獨山子區優勢草本植物根際與非根際土壤微生物功能多樣性
2018-06-22 11:29:30呂光輝
生態學報 2018年9期
陳 悅,呂光輝,李 巖
1 新疆大學資源與環境科學學院,烏魯木齊 830046 2 綠洲生態教育部重點實驗室,烏魯木齊 830046 3 新疆大學干旱生態環境研究所,烏魯木齊 830046
土壤微生物是生物地球化學循環的引擎[1],它在維持生態系統整體服務功能方面具有重要作用,常被比作土壤養分元素循環的“轉化器”、環境污染物的“凈化器”以及陸地生態系統穩定的“調節器”[2]。土壤生態系統中所有的微生物種類、基因以及微生物與環境之間相互作用的多樣化程度便是土壤微生物的多樣性。而土壤微生物功能多樣性是土壤微生物群落狀態與功能的重要指標之一[3],包括分解、營養傳遞以及促進或抑制植物生長的功能,對明確不同環境中微生物群落的作用以及土壤生態功能具有重要意義。
土壤微生物多樣性通常以土壤微域結構和空間分異這樣的環境條件而被直接影響,其中植物根際是土壤微域結構的主要類型之一[4]。目前,根際土壤微生物的研究主要集中在鹽堿地、山區、草地等[5- 7]自然生態系統中,農田連作、施肥和受污染土壤等[8- 10]人為干擾的非自然生態系統中,主要通過對植物根際土壤的微生物量、群落結構及多樣性等研究,發現因地域、環境、土地利用方式以及生長時間的不同而得到不同的根際土壤微生物特征,但對干旱區土壤微生物功能多樣性的研究,鮮見報道。
新疆克拉瑪依市獨山子區是西部重要的石化基地和油氣引進、加工、儲轉的戰略樞紐,也是天山北坡經濟帶重點發展地區。獨山子石油從開采到現今已有近120年。長期的石油加工作業,使得與工業活動相關的土壤問題變得日益突出,生態恢復迫在眉睫。生態系統的恢復不僅要考慮植物多樣性,更要關注土壤微生物多樣性,土壤微生物多樣性對土壤結構及團聚體的形成、土壤穩定性和肥力、有機物質的轉化具有重要意義[11]。植被的存在,為土壤微生物提供其生存的營養物質與能量,并通過影響土壤有機碳、氮和磷的水平、土壤含水量、溫度、孔隙度及pH值等來影響土壤微生物多樣性[12]。因此,研究植被對土壤微生物群落的影響,揭示不同植被與土壤微生物群落的關系,有助于選擇合適的植被類型進行生態恢復和重建。
本研究以獨山子區三種優勢草本植物的根際與非根際土壤為研究對象,采用Biolog-ECO微平板法,對不同植物根際與非根際土壤化學性質、微生物平均顏色變化率、微生物多樣性指數、微生物代謝多樣性類型、微生物多樣性與土壤化學性質的關系進行研究。通過土壤微生物功能多樣性探討,了解當地優勢草本植物根際土壤微生物功能多樣性,旨在為獨山子區生態環境建設與改善提供一定理論依據。
1 研究區概況
