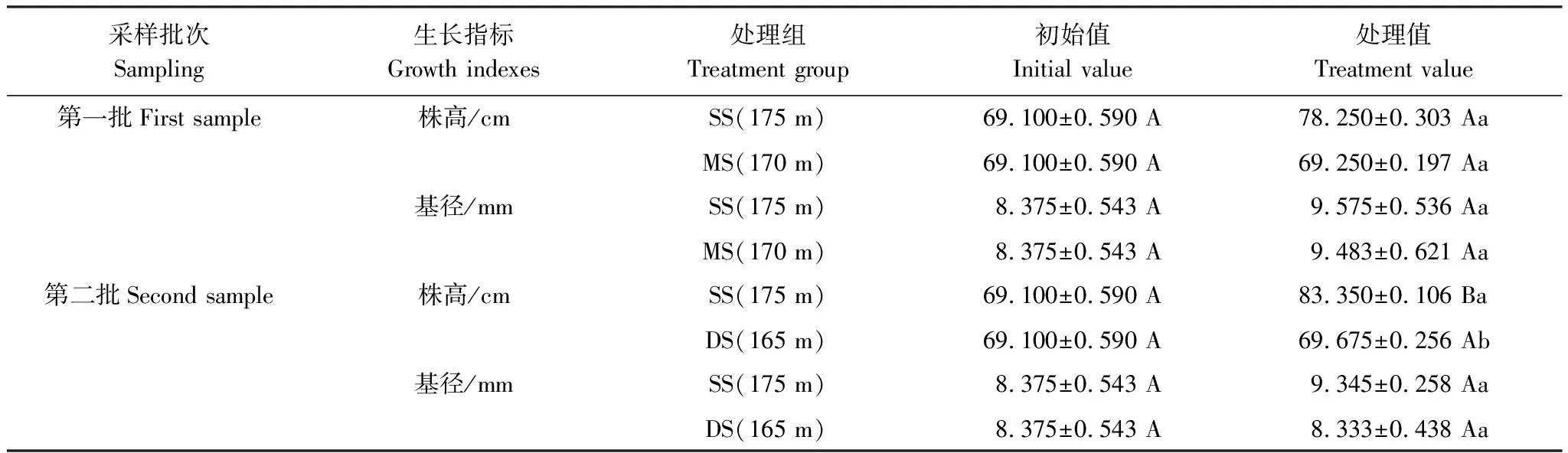
表2 三峽庫區水分變化下落羽杉的生長特征
表中數值為平均值±標準誤(n=5);同行不同大寫字母表示水淹前初始值與水淹后處理值之間有顯著差異性(P<0.05);同列不同小寫字母表示同批次處理之間有顯著差異(P<0.05)
2.2 根系有機酸含量的變化
不同水淹強度對落羽杉主根、側根及總根有機酸含量的影響不同(表3)。第一批樣品中,與SS組相比,MS組水淹導致主根草酸、酒石酸、莽草酸、蘋果酸、檸檬酸含量均增高,其中蘋果酸和檸檬酸達到顯著水平(P<0.05);與SS組相比,側根酒石酸、莽草酸、蘋果酸含量在MS組脅迫時呈不顯著上升趨勢,且均在同組主根含量之上,相反的是,側根草酸含量有所下降,檸檬酸含量顯著下降(P<0.05);總根有機酸含量與側根有機酸含量變化趨勢一致,MS組總根酒石酸、莽草酸、蘋果酸含量均高于SS組,MS組總根草酸、檸檬酸含量低于SS組,但均未達顯著水平(表3)。
由表3可知,第二批樣品中,與SS組相比,DS組主根酒石酸、莽草酸、蘋果酸、檸檬酸均表現為不顯著下降;DS組側根草酸、酒石酸、蘋果酸、檸檬酸含量也均低于SS組,且酒石酸和檸檬酸達到顯著水平(P<0.05);總根有機酸含量與側根有機酸含量變化一致,在消落帶原位水淹脅迫下,DS組總根有機酸含量均低于SS組,且DS組總根酒石酸和檸檬酸顯著低于SS組(表3,P<0.05)。
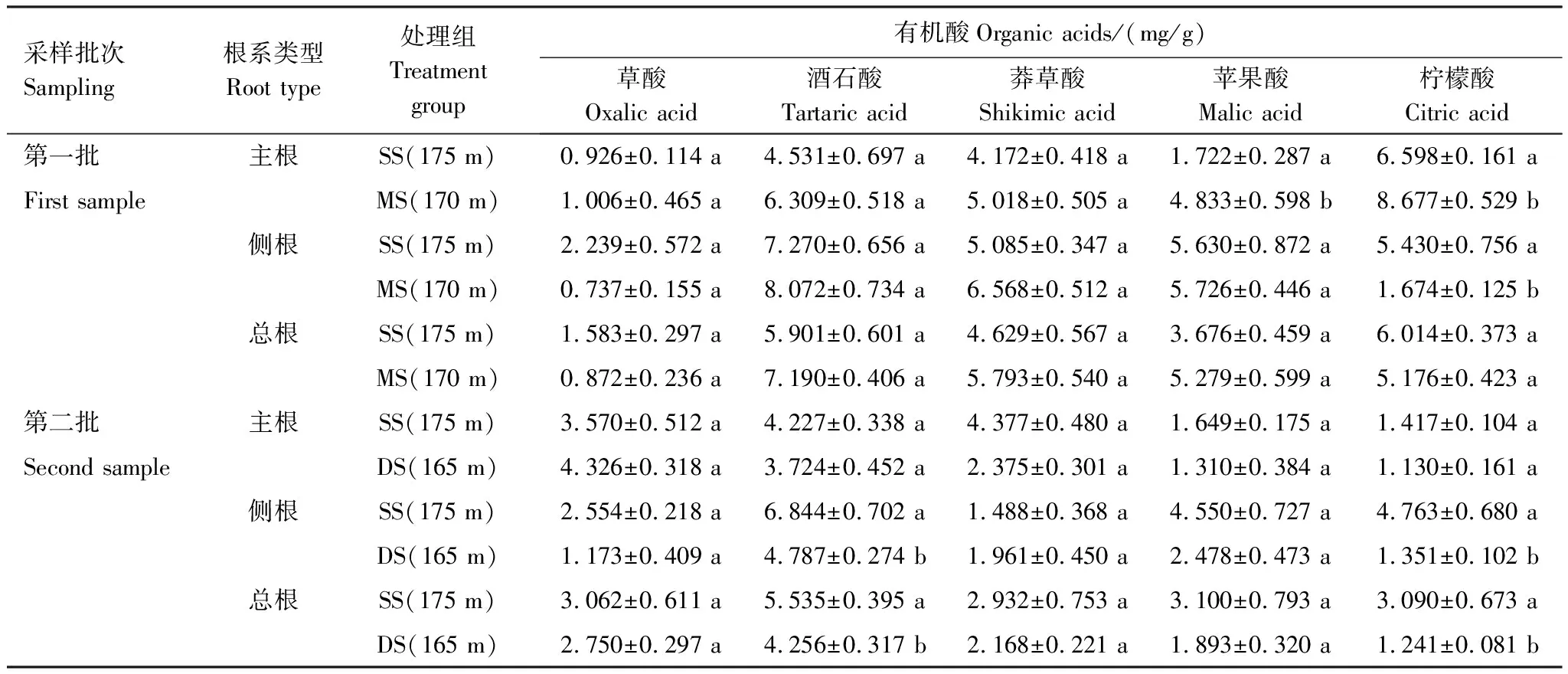
表3 三峽庫區水分變化下落羽杉根系的有機酸含量
表中數值為平均值±標準誤(n=5);同列不同小寫字母分別表示同批次同種根系類型處理之間有顯著差異(P<0.05)
