四川盆地亞熱帶常綠闊葉林凋落葉多酚類物質在不同降雨期間的降解特征
2018-06-22 10:44:10馬志良楊萬勤吳福忠
生態學報 2018年9期
馬志良, 楊萬勤, 吳福忠, 譚 波
四川省林業生態工程省級重點實驗室, 四川農業大學生態林業研究所, 成都 611130
凋落物分解是重要的森林生態系統過程之一,在維持森林生態系統生產力、凈碳儲量、土壤有機質的形成、森林植被群落演替等方面具有不可替代的作用和地位[1]。凋落物分解過程同時受到氣候、凋落物基質質量、分解者群落等生物和非生物因素的綜合調控[1- 2]。植物多酚,也稱單寧,是植物體合成的一類復雜的次生代謝產物,具多元酚結構,在結構上可分為水解單寧和縮合單寧兩大類,廣泛分布于植物的皮、根、葉和果實中,在許多針葉樹種的樹皮中含量可高達20%—40%,僅次于纖維素、半纖維素和木質素[3]。植物多酚由多環芳烴核和活性官能團聚合而成,相對分子質量大,結構復雜,化學性質活潑[4]。這類物質雖不能通過各種代謝途徑直接影響植物的生長發育,但是其在植物進化過程中扮演著重要作用,如能提高植物對環境的適應性和空間競爭能力、抵御植食性動物的啃食、增強抗病性及化感作用等[5];同時對整個森林生態系統過程具有重要的生態反饋意義[6]。多酚類物質在植物體內不易遷移,不能被植物重新利用,在森林凋落物中,尤其是針葉樹種凋落物,多酚類物質含量常常很高[7]。凋落物多酚類物質含量是一類重要的基質質量指標,其含量的多寡常被用作預測凋落物分解速率的指標[1,7- 9]。一般地,多酚類物質含量越高,凋落物分解速率越慢[10- 11],同時也受到水解單寧和縮合單寧含量相對比例的調控。然而,目前關于凋落物分解的研究主要集中在大量元素釋放動態(如C、N、P等)[12- 13]以及木質素、纖維素降解特征等[14- 15]方面,凋落物分解過程中多酚類物質的降解動態仍不十分清楚。在濕熱的亞熱帶常綠闊葉林區,凋落物分解由生物降解主導,土壤微生物和土壤動物對凋落物難分解成分(木質素、纖維素和多酚等)的降解過程具有顯著貢獻[16- 17]。四川盆地亞熱帶常綠闊葉林每年在秋冬季節出現葉凋落高峰[18],新鮮的凋落葉首先經過幾個月的旱季分解,迎來水熱條件較好的雨季。季節性降雨期間,良好的水熱環境更適宜于生物分解者生長、繁殖和活動,凋落物生物降解過程強烈進行。然而,凋落物的生物降解同時受到分解者群落、凋落物基質質量和降解所處的環境條件的綜合影響[19]。不同分解時期凋落物的基質質量和分解所處的環境不斷發生變化,分解者群落結構與活性也在不斷變化[20]。因此,不同基質質量的凋落葉多酚類物質在不同降雨時期可能具有明顯不同的降解特征。然而,目前仍然缺乏季節性降雨對凋落物多酚類物質降解過程的影響研究[21]。
四川盆地地處長江上游的亞熱帶常綠闊葉林區,具有雨熱同季的氣候特點,干濕季分明,季節性降雨期長達5—6個月。前期研究結果發現,季節性降雨期間,由于相對較好的水、熱等環境條件,凋落物具有更為明顯的分解特征,凋落葉在第一年分解了27.75%—72.09%,季節性降雨期的凋落葉質量損失達到69%以上,顯著高于旱季[22]。因此,凋落葉多酚類物質降解過程在季節性降雨期間可能加快,且受到不同樹種凋落葉基質質量的影響。