研究區位于克拉瑪依市獨山子區石化工廠周邊具有原生態草場區域。獨山子區位于84°43′—85°06′E,44°07′—44°23′N,平均海拔400m,年均氣溫8.1℃,年降水量108.9mm,年均蒸發量達3008.9mm,為降水量的20.8倍,屬典型溫帶大陸性氣候。干旱少雨、春秋多風是其突出的氣候特征。研究區域主要土質為荒漠灰鈣土和灰棕荒漠土,春夏季主要優勢植物有建群種博樂蒿(Artemisiaboratalensis)、駱駝刺(Alhagisparsifolia)及短命植物旱麥草(Eremopyrumtriticeum)、四齒芥(Tetracmequadricornis)、叉毛蓬(Petrosimoniasibirica)、庭薺(Alyssumdesertorum)、澀薺(Malcolmiaafricana)、角果藜(Ceratocarpusarenarius)等。
2 材料與方法
2.1 試驗設計
研究樣地位于獨山子石化工廠下風向2—3km范圍的區域,通過樣地植被調查,根據重要值選取3種不同科屬的優勢草本植物作為研究樣本,包括博樂蒿、旱麥草和四齒芥,它們的重要值依次為34.70%、23.50%、11.89%,在下文中分別以BLH、HMC和SCJ表示。
樣方設置為隨機劃分的15塊3m×3m的區組,區組之間距離≥2m,每塊樣地中包含以上3種優勢草種各至少6株。
2.2 土樣采集
土壤樣品于2016年5月25日采集,植物根際土參照Riley和Barber的抖落法[13],非根際土壤取植物根際投影范圍外自地面垂直0—15cm范圍內的土樣。在每個樣方中隨機選擇大小適中的3種植物各4株,取其根際與非根際土,分別混合均勻,裝于無菌瓶中,置于便攜式車載冰箱,在低溫條件下運回實驗室,儲藏于-18℃的環境,用于后續功能多樣性和理化性質測定。
2.3 測定方法
土壤微生物代謝活性和功能多樣性采用Biolog-ECO微平板法進行測定[14]。
操作步驟:稱取相當于10g烘干土壤重量的新鮮土樣于無菌250mL三角瓶中,加入90mL無菌NaCl溶液(0.85%),封口后,在搖床振蕩15min,靜置澄清,取上清液,在超凈工作臺中用無菌NaCl溶液(0.8%)稀釋到10-3,用8通道加樣器將稀釋后的菌液加入Biolog生態培養板上,每孔加150μL。將接種的Biolog-ECO板放在28℃培養箱中培養8d,每隔24h用ELX- 808型酶標儀讀取培養板在590nm波長的吸光值。
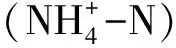
2.4 數據處理
所得數據采用以下公式進行計算分析:
(1)土壤微生物的代謝活性用每孔顏色平均變化率(Average Well Color Development,AWCD)來計算:
AWCD=∑Ci-R/31
式中,Ci為各反應孔在590nm下的光密度值;R為對照孔A1的光密度值;31為Biolog-ECO生態板的C源數目。Ci-R小于零的孔,在計算中記為0,即:Ci-R≥0;
(2)Simpson指數(D):
D=1/∑Pi2
式中,Pi=(Ci-R)/∑(Ci-R),表示有碳源的孔與對照孔A1的光密度值之差與整板總差的比值。
(3)Shannon指數(H):
H=-∑(Pi×lnPi)
(4)McIntosh指數(U),用于評估群落物種均勻度:
數據采用Microsoft Excel 2010、SPSS 22.0以及Canoco 4.5軟件計算分析,通過方差分析(Analysis Of Variance, ANOVA)、相關分析(Correlation Analysis)、主成分分析(Principal Component Analysis, PCA)以及冗余分析(Redundancy Analysis, RDA)進行數據處理,所有數據為15次重復的平均值。
3 結果分析
3.1 不同植物根際與非根際土壤化學性質的變化
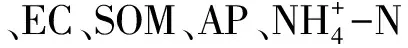
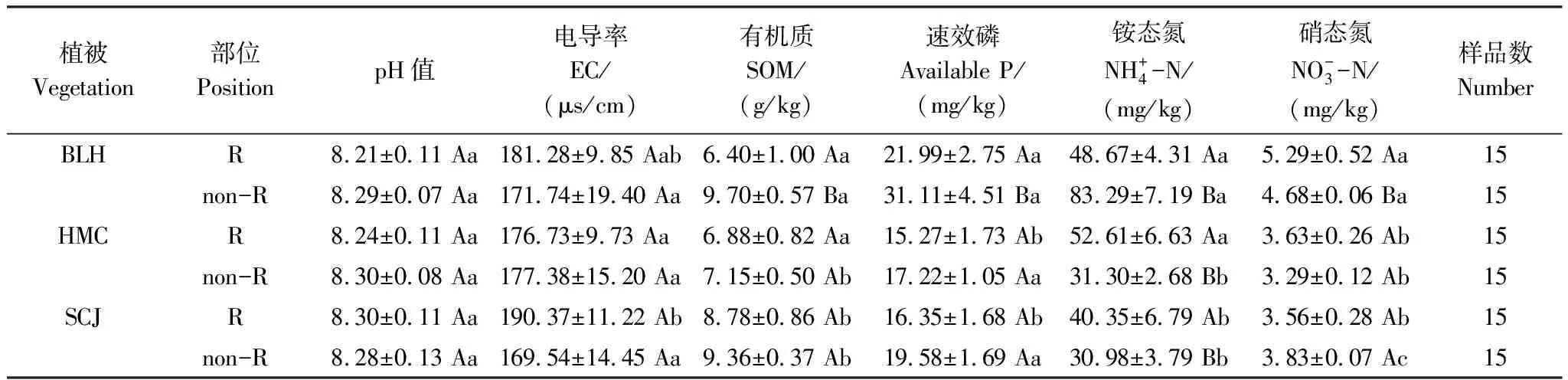
表1 不同植物根際與非根際土壤化學性質
BLH:博樂蒿,Artemisiaboratalensis;HMC:旱麥草,Eremopyrumtriticeum;SCJ:四齒芥,Tetracmequadricornis;EC:電導率,Electrical conductance;SOM:土壤有機質,Soil organic matter;R:根際土壤,Rhizosphere soil;non-R:非根際土壤,non-rhizosphere soil;不同小寫字母表示各植物同一部位土壤理化性質差異顯著(P<0.05),不同大寫字母表示同一種植物不同部位土壤理化性質差異顯著(P<0.05),數值為平均值±標準差(Mean±SD)
3.2 不同植物根際與非根際土壤微生物AWCD變化
平均顏色變化率(AWCD)是一個反映土壤微生物活性,表征利用單一碳源能力的重要指標[16],在一定程度上能夠反映土壤中微生物種群的數量和結構特征。AWCD值越大,表示微生物活性越高,反之越低。
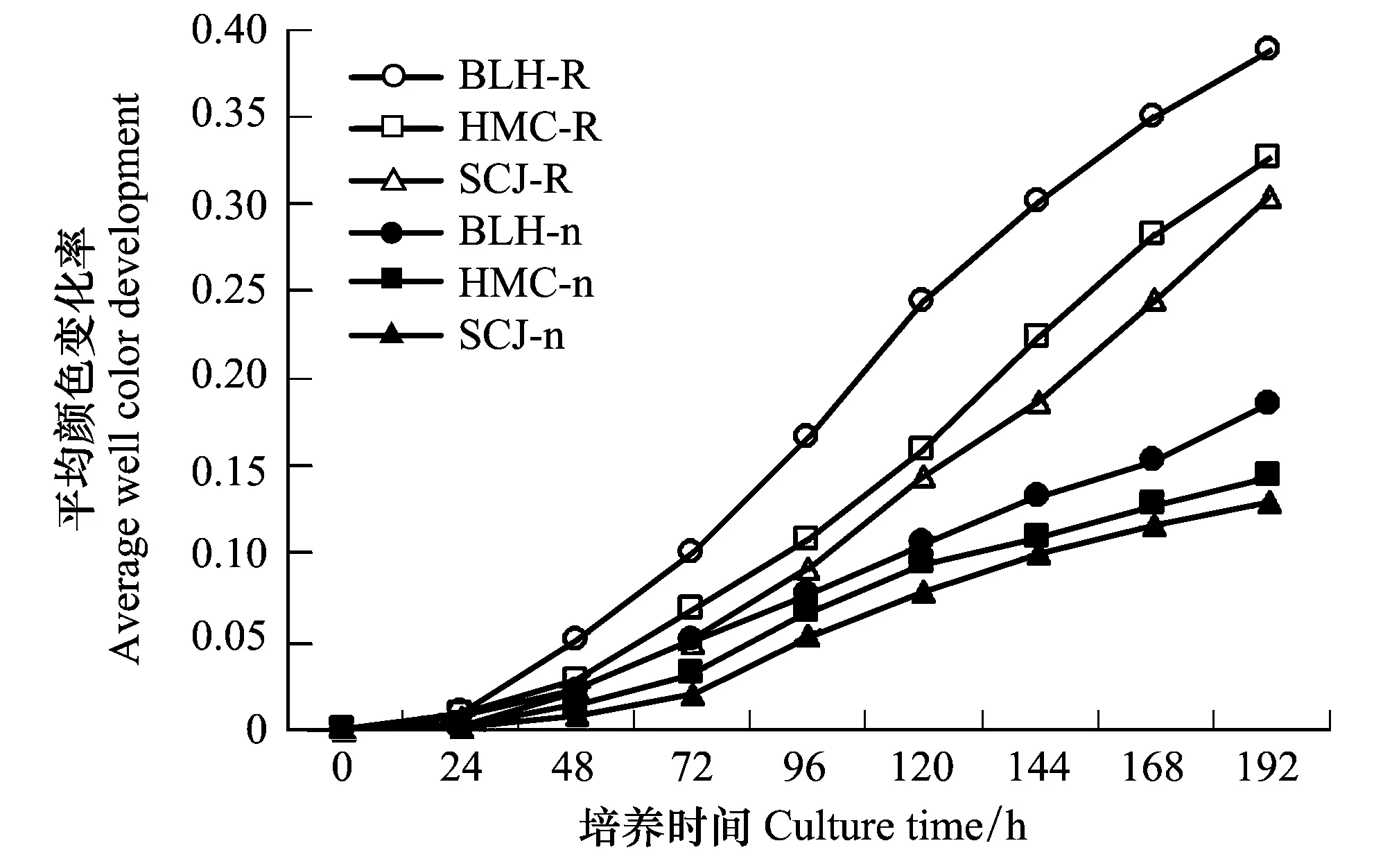