2.3 根系NSC的變化
2.3.1 根系可溶性糖的變化
圖1是三峽庫區消落帶原位水淹對落羽杉根系可溶性糖的影響。由圖1可知,第一批樣品中除側根可溶性糖顯著低于初始值外,MS組落羽杉其余根系可溶性糖含量與SS組、初始值均無顯著變化。在第二批樣品中,DS組落羽杉主根、側根、總根可溶性糖含量均顯著低于SS組和初始值(圖1,P<0.05)。
2.3.2 根系淀粉的變化
庫區消落帶原位水淹對根系淀粉的影響如圖2所示。由圖2可知,第一批樣品中MS組主根、側根、總根淀粉含量均顯著高于SS組(P<0.01),總根淀粉含量與初始值無顯著差異;DS組主根、側根、總根淀粉含量與SS組無明顯差異,但均顯著低于初始值(圖2,P<0.05)。
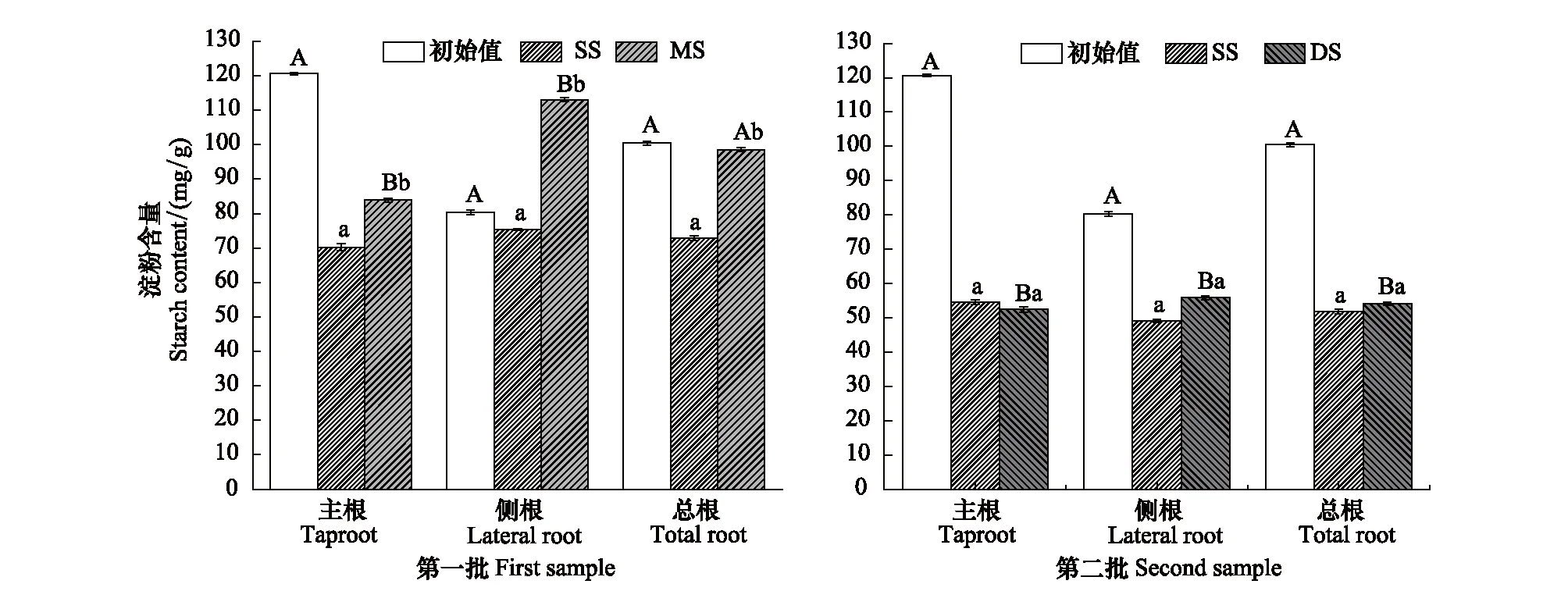
圖2 不同水淹條件下落羽杉根系淀粉含量Fig.2 Starch content of T. distichum in different treatments
2.3.3 根系NSC的變化
圖3是三峽庫區消落帶原位水淹對落羽杉根系NSC含量的影響。與淀粉變化一致,第一批樣品中MS組NSC含量均顯著高于SS組(P<0.01);MS組除主根NSC含量顯著低于初始值外,側根、總根NSC與初始值無明顯差異(圖3)。第二批樣品中NSC含量與可溶性糖變化類似,DS組NSC含量顯著低于SS組和初始值(P<0.01)。
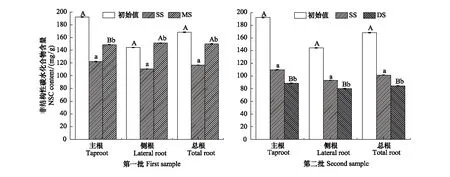
圖3 不同水淹條件下落羽杉根系NSC含量Fig.3 NSC content of T. distichum in different treatments