本文以四川盆地亞熱帶常綠闊葉林最具代表性的3個針葉樹種馬尾松(Pinusmassoniana)、柳杉(Cryptomeriafortunei)、杉木(Cunninghamialanceolata)和3個闊葉樹種香樟Cinnamomumcamphora)、紅椿(Toonaciliata)、麻櫟(Quercusacutissima)凋落葉為研究對象,根據實際降雨特征,采用凋落物袋法研究了6種凋落葉多酚類物質在不同雨熱季節的降解特征,以期為更好的認識亞熱帶常綠闊葉林凋落物分解過程提供基礎理論依據。
1 材料與方法
1.1 研究區概況
本研究區域位于四川省崇州市榿泉鎮四川農業大學現代農業研發基地,該基地占地面積133.33hm2,坐落于長江上游四川省川西平原西部,地理坐標103°38′31′′—103°39′22′′E,30°33′16′′—30°33′54′′N,海拔516m,屬四川盆地亞熱帶濕潤季風氣候。該地區年平均氣溫16.0℃,年平均降水量1015.2mm,降雨主要集中在5—9月,降雪稀少。年平均日照時數為1161.5h,平均無霜期為283d。土壤類型為老沖積黃壤。研究區域土壤性質以及植被狀況詳見文獻[23]。
1.2 試驗方法
1.2.1 試驗設計和樣品處理
2012年10月底在四川盆地典型生態系統用尼龍網收集馬尾松、柳杉、杉木、香樟、紅椿、麻櫟的新鮮凋落葉。收集好的6種凋落葉帶回實驗室,經自然風干后,每個樹種準確稱取10g裝入凋落物袋內(規格為20cm×20cm,網孔大小為底部0.5mm,表面層1mm)并標記,每種凋落葉分別裝15袋,共計90袋。另外分別稱取相同質量(10g)的6種樹種凋落葉各3份,在65℃下烘干至恒量,測定含水量,用于推算放置在凋落物袋內凋落葉的初始干質量。烘干的凋落葉樣品粉碎后用于初始多酚類物質含量及其他基質質量指標的測定。其中,多酚類物質含量采用福林酚法測定。6種凋落葉分解前多酚類物質含量見表1,凋落葉其他初始基質質量特征詳見文獻[22]。
根據前期調查,于2013年1月18日在研究區域內選擇3塊無高大喬木和灌木遮掩、地形平坦的空曠地表作為重復試驗樣地,每塊樣地之間間隔大約10m。首先去除試驗樣地土壤表面的植物和凋落物,再將凋落物袋平鋪于地表,每種凋落物袋在每塊試驗樣地上放置5袋。樣品埋設后,在試驗地地表凋落物袋內設置一個紐扣式溫度記錄器(iButton DS1923-F5, Maxim/Dallas Semiconductor, Sunnyvale, USA),設定為每120min讀取一次溫度數據,自動記錄試驗期間地表的溫度變化(圖1)。根據崇州市多年降雨資料以及試驗期間實際降雨狀況(圖1),將試驗時間劃分為微量降雨期(MRS: 2013年1月18日—2013年2月19日)、春季少雨期(SRS: 2013年2月19日—2013年4月22日)、雨季前期(ERS: 2013年4月22日—2013年8月19日)、雨季后期(LRS: 2013年8月19日—2013年10月22日)和冬季少雨期(WRS: 2013年10月22日—2014年1月18日)5個時期。各時期降雨量數據詳見文獻[22]。具體采樣時間為2013年2月19日、4月22日、8月19日、10月22日和2014年1月18日,每次從每塊樣地內采集每種凋落物袋各1袋,帶回實驗室。小心去除表面泥土和新長入的細根,于65℃烘干至恒量,測定剩余凋落葉的質量,粉碎后測定多酚類物質含量。
