橋山林區麻櫟群落主要喬木種群的種間聯結性
2018-06-22 11:29:32葉權平張文輝于世川薛文艷
生態學報 2018年9期
葉權平,張文輝,于世川,薛文艷
西北農林科技大學陜西省林業綜合重點實驗室, 楊凌 712100
種間聯結是指群落中不同物種在空間分布上的相互關聯性[1- 2],包括空間分布關系及物種之間的功能依賴關系[3- 5],描述的是物種關系外在現象,展現的是種間競爭的結果或群落的現狀[6]。它是群落數量和結構的重要特征之一,是群落形成、演化的基礎,也是種間相互關系的一種表現形式和群落分類的依據[7-8]。分析不同植物種個體間聯接程度,對研究種間相互作用和群落的組成與動態有重要意義,它不僅反映了現階段群落組成的結構特點,而且在很大程度上表現了群落未來的動態,演替趨勢和演替進程[9-10]。生態種組是群落生態習性相似的種的聯合,一個群落中表現出具有類似生態習性的相關物種組成的整體,反映群落和種群的關系以及種群對環境和主導生態因子的適應方式[11],對認識群落結構與功能具有重要意義。
麻櫟(Quercusacutissima)屬殼斗科櫟屬落葉喬木,為深根、喜光樹種,主要分布在暖溫帶和亞熱帶地區,因其耐干旱,萌芽能力強,在山區、丘陵均可栽植,麻櫟樹形高大、通直,生產潛力大,用途廣泛。屬橋山林區主要地帶性成林樹種之一,其對當地的水土保持、生物多樣性保護與生態區域安全發揮著重要作用[12]。目前,許多學者對不同植物群落的種間聯結和相關性進行了研究,如在三峽庫區栲屬群落的研究中表明群落在初級演替階段,種間關聯程度較低,有時產生較大的負關聯,競爭相對激烈[13]。對縉云山亞熱帶栲樹(Castanopsisfargesii)優勢種群研究顯示當群落演替到頂級階段時,群落的種類之間常常會達到某種協調,生態位彼此分化,表現出明顯的正關聯特性[14];對秦嶺華山松(Pinusarmandii)天然次生林群落研究顯示秦嶺山地華山松天然次生林群落喬木層、灌木層、草本層各優勢種對間種間關聯明顯,物種之間存在一定的獨立性,趨于形成一個整體較為穩定的群落,認為種對間正聯結比例越高,群落結構越趨于穩定[15];對滇西北云南紅豆杉(Taxusyunnanensis)群落研究表明聯結系數與生態位重疊值之間存在極顯著正相關[16]等。對麻櫟的研究也主要集中在碳密度及其空間分布格局[17],地上部分養分的積累和分配[18],葉片光合特征[19],環境及遺傳對麻櫟葉片N和P含量的影響[20]等。迄今為止,尚無研究報道麻櫟群落的種間聯結性。本研究以橋山林區麻櫟天然次生林為研究對象,通過方差比率法(VR),基于2×2聯列表的χ2檢驗、聯結系數(AC)、共同出現百分率(PC)和點相關系數Ф對橋山林區麻櫟群落主要喬木種群的種間聯結性進行測定分析,闡明現階段群落組成的結構特點以及未來群落的動態、演替趨勢和演替進程,為該區域麻櫟天然次生林的經營、更新、維持群落穩定提供依據;結合各種群的生態習性,對17個主要種群劃分生態種組,揭示各生態種組內物種間的相關性,為林區造林、林分改造提供借鑒。
1 研究區概況
研究區位于陜北黃土高原延安市橋山林業局雙龍林場,(108°31′—109°11′E,35°30′—35°50′N),海拔800—1700m,屬暖溫帶季風氣候,年平均氣溫9.4℃,最熱月(7月)平均氣溫21℃,極端最高氣溫38℃,最冷月(1月)平均氣溫-5℃,極端最低氣溫-23℃,無霜期190—225d,年均降水量677.4mm,多集中在7、8、9月,年日照時間為2528h,主要土壤類型為森林褐土和灰褐土。該區為天然次生林,主要喬木樹種有麻櫟、遼東櫟(Quercuswutaishanica)、油松(Pinustabuliformis)、茶條槭(Acerginnala)、槲櫟(Quercusaliena)、白樺(Betulaplatyphylla)等;主要灌木樹種有美麗胡枝子(Lespedezaformosa)、繡線菊(Spiraeasalicifolia)、刺五加(Acanthopanaxsenticosus)、水栒子(Cotoneastermultiflorus)、狼牙刺(Sophoraviciifolia)、胡頹子(Elaeagnuspungens)等;主要草本植物有披針葉苔草(Carexlanceolata)、白茅(Imperatacylindrica)、茜草(Rubiacordifolia)、地榆(Sanguisorbaofficinaliss)、野棉花(Anemonehupehensis)等[12]。