圖1 不同植物根際與非根際土壤微生物AWCD隨培養時間的變化Fig.1 Average well color development (AWCD) between rhizosphere and non-rhizosphere soil of different speciesBLH:博樂蒿,Artemisia boratalensis;HMC:旱麥草,Eremopyrum triticeum;SCJ:四齒芥,Tetracme quadricornis;R:根際土壤,Rhizosphere soil;n:非根際土壤,non-rhizosphere soil
由圖1可知,3種植物類型中AWCD值均隨時間延長而增加,表明不同植物土壤的微生物活性都隨時培養時間的增加而增加,并且各植物根際土壤的AWCD值在24h內無明顯變化,在24h后急劇上升,根際土上升速度快于非根際土。AWCD值表現為:BLH>SCJ>HMC。
3.3 不同植物根際與非根際土壤微生物多樣性指數變化
根據不同植物根際與非根際碳源利用情況,綜合考慮其變化趨勢,選取光密度趨于穩定的120h的光密度值進行土壤微生物群落代謝多樣性分析。從表2中看到,BLH和HMC的根際土壤微生物Simpson指數均顯著小于非根際土(P<0.05),SCJ根際土壤Simpson最大;Shannon指數中,BLH與HMC根際土壤微生物Shannon均大于非根際土,而SCJ顯著小于(P<0.05);BLH非根際土McIntosh指數顯著大于HMC與SCJ(P<0.05),SCJ根際土壤均勻度顯著小于非根際土(P<0.05),其他比較無顯著差異。總體來看,BLH根際土壤微生物群落豐富度與均勻度最好,相比HMC和SCJ具有較寬的微生物種群生態位。
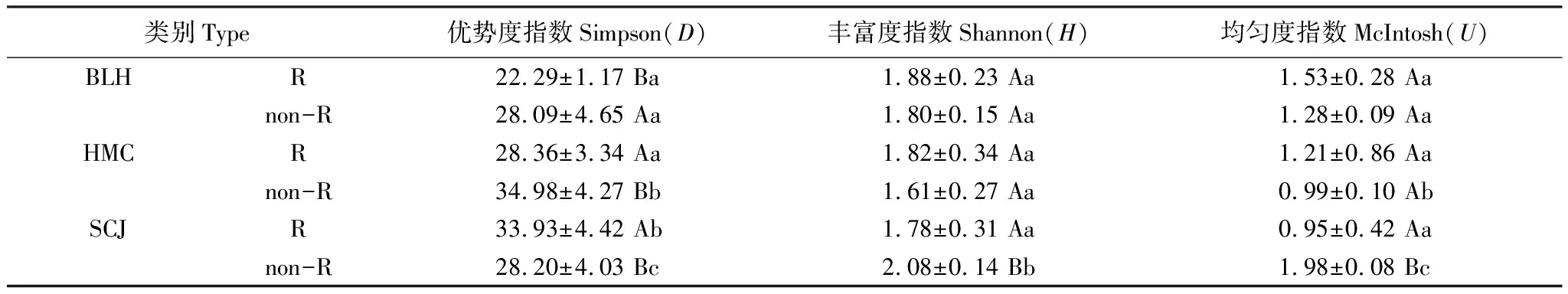
表2 不同植物根際與非根際土壤微生物功能多樣性指數(120h)
R與non-R,不同大寫、小寫字母與數值同上
3.4 不同植物根際與非根際土壤微生物代謝多樣性類型的變化
運用SPSS軟件對培養120h后的土壤微生物進行主成分分析。數據矩陣包括15行代表獨山子試驗區的15個樣地,31列代表生態板上分布的31種不同的碳源物質。
3種優勢草本植物根際土壤微生物碳源的利用結果見表3,共提取了3個主成分,累計貢獻率達92.225%。第1主成分(PC1)的方差貢獻率為48.515%,第2主成分(PC2)為30.398%,第3主成分(PC3)為13.312%。對第1主成分(PC1)貢獻較大的C源有12種,為β-甲基D-葡萄糖苷、D-半乳糖酸γ內酯、D-半乳糖醛酸、吐溫80、N-乙酰基-D-葡萄胺、r-羥基丁酸、D-纖維二糖、D,L-a-甘油、a-環狀糊精、D-氨基葡萄糖酸、葡萄糖- 1-磷酸鹽、以及腐胺;對第2主成分(PC2)貢獻較大的C源有5種,為D-木糖、吐溫40、I-赤藻糖醇、L-蘇氨酸和肝糖;對第3主成分(PC3)貢獻較大的有甘氨酰-L-谷氨酸、苯乙基胺和a-D-乳糖3種。上述主成分中包含的碳源類型有糖類7種,脂類、酸類和胺類各3種,醇類、氨基酸類各包含2種,主要碳源利用類型是糖類、脂類、酸類以及胺類,表明這3種植物根際土壤微生物對上述4類碳源比較敏感。