1.2.2 數據計算
多酚類物質降解率計算公式為:
E(%) = (M0×C0-Mt×Ct)/ (M0×C0) × 100%;
多酚類物質降解量計算公式為:
R=Mt×Ct-Mt+1×Ct+1;
式中,M0為放置凋落物袋時袋內凋落葉的干質量(g);Mt和Mt+1分別為t和t+1采樣時期凋落物袋內凋落葉的干質量(g);C0為初始多酚類物質含量(mg/g);Ct和Ct+1分別為t和t+1采樣時期多酚類物質含量(mg/g)。
1.3 數據統計分析
使用Excel 2003和SPSS 20.0進行數據統計與分析。使用單因素方差分析(one-way ANOVA)和最小顯著差異法(LSD)比較凋落葉分解前多酚類物質含量和各分解時期多酚類物質降解量之間的差異。當方差不齊時執行不等方差假設項下的Dunnett′s T3差異顯著性檢驗。制圖使用Origin 9.0完成。
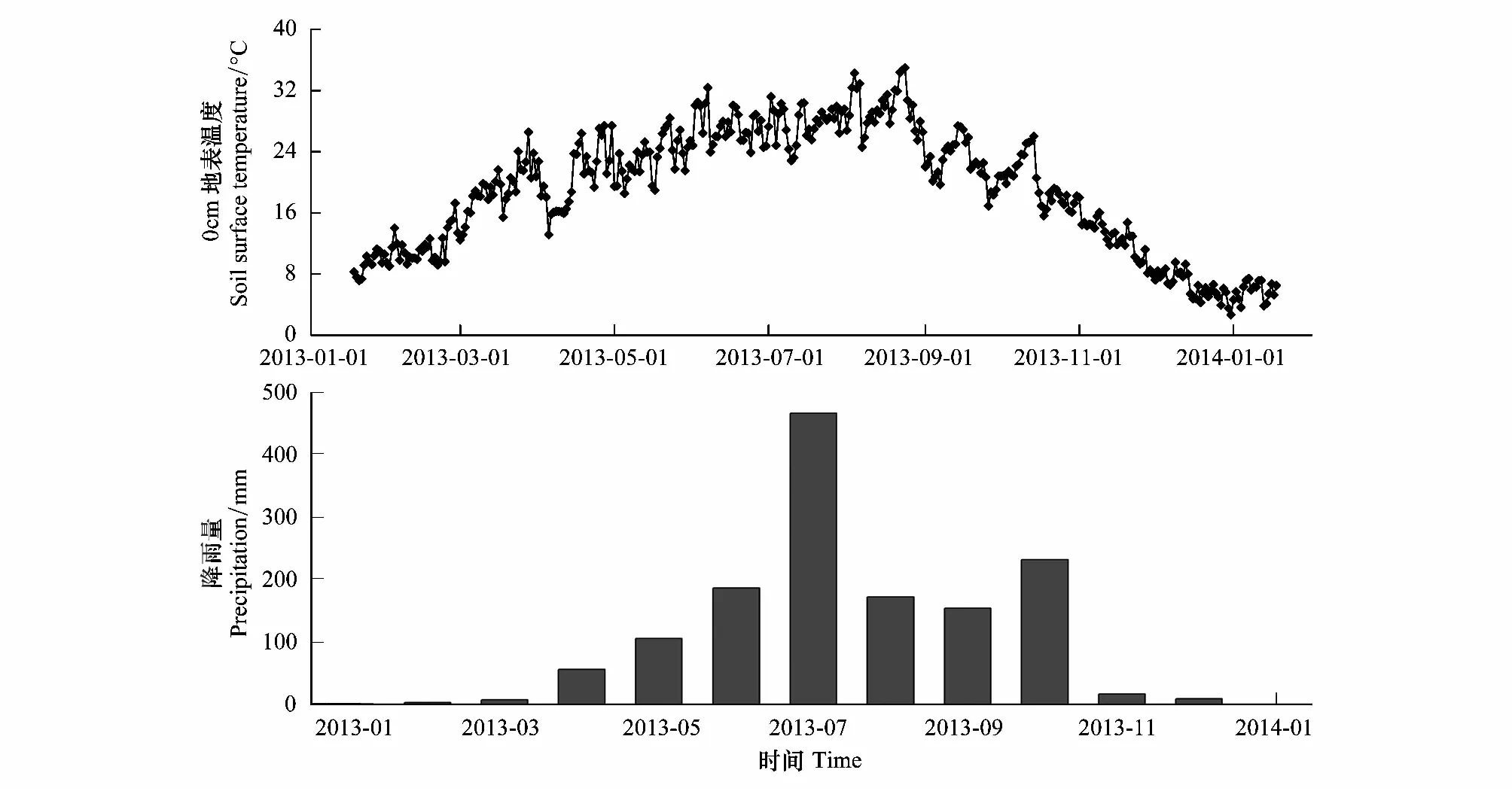
圖1 試驗期間日平均溫度和月實際降水量Fig.1 The daily mean temperature and the month actual precipitation during the incubation experiment
2 結果與分析
2.1 凋落葉多酚類物質含量動態
麻櫟凋落葉多酚類物質含量最高,香樟最低,且6種凋落葉初始多酚類物質含量之間差異顯著(表1)。分解過程中6種凋落葉多酚類物質含量動態較為一致,均表現為自微量降雨期起至雨季前期止,多酚類物質含量均表現為迅速下降的趨勢;雨季前期結束時,紅椿凋落葉多酚類物質降解完畢,其余5種凋落葉多酚類物質含量均降低至5mg/g以下,此后均保持動態平衡(圖2)。

表1 6種凋落葉分解前多酚類物質含量/(mg/g)
不同的小寫字母表示不同樹種之間差異顯著(P<0.05)。數據為平均值±標準差,n=3
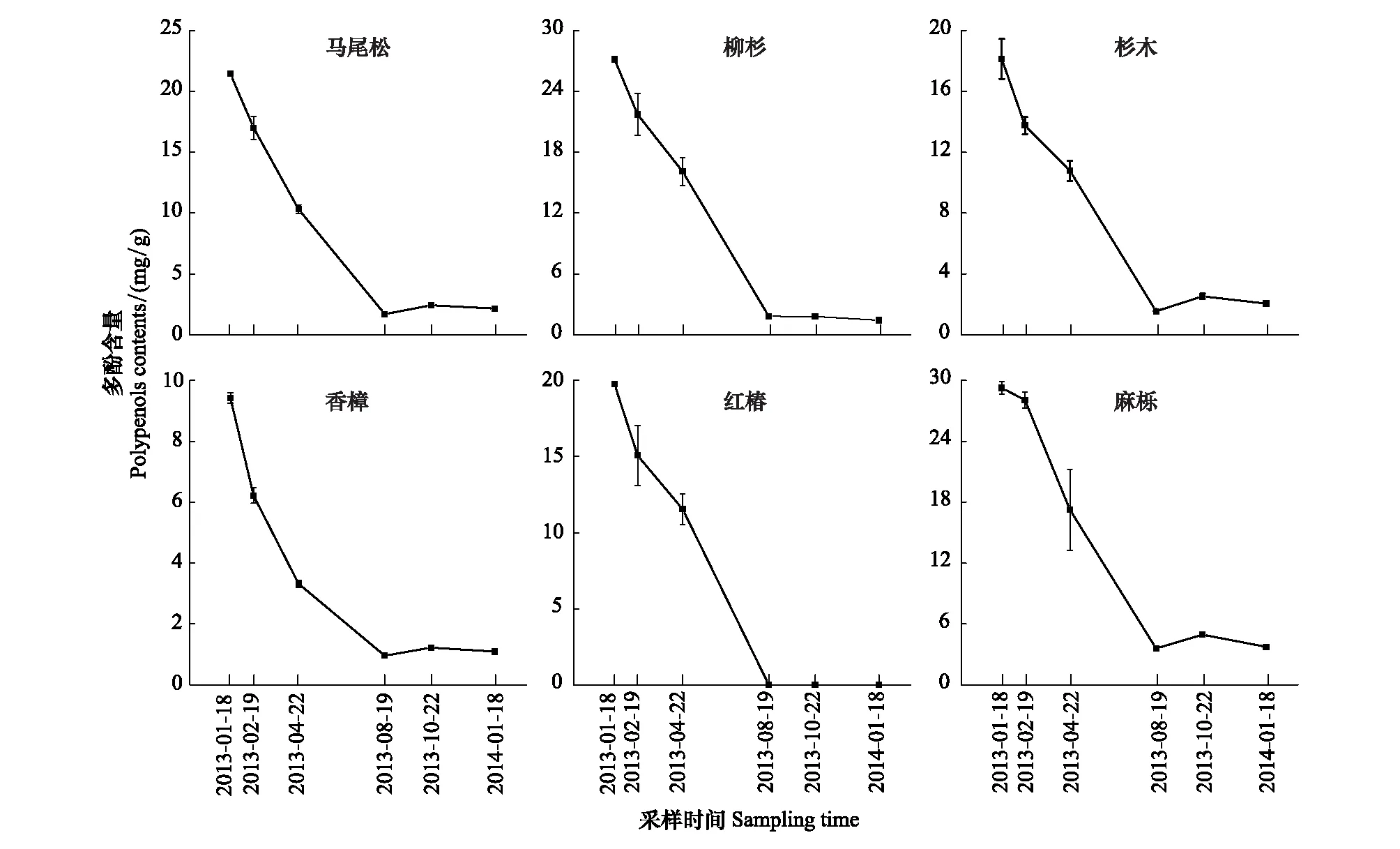
圖2 常綠闊葉林6個樹種凋落葉多酚類物質含量動態Fig.2 Dynamics of polyphenols contents of foliar litter of six tree species in the subtropical evergreen broadleaved forest
2.2 凋落葉多酚類物質降解動態
由圖3可見,與凋落葉多酚類物質含量動態類似,6種凋落葉多酚類物質降解動態變化較為一致,均表現為自分解開始起至雨季前期止,隨降雨量的增加,多酚類物質降解率呈現直線增加的趨勢,之后處于穩定狀態。分解1年后,6種凋落葉多酚類物質降解率大小順序依次為:紅椿(100%) > 柳杉(97.81%) > 杉木(94.45%) > 麻櫟(93.67%) > 馬尾松(93.06%) > 香樟(91.64%);并且除香樟(雨季前期降解率為28.78%)以外的5種凋落葉多酚類物質大量降解發生在雨季前期(ERS),占全年降解率的44.46%—55.72%;6種凋落葉多酚類物質在分解初期旱季兩時期(MRS和SRS)均有較大的降解量,此時期多酚類物質降解率占全年降解率的42.16%—71.20%。由表2可以看出,凋落葉多酚類物質降解受物種和分解時期的調控,不同物種凋落葉多酚類物質在不同分解時期具有不同的降解特征,并且降解量具有顯著差異。此外,凋落物初始多酚類物質含量對其最終降解率的大小具有決定作用。分解1年后,對凋落葉初始多酚類物質含量與其降解率進行函數擬合,發現二者之間呈顯著地二次函數關系(圖4)。