2.4 相關性分析
2.4.1 根系不同有機酸的相關性分析
落羽杉根系不同種類有機酸的相關性分析表明,草酸、酒石酸、莽草酸、蘋果酸、檸檬酸含量在主根與側根間均未表現出顯著相關性,但均分別與總根有機酸表現出顯著正相關關系(表4,P<0.05)。側根草酸與其他有機酸無顯著相關性,而主根、總根草酸分別與總根莽草酸、主根蘋果酸及主根、總根檸檬酸表現出顯著負相關關系(P<0.05)。落羽杉主根、側根、總根酒石酸與側根、總根莽草酸具有極顯著的正相關關系(P<0.01)。主根莽草酸與其他有機酸的相關性較弱,而側根、總根莽草酸與酒石酸、主根檸檬酸達到極顯著正相關水平(P<0.01)。與側根草酸含量變化趨勢類似,側根蘋果酸、檸檬酸與其他有機酸無顯著相關性,而主根、總根蘋果酸與主根檸檬酸表現為極顯著正相關關系(P<0.01)。
2.4.2 根系有機酸和NSC的相關性分析
相關性分析表明,落羽杉主根、側根和總根有機酸與NSC間呈現出不同的相關關系(表5)。結果顯示,相對淀粉、NSC而言,落羽杉根系可溶性糖與有機酸的相關性更弱,其中側根可溶性糖與根系有機酸均未表現出顯著相關性。側根草酸、酒石酸、檸檬酸及主根酒石酸、莽草酸與可溶性糖、淀粉、NSC間的相關性較弱,而主根、總根草酸、蘋果酸、檸檬酸及側根、總根莽草酸分別與淀粉、NSC含量相關性較強,均達到顯著相關水平(P<0.05)。主根、總根草酸與淀粉、NSC含量表現出顯著負相關關系;主根、總根蘋果酸、檸檬酸及側根、總根莽草酸與淀粉、NSC表現出顯著正相關關系;主根檸檬酸與淀粉、NSC含量的相關程度較高,相關系數均在0.9左右。
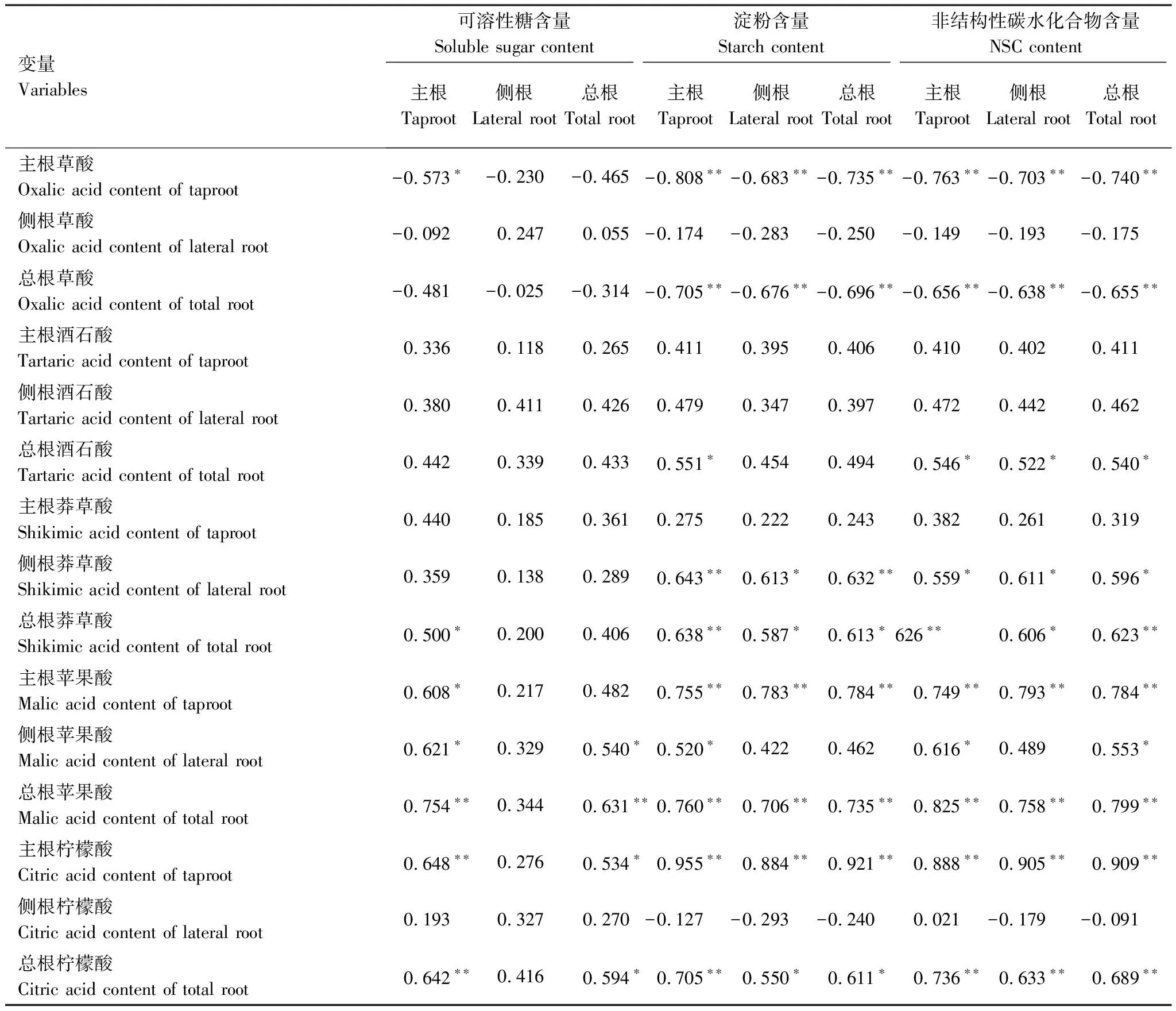
表5 落羽杉根系有機酸與NSC含量間的相關性分析