2 研究方法
2.1 樣地設置與數據統計
在橋山林區雙龍林場設置調查了22塊20m×30m的麻櫟群落典型樣地,計有喬木26種。調查記錄喬木(H≥1.3m)的種類、數量、高度、胸徑、冠幅,同時測定海拔、坡向、坡位等立地因子。統計每個樣地內出現的喬木樹種和樹種個體數,計算出頻度、密度以及種對出現的樣地數,計算樣地內物種的重要值表示喬木層物種優勢程度。選取重要值大于等于1.00(研究區內的優勢種、亞優勢種和伴生種)的主要種群計算和分析其種間聯結性,對于重要值小于1.00(在研究區屬于偶見種)的種群,不列入研究的主要種群范圍內。

(1)
2.2 總體聯結性檢驗
用D.Schluter提出的方差比率法(VR)確定整個群落的總體關聯性,計算公式為:
(2)
(3)
Pi=ni/N
(4)
(5)
式中,S為總物種數,N為總樣方數,Tj為樣方j內出現的目標物種總數,ni為物種i出現的樣方數,t為樣方中物種的平均數,VR為群落內植物間的總體關聯指數。
VR=1,則符合所有種間無關聯的零假設;VR>1,表示物種間總體上表現為正關聯;VR<1,表示物種間總體上表現為負關聯。
采用統計量W(W=VR×N)來檢驗VR值偏離1的顯著程度。若種間不顯著關聯,則W值落入χ2分布界限內的概率有90%。
(6)
2.3 種對間聯結性檢驗
2.3.1建立2×2聯列表1,進行種間關聯程度檢驗。
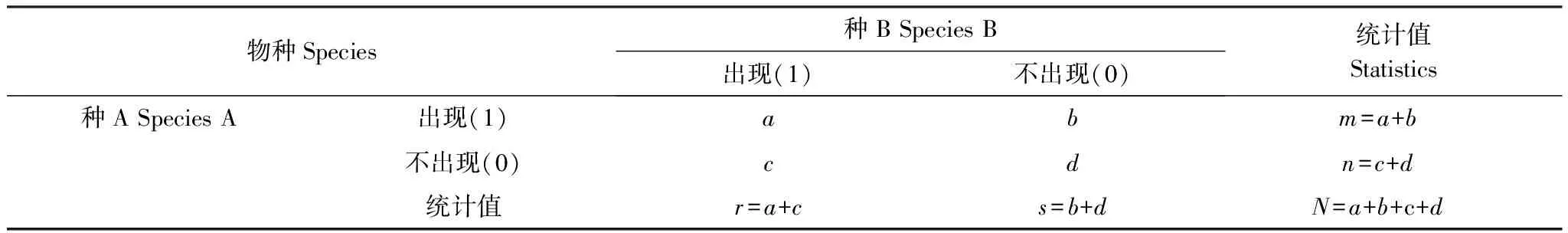
表1 種間聯結性測定2×2聯列表
2.3.2χ2檢驗
用Yates的連續性公式計算χ2值:
(7)
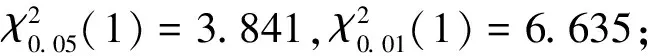
2.4 種間關聯度的測定
2.4.1 聯結系數AC
若ad≥bc,則:AC=(ad-bc) /[(a+b) (b+d)]
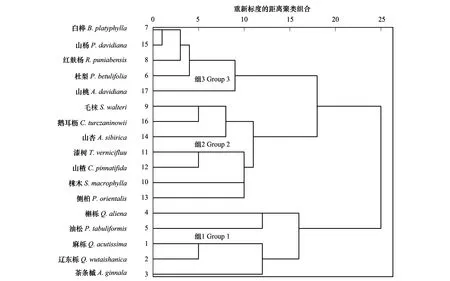
ad ad 聯結系數(AC)的值域為[-1,1],AC越接近于1,則種對間的正聯結性越強;反之AC值越接近于-1,則種對間的負聯結性越強;AC值為0,則種對間完全獨立。 2.4.2 共同出現百分率PC 為克服聯結系數(AC)受d值影響而造成偏差,本文再選用共同出現百分率PC測定種對聯結性程度: (8) 0≤PC≤1;a=0,PC=0,表明兩樹種不同時出現在同一樣方內,種對無關聯;a=N,PC=1,表明兩樹種同時出現在所有樣方內,種對關聯程度最緊密。 2.4.3 點相關系數Ф 為降低χ2檢驗、聯結系數(AC)和共同出現百分率(PC)對聯結性分析得影響,選用點相關系數Ф對聯結性進行檢驗測定: (9) Ф值域為[-1,1],Ф值越接近于1,則種對間的正聯結性越強;Ф值越接近于-1,則種對間的負聯結性越強。 統計分析橋山林區櫟屬群落26個喬木樹種的重要值(表2),選取重要值大于等于1.00的17個主要種群進行種間聯結性分析。 表2 喬木層物種的組成及重要值/% (10) 用統計量W來檢測VR值偏離1的顯著性:W=VR×N=15.40。 3.3.1χ2檢驗 圖1 χ2檢驗半矩陣圖Fig.1 Semi-matrix diagram of χ2 correlation test ▲:顯著正關聯 Significantly positive correlated; △:顯著負關聯 Significantly negative correlated; ■:正關聯 Positive correlated; □:負關聯 Negative correlation. 種序號同表1 17個主要種共136個種對的χ2統計量半矩陣見圖1。檢驗呈正關聯的有58對,占總對數的42.65%,其中顯著關聯的種對數為4個,不顯著相關的種對數為54個;負關聯的有78個,占總對數的57.35%,其中顯著關聯的種對數為1個,不顯著關聯的種對數為76個。