表3 3種優勢草本植物根際土壤微生物碳源各個主成分特征值和累計貢獻率
3種優勢草本植物非根際土壤微生物碳源的利用結果見表4,共提取了3個主成分,累計貢獻率達92.470%。第1主成分(PC1)的方差貢獻率為63.417%,第2主成分(PC2)為17.323%,第3主成分(PC3)為11.730%。其中對第1主成分(PC1)貢獻較大的C源有14種,為β-甲基D-葡萄糖苷、D-半乳糖酸γ內酯、L-精氨酸、D-木糖、D-半乳糖醛酸、I-赤藻糖醇、2-羥苯甲酸、4-羥基苯甲酸、L-絲氨酸、r-羥基丁酸、D-氨基葡萄糖酸、衣康酸以及腐胺;對第2主成分貢獻較大的C源有5種,分別是L-苯基丙氨酸、a-環狀糊精、N-乙酰基-D-葡萄胺、L-蘇氨酸和a-丁酮酸;對第3主成分貢獻較大的有肝糖和苯乙基胺2種。以上主成分中的貢獻碳源類型包括酸類7種,氨基酸類5種,糖類4種,胺類3種,脂類與醇類各1種。主要碳源利用類型是酸類、氨基酸類以及糖類,表明非根際土壤微生物對上述3種碳源比較敏感。

表4 3種優勢草本植物非根際土壤微生物碳源各個主成分特征值和累計貢獻率
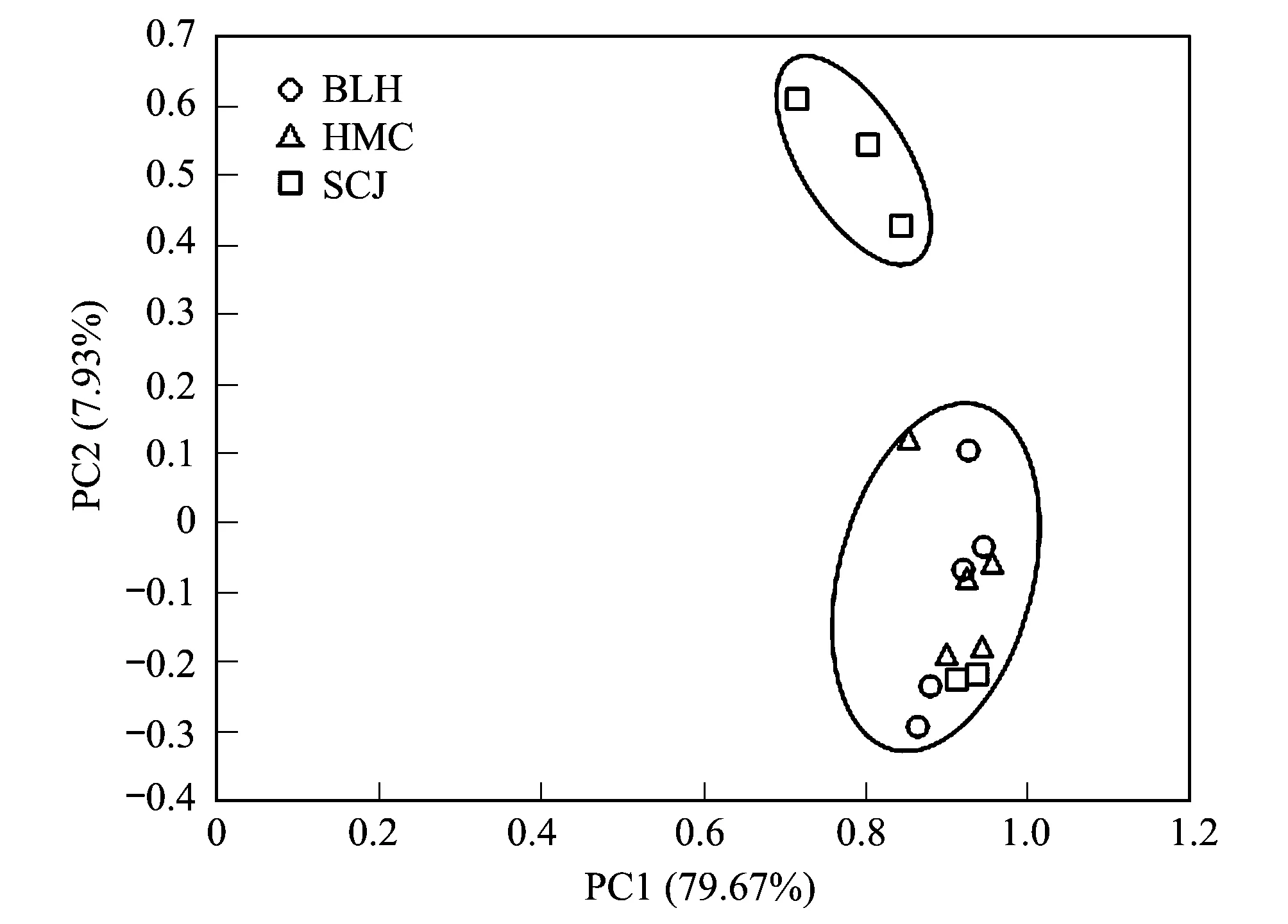
圖2 不同草本植物土壤微生物群落功能的主成分分析 Fig.2 Principal component analysis of soil microbial community function at different plants
對3種植物根際與非根際土壤微生物碳源物質利用情況進行主成分分析,得到兩個主成分,分別為PC1(79.67%)和PC2(7.93%),累計解釋量為87.59%,可以解釋BLH、HMC及SCJ這3種植物在碳源物質利用上差異的大部分信息。如圖2所示,不同植物根際環境微生物對碳源的利用存在較明顯的空間分異,可見植物根際環境微生物代謝特征受不同植物類型的顯著影響。其中BLH與HMC之間碳源利用差異不顯著。
3.5 不同植物根際與非根際土壤微生物多樣性與土壤化學性質的相關關系
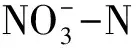
4 討論