**表示在α = 0.01水平下達到極顯著相關性;*表示α = 0.05水平下達到顯著相關性
3 討論與結論
三峽水庫建成運行后,145—175 m海拔位間周期性漲落的水位導致庫岸植被退化,生物多樣性降低,三峽庫區生態問題突出。目前,消落帶的植被恢復研究備受關注與重視[2- 3,22]。落羽杉是杉科落羽杉屬喬木,具有屈膝狀的呼吸根,是消落帶植被恢復的優良候選物種[3,7]。本研究顯示,落羽杉在消落帶經過非生長季節水淹后,存活率為100%,僅有DS組明顯抑制了落羽杉的高生長(表2)。有文獻表明,植物在水淹下可加快形成通氣組織,有助于耐受氧氣缺乏的水淹逆境[23]。與模擬研究一致,落羽杉在長期完全淹沒下,基徑均無顯著變化[24],可能與莖基通氣組織等的形成有關,表明落羽杉株高、基徑能較積極地響應庫區水淹。
水淹是限制消落帶植物生長的主要因素之一。水淹下氣體擴散受阻,首先導致植物根際缺氧,植物根系代謝受到嚴重影響[25- 26]。在長期適應水淹過程中,植物根系有機酸代謝發揮著重要作用[27]。已有研究表明,植物根系分泌有機酸種類和含量的增加是抵御水分等逆境脅迫的重要途徑之一[17,28]。如蘋果酸可維持逆境下跨膜電荷的平衡,與草酸等可作為硝酸鹽還原過程的電荷補償[29- 30],提供能量來源。相關模擬三峽庫區水分變化的研究發現,落羽杉通過側根分泌大量蘋果酸和莽草酸,增強水淹耐受能力[10],本研究結果與之相似。與SS組相比,MS組落羽杉主根有機酸含量均增加;側根、總根酒石酸、莽草酸、蘋果酸含量均不顯著增加。水淹導致落羽杉根系缺氧,根系代謝受阻進一步破壞活性氧代謝平衡,誘發氧化脅迫[31],有機酸種類與含量的增加可能與其為落羽杉根系抗氧化系統提供質子,減少氧化傷害有關[32]。