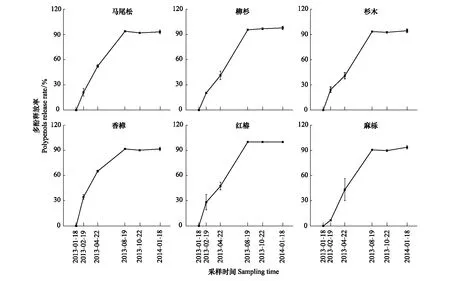
圖3 常綠闊葉林6個樹種凋落葉多酚類物質降解率動態Fig.3 Dynamics of polyphenols degradation rates of foliar litter of six tree species in the subtropical evergreen broadleaved forest
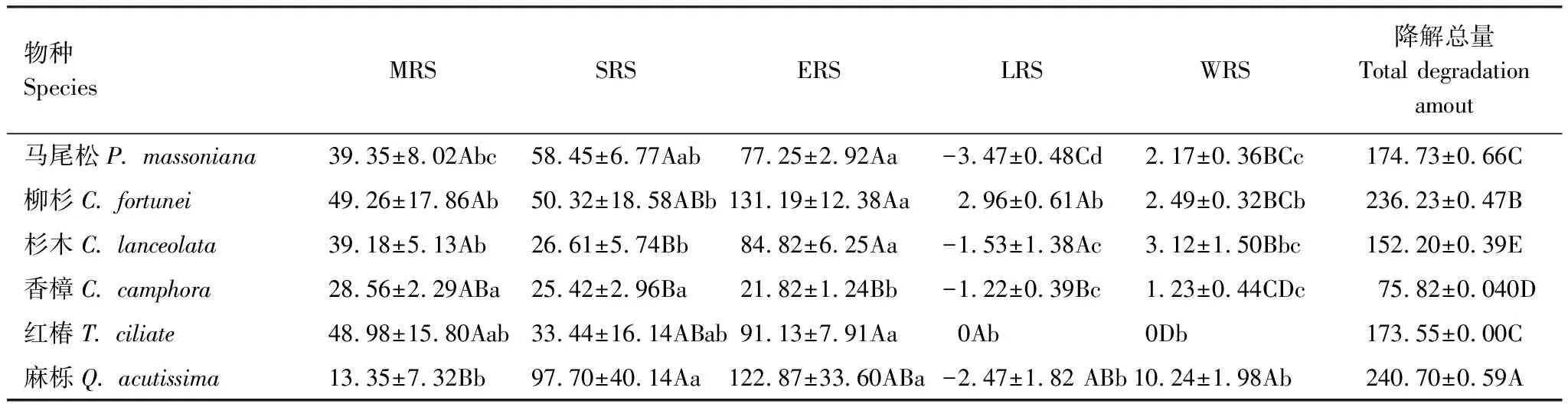
物種SpeciesMRSSRSERSLRSWRS降解總量Total degradation amout馬尾松P. massoniana39.35±8.02Abc58.45±6.77Aab77.25±2.92Aa-3.47±0.48Cd2.17±0.36BCc174.73±0.66C柳杉C. fortunei49.26±17.86Ab50.32±18.58ABb131.19±12.38Aa2.96±0.61Ab2.49±0.32BCb236.23±0.47B杉木C. lanceolata39.18±5.13Ab26.61±5.74Bb84.82±6.25Aa-1.53±1.38Ac3.12±1.50Bbc152.20±0.39E香樟C. camphora28.56±2.29ABa25.42±2.96Ba21.82±1.24Bb-1.22±0.39Bc1.23±0.44CDc75.82±0.040D紅椿T. ciliate48.98±15.80Aab33.44±16.14ABab91.13±7.91Aa0Ab0Db173.55±0.00C麻櫟Q. acutissima13.35±7.32Bb97.70±40.14Aa122.87±33.60ABa-2.47±1.82 ABb10.24±1.98Ab240.70±0.59A
同列不同的大寫字母表示同一分解時期不同物種多酚類物質降解量之間差異顯著(P<0.05),同行不同的小寫字母表示同一物種不同分解時期多酚類物質降解量之間差異顯著(P<0.05)。MRS: 微量降雨期, Micro rainy stage;SRS: 春季少雨期, Spring rainy stage;ERS: 雨季前期, Early stage of rainy season;LRS: 雨季后期, Later stage of rainy season;WRS: 冬季少雨期, Winter brief rainy stage。數據為平均值±標準差,n=3
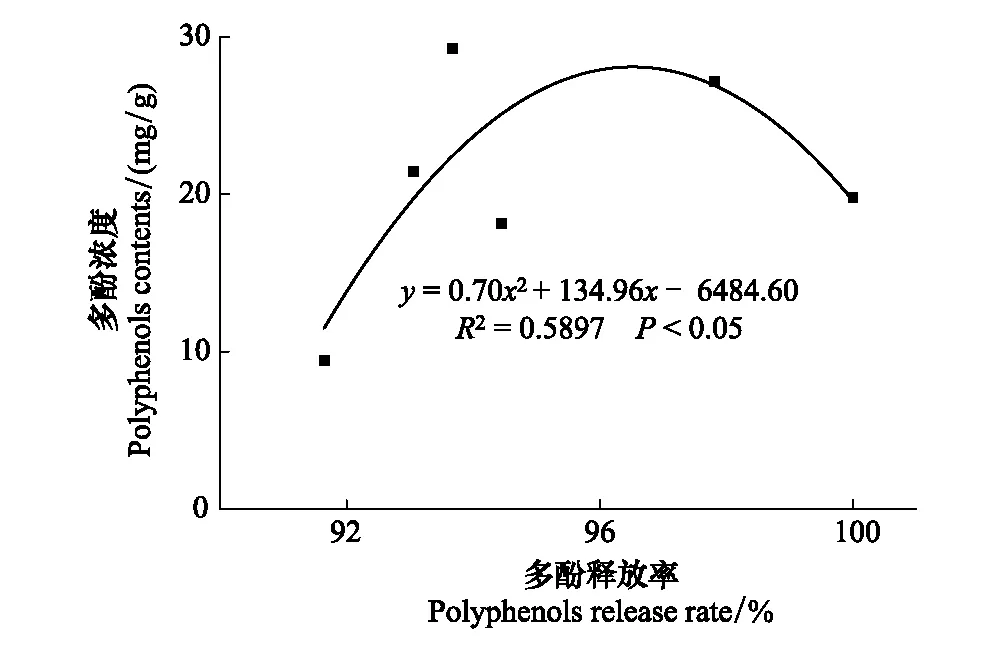