本試驗通過Biolog-ECO微平板法對獨山子石化工業區周邊3種優勢草本植物根際與非根際土壤微生物功能多樣性進行研究,發現不同植物之間的土壤微生物活性(AWCD)、優勢度指數Simpson(D)、豐富度指數Shannon(H)、均勻度指數McIntosh(U)均存在一定差異。其中,博樂蒿相比其他兩種植物根際土微生物具有更好的微生物代謝能力、豐富度與多樣性,這是因為BLH具有較長的主根,側根也較發達,根系越旺盛,根際土壤中便會有更多種類和數量的根系分泌物和脫落物,且能夠減少外部環境對土壤微生物功能多樣性的影響[17],當然,不同的植被類型也是造成差異的原因之一。Melany發現不同植被類型能夠影響土壤微生物活性和功能組成格局[18]。Kaiser[19]、Smalla[20]等人也發現同一地區不同植被類型、同一植物不同基因型或同一基因型不同發育階段的根際土壤微生物多樣性均有不同。以上報道均與本研究結果一致,導致差異的原因可能與植被組成、植物殘體、根系分泌物和土壤理化性質等生態因子有關[21]。也表明,植物根際土壤微生物功能多樣性不僅與植物類型有關,在不同生長階段也依然存在差異,具有一定時空特征。

表5 石化工業區優勢草本植物根際與非根際土壤化學性質與微生物功能多樣性的相關關系
*表示P<0.05;**表示P<0.01
研究結果還顯示,不同植物根際土壤微生物各指數均大于自身非根際土,這可能是因為植物根系和植物殘體給根際土壤微生物提供了適宜生長的場所與物質來源,植物向根際土壤分泌的碳水化合物越多,根際微生物對碳底物利用的能力便越強[22]。與騰應等在研究復墾紅壤中牧草根際微生物群落時的結果一致[23]。趙燕娜等人也發現土壤微生物數量與土壤酶活性均表現出根際聚集現象[24]。同時,除SCJ外,其他兩種植物根際土壤pH值和EC均小于非根際土,造成這種差異的原因可能是因為BLH與HMC在其根際能夠選擇性的富集一些鹽基離子,從而減少土壤中OH-[25],又由于植物根系對不同物質的吸收速率不同,使植物根際出現土壤礦物質鹽分離子的吸收或富集,并與非根際土壤表現出不同的鹽分含量[26]。從本研究中可以看出,博樂蒿與旱麥草能夠較好的適應鹽堿條件,并給土壤微生物提供較好生存環境,提高土壤肥力,改善土壤狀況。
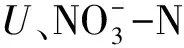
5 結論
(1)不同植物根際與非根際土微生物代謝強度不同,本研究中博樂蒿根際土具有較好的微生物活性;各植物根際土壤微生物對糖類、脂類、酸類和胺類碳源物質比較敏感,非根際土壤微生物敏感于酸類、氨基酸類以及糖類碳源物質,根際土壤微生物利用碳源寬度更大,且不同植物類型根際環境微生物碳源利用特征不同。研究表明,植被的存在能夠提高土壤微生物活性和功能多樣性,土壤狀況越好,越有利于適應環境變化。
參考文獻(References):