說明消落帶中度水淹脅迫下,落羽杉根系保持著較高的有機酸代謝活力。與上述有機酸含量變化不同,側根檸檬酸含量表現為顯著下降,且隨著水淹強度的增大,DS組落羽杉側根、總根檸檬酸顯著低于SS組(表3)。水淹逆境的實質是缺氧所誘導的次生脅迫,深度水淹導致植物面臨嚴重缺氧[33- 34],植物三羧酸循環嚴重受阻[26],由此可能導致檸檬酸含量顯著下降。已有研究發現,楓楊根系酒石酸在水淹脅迫下顯著增加,但隨著水淹時間的延長顯著下降[35]。與已有研究一致,MS組落羽杉根系酒石酸不顯著高于SS組,但在更長時間的深度水淹脅迫下,DS組根系酒石酸均低于SS組,且側根和總根達到顯著下降水平(表3),可能與長時間深度水淹脅迫加大對酒石酸的利用效率有關。除側根、總根酒石酸、檸檬酸顯著低于SS組,主根草酸和側根莽草酸不顯著高于SS組外,DS組其他有機酸均不顯著低于SS組,說明落羽杉根系通過維持與對照SS組相同水平的有機酸代謝,適應消落帶長期深度水淹脅迫。與模擬研究一致[10],經歷不同程度水淹脅迫的落羽杉側根、總根有機酸含量變化趨勢一致,表明在有機酸代謝中,側根一直發揮著主導作用(表3)。
有機酸作為次生代謝產物,是由初生代謝衍生,長期與環境適應的結果。相關分析發現,根系不同有機酸之間存在不同程度相關性,表明不同有機酸代謝聯系緊密(表4)。落羽杉主根、總根草酸、蘋果酸、檸檬酸及側根、總根莽草酸與淀粉、NSC間表現出顯著相關關系(表5),進一步證實根系有機酸與NSC代謝密切相關。缺氧脅迫下,糖酵解、硝酸鹽還原等厭氧代謝途徑是耐水淹植物維持生命活力的能源[36]。