圖4 常綠闊葉林6個樹種凋落葉多酚類物質降解率與多酚類物質初始濃度的關系Fig.4 Relationships between polyphenols degradation rates and initial polyphenols contents of foliar litter of six tree species in the subtropical evergreen broadleaved forest
3 討論
植物多酚類物質以凋落物和根系分泌物的形式進入到土壤后能改善土壤環境,提高微生物活性,有利于提高土壤中營養元素的有效性[24- 25],對植物-凋落物-土壤之間的相互作用產生重要影響。植物多酚類物質是凋落物的主要組分,它可通過化感作用或者形成蛋白-單寧復合體“屏障”,阻礙分解者活動,抑制凋落物分解過程,進而促進土壤腐殖質的形成,將更多的有機碳固定在土壤中[7]。亞熱帶常綠闊葉林凋落物分解過程由生物降解主導,凋落物的生物降解其實質是凋落物在分解酶系統作用下的生物化學過程[22, 26]。多酚氧化酶是凋落物多酚類物質降解的關鍵酶[6],它主要來源于土壤微生物與土壤動物釋放的胞外酶,能將植物殘體中多酚類物質轉化為腐殖質組分,同時也參與木質素的降解[27- 28]。降水、溫度等通過影響分解者群落結構與活性控制著整個分解過程。然而,本研究發現,6種凋落葉在分解初期(微量降雨期和春季少雨期)多酚類物質含量明顯減少(圖2),多酚類物質在這兩個時期也呈現出明顯的降解釋放過程,占全年降解率的42.16%—71.20%(圖3和表2)。凋落葉分解初期,降水量少,溫度較低,這均不利于土壤微生物和動物的活動;此時期內多酚氧化酶的活性也較低,更加不利于多酚類物質的生物降解[29]。凋落物分解初期多酚類物質呈現大量降解釋放的原因可能是由于低溫和干旱導致的機械破碎所造成的物理損失。前期研究發現,此時期機械破碎導致凋落葉出現明顯的質量損失[22],同時對凋落葉木質素和纖維素的降解也有一定的貢獻[21]。故凋落葉分解初期強烈的機械破碎同時會導致這一時期多酚類物質的降解損失[30]。隨著分解時間的進行,凋落葉分解進入水熱條件較好的雨季,大量的土壤微生物和動物在凋落葉上聚集、生長、繁殖,加速凋落葉的分解進程[22]。與其他分解時期相比,雨季前期凋落葉多酚類物質的含量呈現出急劇減少的現象,凋落葉多酚類物質降解釋放也大量發生于此時期(圖2、圖3和表2)。其原因可能有以下幾點:(1) 分解初期,強烈的機械破碎會嚴重破壞凋落葉中蛋白-單寧復合體結構,多酚類物質失去“屏障”的保護,有利于多酚類物質在雨季時期的降解[7];(2) 季節性降雨期間,凋落物分解所處的水、熱條件較好,土壤生物活動頻繁加速多酚類物質的降解;(3) 土壤微生物大量生長,釋放出更多的多酚氧化酶,同時多酚氧化酶在該時期內具有較高的活性[31],有利于其降解;(4) 凋落葉分解過程中產生的小分子量酚類物質隨即被雨季前期較多的降水淋溶到土壤中[32]。至雨季前期結束時,凋落葉多酚類物質的含量降低至一個極低的水平(5mg/g),此后一直處于動態平衡狀態,在雨季后期和冬季少雨期,6種凋落葉多酚類物質含量趨于穩定,均沒有表現出明顯的降解特征(圖3),這可能是由于多酚類物質形成了穩定的腐殖質的緣故[33- 34]。