[1] Falkowski P G, Fenchel T, Delong E F. The microbial engines that drive Earth′s biogeochemical cycles. Science, 2008, 320(5879): 1034- 1039.
[2] 宋長青, 吳金水, 陸雅海, 沈其榮, 賀紀正, 黃巧云, 賈仲君, 冷疏影, 朱永官. 中國土壤微生物學研究10年回顧. 地球科學進展, 2013, 28(10): 1087- 1105.
[3] 羅希茜, 郝曉暉, 陳濤, 鄧嬋娟, 吳金水, 胡榮桂. 長期不同施肥對稻田土壤微生物群落功能多樣性的影響. 生態學報, 2009, 29(2): 740- 748.
[4] 林先貴, 胡君利. 土壤微生物多樣性的科學內涵及其生態服務功能. 土壤學報, 2008, 45(2): 892- 900.
[5] 杜瀅鑫, 謝寶明, 蔡洪生, 唐璐, 郭長虹. 大慶鹽堿地九種植物根際土壤微生物群落結構及功能多樣性. 生態學報, 2016, 36(3): 740- 747.
[6] 安韶山, 李國輝, 陳利頂. 寧南山區典型植物根際與非根際土壤微生物功能多樣性. 生態學報, 2011, 31(18): 5225- 5234.
[7] Nicolitch O, Colin Y, Turpault M P, Uroz S. Soil type determines the distribution of nutrient mobilizing bacterial communities in the rhizosphere of beech trees. Soil Biology and Biochemistry, 2016, 103: 429- 445.
[8] 馬琨, 楊桂麗, 馬玲, 汪春明, 魏常慧, 代曉華, 何文壽. 間作栽培對連作馬鈴薯根際土壤微生物群落的影響. 生態學報, 2016, 36(10): 2987- 2995.
[9] 馮偉, 管濤, 王曉宇, 朱云集, 郭天財. 沼液與化肥配施對冬小麥根際土壤微生物數量和酶活性的影響. 應用生態學報, 2011, 22(4): 1007- 1012.
[10] Wei J, Liu X Y, Zhang X Y, Chen X, Liu S, Chen L. Rhizosphere effect ofScirpustriqueteron soil microbial structure during phytoremediation of diesel-contaminated wetland. Environmental Technology, 2014, 35(4): 514- 520.