一方面,NSC是植物在水淹下的能量來源,以淀粉形式儲存,通過葡萄糖、果糖、蔗糖等可溶性糖方式被厭氧代謝等途徑利用[37];另一方面,草酸、蘋果酸等有機酸可平衡硝酸還原過程中的電荷[29- 30],加強硝酸鹽還原等代謝途徑,共同增強缺氧脅迫下的能量供應。相關研究表明,杉科植物可通過增加根系淀粉、可溶性糖含量增強耐受長期水淹的能力[12]。與前人研究一致,本試驗發現,第一批樣品中MS組落羽杉淀粉、NSC含量均顯著高于SS組,可溶性糖無明顯變化,總根NSC與水淹前初始值無明顯差異,說明落羽杉根系能維持較高的淀粉含量適應中度水淹脅迫。而與已有研究不同的是,第二批樣品中DS組落羽杉可溶性糖和NSC含量均顯著低于SS組和初始值(圖1,圖3)。可能因為在環境更嚴峻的深度水淹脅迫下,為保證落羽杉正常的生理功能,能耗的增加導致根系的NSC逐漸被消耗。雖然DS組淀粉含量顯著低于水淹前的初始值,但與同批次的SS組無明顯差異。表明落羽杉在消落帶長時間深度水淹脅迫下,根系可維持一定的淀粉含量增強適應能力。
綜上所述,在三峽庫區消落帶原位水淹脅迫下,根系有機酸和NSC代謝在維持落羽杉長時間深度水淹下的生理功能方面發揮著重要作用。研究結果表明,側根有機酸代謝作用優于主根作用,根系有機酸與淀粉、NSC間呈現出較強的相關性。落羽杉根系可通過維持一定的NSC含量,尤其是淀粉含量,保持與對照相同的有機酸代謝水平,保證長期深度水淹下生理功能所需的能量供應,較好地適應消落帶原位環境。
參考文獻(References):
[1] 羅芳麗, 王玲, 曾波, 葉小齊, 陳婷, 劉巔, 張艷紅, Kuhn A. 三峽庫區岸生植物野古草(ArundinellaanomalaSteud.)光合作用對水淹的響應. 生態學報, 2006, 26(11): 3602- 3609.