本研究發現,凋落葉多酚類物質降解過程受樹種組成和分解時期的影響,不同樹種凋落葉在不同分解時期降解量差異顯著(表2)。這是因為不同樹種凋落葉所含的多酚類物質種類及其相對比例明顯不同,其結構存在很大差異[35],因此在不同的分解時期呈現出明顯不同的降解釋放特征。然而,本研究并未區分鑒定6種凋落葉多酚類物質的具體組分,以及水解單寧和縮合單寧的相對比例,以后應加強凋落物分解過程中多酚類物質不同組分對季節性降雨的響應等相關研究。樹種組成影響著凋落物不同組分的初始含量,對凋落物初始基質質量具有決定作用,進而調節凋落物整個分解過程[20, 36]。本研究也發現凋落葉多酚類物質的初始濃度與其最終降解率呈現出顯著的二次函數關系(圖4)。而不同分解時期則是通過影響降雨、溫度等環境因子調節土壤生物群落結構與活性以及胞外酶活性進而作用于凋落葉的分解過程[22]。
綜上所述,四川盆地亞熱帶常綠闊葉林6個樹種凋落葉多酚類物質降解在各個分解時期呈現出明顯不同的動態變化特征。受季節性降雨和樹種因素共同調控,凋落葉多酚類物質自微量降雨期起至雨季前期止,多酚類物質含量迅速下降;在降水量較多的雨季前期,凋落葉多酚類物質降解量較大。由此可見,降水量是濕潤亞熱帶常綠闊葉林凋落物多酚類物質降解釋放的關鍵驅動因子之一,樹種組成是影響凋落葉多酚類物質降解動態的內部因子。以上結果可為更加清晰地認識亞熱帶常綠闊葉林凋落物分解過程提供一定的理論依據。
參考文獻(References):
[1] 楊萬勤, 鄧仁菊, 張健. 森林凋落物分解及其對全球氣候變化的響應. 應用生態學報, 2007, 18(12): 2889- 2895.
[2] Berg B, McClaugherty C. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration. Berlin, Germany: Springer, 2003.
[3] 孫達旺. 植物單寧化學. 北京: 中國林業出版社, 1992.
[4] 張力平, 孫長霞, 李俊清, 劉艷紅. 植物多酚的研究現狀及發展前景. 林業科學, 2005, 41(6): 157- 162.
[5] 秦小玲. 北方干旱地區幾種植物多酚含量動態研究[D]. 北京: 北京林業大學, 2006.
[6] 鐘哲科, 王人潮, 江波. 森林土壤有機質層中多酚類物質的生態反饋意義. 應用生態學報, 2003, 14(3): 341- 344.
[7] Northup R R, Dahlgren R A, McColl J G. Polyphenols as regulators of plant-litter-soil interactions in northern California′s pygmy forest: a positive feedback? Biogeochemistry, 1998, 42(1/2): 189- 220.
[8] Kazakou E, Vile D, Shipley B, Gallet C, Garnier E. Co-variations in litter decomposition, leaf traits and plant growth in species from a Mediterranean old-field succession. Functional Ecology, 2006, 20(1): 21- 30.