[11] Lal R, Mokma D, Lowery B. Relation between soil quality and erosion//Ratta R, Lal R, eds. Soil Quality and Soil Erosion. Washington DC: CRC Press, 1999: 237- 258.
[12] 周桔, 雷霆. 土壤微生物多樣性影響因素及研究方法的現狀與展望. 生物多樣性, 2007, 15(3): 306- 311.
[13] Riley D, Barber S A. Bocarbonate accumulation and pH changes at the soybean (Glycinemax(L.) Merr.) root-soil interface. Soil Science Society of America Journal, 1969, 33(6): 905- 908.
[14] Schutter M, Dick R. Shifts in substrate utilization potential and structure of soil microbial communities in response to carbon substrates. Soil Biology and Biochemistry, 2001, 33(11): 1481- 1491.
[15] 鮑士旦. 土壤農化分析. 北京: 中國農業出版社, 2000: 179- 183.
[16] Benizri E, Amiaud B. Relationship between plants and soil microbial communities in fertilized grasslands. Soil Biology and Biochemistry, 2005, 37(11): 2055- 2064.
[17] 曹永昌, 楊瑞, 劉帥, 王紫泉, 和文祥, 耿增超. 秦嶺典型林分夏秋兩季根際與非根際土壤微生物群落結構. 生態學報, 2017, 37(5): 1667- 1676.
[18] Fisk M C, Ruether K F, Yavitt J B. Microbial activity and functional composition among northern peatland ecosystems. Soil Biology and Biochemistry, 2003, 35(4): 591- 602.
[19] Kaiser O, Pühler A, Selbitschka W. Phylogenetic analysis of microbial diversity in the rhizoplane of oilseed rape (Brassica napus cv. Westar) employing cultivation-dependent and cultivation-independent approaches. Microbial Ecology, 2001, 42(2): 136- 149.
[20] Smalla K, Wieland G, Buchner A, Zock A, Parzy J, Kaiser S, Roskot N, Heuer H, Berg G. Bulk and rhizosphere soil bacterial communities studied by denaturing gradient gel electrophoresis: plant-dependent enrichment and seasonal shifts revealed. Applied and Environmental Microbiology, 2001, 67(10): 4742- 4751.
[21] Kowalchuk G A, Buma D S, de Boer W, Klinkhamer P G L, van Veen J A. Effects of above-ground plant species composition and diversity on the diversity of soil-borne microorganisms. Antonie van Leeuwenhoek, 2002, 81(1): 509- 520.
[22] 吳林坤, 林向民, 林文雄. 根系分泌物介導下植物-土壤-微生物互作關系研究進展與展望. 植物生態學報, 2014, 38(3): 298- 310.
[23] 滕應, 黃昌勇, 龍健, 姚槐應. 復墾紅壤中牧草根際微生物群落功能多樣性. 中國環境科學, 2003, 23(3): 295- 299.
[24] 趙燕娜, 廖超英, 李曉明. 毛烏素沙地4種固沙植物根際與非根際土壤生物學特性. 干旱區研究, 2015, 32(4): 680- 686.
[25] 王宏偉, 黃清俊, 李萍, 朱繼軍, 陳必勝. 3種草本植物鹽堿土栽培地的根際環境變化. 上海農業學報, 2012, 28(3): 66- 69.
[26] Kasuga M, Liu Q, Miura S, Yamaguchi-Shinozaki K, Shinozaki K. Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-inducible transcription factor. Nature Biotechnology, 1999, 17(3): 287- 291.
[27] 胡嬋娟, 傅伯杰, 劉國華, 靳甜甜, 劉宇. 黃土丘陵溝壑區典型人工林下土壤微生物功能多樣性. 生態學報, 2009, 29(2): 727- 733.
[28] Cartwright J, Dzantor E K, Momen B. Soil microbial community profiles and functional diversity in limestone cedar glades. Catena, 2016, 147: 216- 224.
[29] Torsvik V, ?vre?s L. Microbial diversity and function in soil: from genes to ecosystems. Current Opinion in Microbiology, 2002, 5(3): 240- 245.
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
中國科技論壇(2017年7期)2017-07-25 08:49:53
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
媽媽寶寶(2017年2期)2017-02-21 01:21:24
爆笑show(2016年7期)2017-02-09 09:36:13
國際漢語學報(2016年1期)2017-01-20 08:21:20
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