[2] 樊大勇, 熊高明, 張愛英, 劉曦, 謝宗強, 李兆佳. 三峽庫區水位調度對消落帶生態修復中物種篩選實踐的影響. 植物生態學報, 2015, 39(4): 416- 432.
[3] 馬文超, 劉媛, 周翠, 王婷, 魏虹. 水位變化對三峽庫區消落帶落羽杉營養特征的影響. 生態學報, 2017, 37(4): 1128- 1136.
[4] 李昌曉, 耿養會, 葉兵, Rebcca S. 落羽杉與池杉幼苗對多種脅迫環境的響應及其對三峽庫區庫岸防護林營建的啟示(英文). 林業科學, 2010, 46(10): 144- 152.
[5] Powell A S. Response of Baldcypress (Taxodiumdistichum) at different life stages to flooding and salinity[D].Greenville, North Carolina: East Carolina University, 2014.
[6] Li B, Du C L, Yuan X Z, Willison J H M, Xiao H Y. Suitability ofTaxodiumdistichumfor afforesting the littoral zone of the Three Gorges Reservoir. PloS One, 2016, 11(1): e0146664.
[7] Wang C Y, Li C X, Wei H, Xie Y Z, Han W J. Effects of long-term periodic submergence on photosynthesis and growth ofTaxodiumdistichumandTaxodiumascendenssaplings in the hydro-fluctuation zone of the Three Gorges Reservoir of China. PloS One, 2016, 11(9): e0162867.
[8] Lauer N, Ross C. Physiological and oxidative stress responses of baldcypress in response to elevated salinity: linking and identifying biomarkers of stress in a keystone species. Acta Physiologiae Plantarum, 2016, 38(12): 275-275.
[9] 汪貴斌, 曹福亮. 不同土壤水分含量下落羽杉根、莖、葉營養水平的差異. 林業科學研究, 2004, 17(2): 213- 219.
[10] 李昌曉, 鐘章成. 三峽庫區消落帶土壤水分變化對落羽杉(Taxodiumdistichum)幼苗根部次生代謝物質含量及根生物量的影響. 生態學報, 2007, 27(11): 4394- 4402.
[11] Jackson M B, Colmer T D. Response and adaptation by plants to flooding stress. Annals of Botany, 2005, 96(4): 501- 505.
[12] 張艷婷, 張建軍, 王建修, 吳曉洪, 陳寶強, 李鵬飛, 王志臻. 長期水淹對‘中山杉118’幼苗呼吸代謝的影響. 植物生態學報, 2016, 40(6): 585- 593.
[13] Qi B Y, Yang Y, Yin Y L, Xu M, Li H G. De novo sequencing, assembly, and analysis of theTaxodium′Zhongshansa′ roots and shoots transcriptome in response to short-term waterlogging. BMC Plant Biology, 2014, 14(1): 201-201.
[14] 施美芬, 曾波, 申建紅, 類淑桐, 朱智, 劉建輝. 植物水淹適應與碳水化合物的相關性. 植物生態學報, 2010, 34(7): 855- 866.
[15] Kutchan T M. Ecological arsenal and developmental dispatcher. The paradigm of secondary metabolism. Plant Physiology, 2001, 125(1): 58- 60.
[16] 王莉, 史玲玲, 張艷霞, 劉玉軍. 植物次生代謝物途徑及其研究進展. 武漢植物學研究, 2007, 25(5): 500- 508.
[17] 黃文斌, 馬瑞, 楊迪, 劉永, 宋金鳳. 土壤逆境下植物根系分泌的有機酸及其對植物生態適應性的影響. 安徽農業科學, 2013, 41(34): 13316- 13319.