[9] Aerts R, De Caluwe H, Beltman B. Plant community mediated vs. nutritional controls on litter decomposition rates in grasslands. Ecology, 2008, 84(12): 3198- 3208.
[10] Kalburtji K L, Mosjidis J A, Mamolos A P. Litter dynamics of low and high tannin sericea lespedeza plants under field conditions. Plant and Soil, 1999, 208(2): 271- 281.
[11] Driebe E M, Whitham T G. Cottonwood hybridization affects tannin and nitrogen content of leaf litter and alters decomposition. Oecologia, 2000, 123(1): 99- 107.
[12] Ndibe T O, Onwumere G B, Bulus T. Litter decomposition and release of nutrients formEucalyptuscamaldulensisleaf on Eucalypt plantation soils. World Scientific News, 2017, 62: 24- 45.
[13] Ibrahima A, Ntonga J C, Ze A D M. Litter decomposition and nutrient dynamics in tropical rainforests of Ebom, Southwestern Cameroon: effects of logging-disturbed. Global Journal of Botanical Science, 2016, 4(1): 24- 36.
[14] He W, Wu F Z, Yang W Q, Tan B, Zhao Y Y, Wu Q Q, He M. Lignin degradation in foliar litter of two shrub species from the gap center to the closed canopy in an alpine fir forest. Ecosystems, 2016, 19(1): 115- 128.
[15] Yue K, Wu F Z, Yang W Q, Zhang C, Peng Y, Tan B, Xu Z F, Huang C P. Cellulose dynamics during foliar litter decomposition in an alpine forest meta-ecosystem. Forests, 2016, 7(8): 176- 176.
[16] M?kel? M R, Lundell T, Hatakka A, Hildén K. Effect of copper, nutrient nitrogen, and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungusPhlebiaradiata. Fungal Biology, 2013, 117(1): 62- 70.
[17] 卜濤, 張水奎, 宋新章, 江洪. 幾個環境因子對凋落物分解的影響. 浙江農林大學學報, 2013, 30(5): 740- 747.
[18] 鐘章成. 常綠闊葉林生態系統研究. 重慶: 西南師范大學出版社, 1992.
[19] Torres I F, Bastida F, Hernández T, Bombach P, Richnow H H, García C. The role of lignin and cellulose in the carbon-cycling of degraded soils under semiarid climate and their relation to microbial biomass. Soil Biology and Biochemistry, 2014, 75: 152- 160.
[20] García-Palacios P, McKie B G, Handa I T, Frainer A, H?ttenschwiler S. The importance of litter traits and decomposers for litter decomposition: a comparison of aquatic and terrestrial ecosystems within and across biomes. Functional Ecology, 2016, 30(5): 819- 829.
[21] 馬志良, 高順, 楊萬勤, 吳福忠. 亞熱帶常綠闊葉林區凋落葉木質素和纖維素在不同雨熱季節的降解特征. 生態學雜志, 2015, 34(1): 122- 129.
[22] 馬志良, 高順, 楊萬勤, 吳福忠, 譚波, 張璽濤. 亞熱帶常綠闊葉林6個常見樹種凋落葉在不同降雨期的分解特征. 生態學報, 2015, 35(22): 7553- 7561.
[23] 馬志良, 高順, 楊萬勤, 吳福忠. 亞熱帶常綠闊葉林凋落葉分解過程中氮和磷在不同雨熱季節的釋放動態. 應用與環境生物學報, 2015, 21(2): 308- 315.
[24] 安樹青. 濕地生態工程. 北京: 化學工業出版社, 2003.
[25] 付美云, 周立祥. 植物多酚在環境保護與農業生產中的應用. 應用生態學報, 2004, 15(9): 1673- 1677.
[26] García-Palacios P, Shaw E A, Wall D H, H?ttenschwiler S. Temporal dynamics of biotic and abiotic drivers of litter decomposition. Ecology Letters, 2016, 19(5): 554- 563.
[27] 宋影, 辜夕容, 嚴海元, 毛文韜, 吳雪蓮, 萬宇軒. 中亞熱帶馬尾松林凋落物分解過程中的微生物與酶活性動態. 環境科學, 2014, 35(3): 1151- 1158.
[28] 趙恒毅, 耿玉清, 楊英, 周紅娟, 張海蘭, 王玲, 趙廣亮. 北京低山區油松林和元寶楓林凋落物酶活性研究. 中南林業科技大學學報, 2016, 36(6): 23- 28.
[29] Veres Z, Kotroczó Z, Fekete I, Tóth J A, Lajtha K, Townsend K, Tóthmérész B. Soil extracellular enzyme activities are sensitive indicators of detrital inputs and carbon availability. Applied Soil Ecology, 2015, 92: 18- 23.
[30] Martins R T, Melo A S, Gon?alves J F Jr, Hamada N. Leaf-litter breakdown in urban streams of Central Amazonia: direct and indirect effects of physical, chemical, and biological factors. Freshwater Science, 2015, 34(2): 716- 726.
[31] 郝建朝, 吳沿友, 連賓, 吳春篤. 土壤多酚氧化酶性質研究及意義. 土壤通報, 2006, 37(3): 470- 474.
[32] 陸麗芳. 珠芽蓼凋落物中酚類物質分解動態研究[D]. 蘭州: 蘭州大學, 2015.
[33] 高艷, 馬紅亮, 高人, 尹云鋒, 陳仕東, 章偉, 朱祥妹, 楊玉盛. 模擬氮沉降對森林土壤酚類物質和可溶性糖含量的影響. 土壤, 2014, 46(1): 41- 46.
[34] Ni X Y, Yang W Q, Tan B, He J, Xu L Y, Li H, Wu F Z. Accelerated foliar litter humification in forest gaps: dual feedbacks of carbon sequestration during winter and the growing season in an alpine forest. Geoderma, 2015, 241- 242: 136- 144.
[35] Halvorson J J, Gonzalez J M, Hagerman A E, Smith J L. Sorption of tannin and related phenolic compounds and effects on soluble-N in soil. Soil Biology and Biochemistry, 2009, 41(9): 2002- 2010.
[36] Veen G F, Sundqvist M K, Wardle D A. Environmental factors and traits that drive plant litter decomposition do not determine home-field advantage effects. Functional Ecology, 2015, 29(7): 981- 991.