[18] 高智席, 周光明, 黃成, 李昌曉, 王莉. 離子抑制-反相高效液相快速測定池杉、落羽杉根系中有機酸. 藥物分析雜志, 2005, 25(9): 1082- 1085.
[19] 黃天志, 王世杰, 劉秀明, 劉虹, 吳沿友, 羅緒強. 逐級提取-高效液相色譜法快速測定植物組織中8種有機酸. 色譜, 2014, 32(12): 1356- 1361.
[20] 葉思誠, 譚曉風, 袁軍, 張雪潔, 石斌. 油茶根系及分泌物中有機酸的HPLC法測定. 南京林業大學學報: 自然科學版, 2013, 37(6): 59- 63.
[21] Yemm E W, Willis A J. The estimation of carbohydrates in plant extracts by anthrone. Biochemical Journal, 1954, 57(3): 508- 514.
[22] 任慶水, 馬朋, 李昌曉, 楊予靜, 馬駿. 三峽庫區消落帶落羽杉(Taxodiumdistichum)與柳樹(Salixmatsudana)人工植被對土壤營養元素含量的影響. 生態學報, 2016, 36(20): 6431- 6444.
[23] 張小萍, 曾波, 陳婷, 葉小齊, 羅芳麗, 劉巔. 三峽庫區河岸植物野古草(Arundinellaanomalavar.depauperataKeng)莖通氣組織發生對水淹的響應. 生態學報, 2008, 28(4): 1864- 1871.
[24] Iwanaga F, Tanaka K, Nakazato I, Yamamoto F. Effects of submergence on growth and survival of saplings of three wetland trees differing in adaptive mechanisms for flood tolerance. Forest Systems, 2015, 24(1): e001.
[25] Gibbs J, Greenway H. Review: mechanisms of anoxia tolerance in plants. I. Growth, survival and anaerobic catabolism. Functional Plant Biology, 2003, 30(3): 1- 47.
[26] Fukao T, Bailey-Serres J. Plant responses to hypoxia—is survival a balancing act? Trends in Plant Science, 2004, 9(9): 449- 456.
[27] 李昌曉, 鐘章成, 陶建平. 不同水分條件下池杉幼苗根系的蘋果酸、莽草酸含量及生物量. 林業科學, 2008, 44(10): 1- 7.
[28] 楊迪, 馬瑞, 黃文斌, 劉永, 宋金鳳. 養分和水分脅迫下2年生落葉松根系有機酸的分泌行為研究. 安徽農業科學, 2013, 41(36): 13932- 13934, 14023- 14023.
[29] Henry A, Doucette W, Norton J, Bugbee B. Changes in crested wheatgrass root exudation caused by flood, drought, and nutrient stress. Journal of Environmental Quality, 2007, 36(3): 904- 912.
[30] Marschner H. Mineral Nutrition of Higher Plants. 2nd ed. London, UK: Academic Press, 1995: 1250- 1250.
[31] Blokhina O, Virolainen E, Fagerstedt K V. Antioxidants, oxidative damage and oxygen deprivation stress: a review. Annals of Botany, 2003, 91(2): 179- 194.
[32] 陳暄, 周家樂, 唐曉清, 王康才. 水分脅迫條件下不同栽培居群菘藍中4種有機酸的變化. 中國中藥雜志, 2009, 34(24): 3195- 3198.
[33] Colmer T D. Long-distance transport of gases in plants: a perspective on internal aeration and radial oxygen loss from roots. Plant, Cell & Environment, 2003, 26(1): 17- 36.
[34] 趙可夫. 植物對水澇脅迫的適應. 生物學通報, 2003, 38(12): 11- 14.
[35] 李昌曉, 魏虹, 呂茜, 張曄. 水分脅迫對楓楊幼苗生長及根系草酸與酒石酸含量的影響. 林業科學, 2010, 46(11): 81- 88.
[36] 譚淑端, 朱明勇, 張克榮, 黨海山, 張全發. 植物對水淹脅迫的響應與適應. 生態學雜志, 2009, 28(9): 1871- 1877.
[37] 譚淑端, 朱明勇, 黨海山, 王勇, 張全發. 三峽庫區狗牙根對深淹脅迫的生理響應. 生態學報, 2009, 29(7): 3685- 3691.