根據χ2檢驗的基本原理分析得:所有種對中不顯著相關的有131個占96.32%,說明群落中喬木種群種對間關聯較弱,各物種間呈獨立分布格局,群落發育不成熟。4對顯著正關聯的種對為麻櫟與茶條槭、茶條槭與杜梨、白樺與毛梾、白樺和山楊,說明它們對生境和資源有相似的需求[21];顯著負關聯的種對為槲櫟與杜梨,說明它們對生境的需求不同呈現相互排斥性[22]。 3.3.2 聯結系數AC與共同出現百分率PC分析 由橋山林區麻櫟群落17個主要種聯結系數AC矩陣圖(表3)可得出,聯結系數AC≥0.5的種對數有3個占總對數2.21%,為毛梾與山楂、白樺與山楊、山楊與鵝耳櫪,說明各種對間正聯結程度較高;0.2≤AC<0.5的種對為18個,占總對數的13.24%,說明種對間呈不顯著正聯結性;-0.2≤AC<0.2的種對數分別為63,占總對數的46.32%,說明種對間聯結松散,分布趨于獨立;-0.5≤AC<-0.2的種對數有19對占總對數的13.97%,這表明種對間呈不顯著負相關;AC<-0.5的種對數有32個,其中AC= -1的種對數有25個占總對數的18.38%,相對于χ2檢驗值,負關聯的種對AC值要高一些,這與聯結系數AC夸大兩物種均不出現時種對間的聯結顯著程度有關。根據聯結系數AC測定種間關聯程度結果可知,麻櫟群落喬木層17個優勢種的種間有少部分種對間呈現顯著正相關或顯著負相關,但大部分種對間相關性不顯著,種間聯結較松散。 根據共同出現百分率PC測定原理,以PC>0.5表示種對間呈顯著-極顯著關聯,0 3.3.3 點相關系數Ф值分析 從點相關系數Ф矩陣圖(表5)可知,136個種對中,正聯結種對63個,其中Ф≥0.5即正聯結程度較強的種對6個,占總對數4.41%;負聯結種對73個,其中Ф<-0.5即負聯結程度較強的種對2個,占總對數的1.47%;-0.3≤Ф<0.3即聯結程度弱的種對數101個,占總對數的74.26%。 表3 聯結系數AC矩陣 表4 共同出現百分率PC矩陣 表5 點相關系數Ф矩陣 生態種組是群落生態習性相似的植物種的組合,根據樹種在所處的海拔,坡向、坡位等微地形條件內分布的不同[22],對麻櫟群落21個樣地內17個主要喬木樹種的物種分布狀況進行統計,將統計數據轉換為0,1形成的二元數據矩陣,進行系統聚類分析[23-24](圖2),數據統計分析及圖形輸出均使用SPSS 23.0軟件。物種的相對頻度(表2)體現物種在群落中水平結構上分布的均勻程度,及對不同生境條件的適應狀況。根據系統聚類分析結果,以相對頻度表示物種對環境適應性的相似程度,綜合相關系數檢驗結果,以環境適應性相似,同一生態種組的物種間多呈正聯結,不同生態種組的物種間多呈負聯結的物種分為一組為原則,劃分橋山林區麻櫟群落優勢種群的生態種組[6-7]。將17個主要種群分為3個不同的生態種組(表6)。 圖2 橋山林區麻櫟群落17個主要喬木種聚類分析Fig.2 Clustering analysis of 17 major tree species of Quercus acutissima community in the Qiaoshan forest area 生態種組Ⅰ中樹種:個體數量多,為喬木層的優勢種或亞優勢種,屬于群落演替后期的主導樹種,種對間多呈現正聯結,其中麻櫟為研究區主要地帶性成林樹種,群落的絕對優勢樹種。生態種組Ⅱ中樹種:分布較分散,對陽光的需求不高,具有一定的耐蔭性,樹種的獨立性較強,種對間聯結程度小。生態種組Ⅲ中樹種:分布較為集中,多喜光,生于陽坡,與群落中的優勢喬木樹種即生態種組Ⅰ樹種呈負聯結,其中白樺、山楊為群落中典型的演替初期先鋒樹種。 表6 3個生態種組樹種組成 種間聯結指標是對一定時期內群落組成物種之間相互關系的靜態描述,它隨群落演替階段不同而變化,在一定程度上反映物種間的相互關系及其與環境間的關系[25-26]。χ2檢驗方法正確判斷了麻櫟群落17個主要種群間聯結的顯著程度,種群聯結的顯著程度反映了群落的穩定性[27]。一般而言,群落正負關聯比值越大,群落越穩定[28],穩定性越強,群落越向頂極方向演替[29]。橋山林區麻櫟群落喬木層的種間聯結測定結果表明,17個主要種總體聯結性為不顯著負相關,大多數種對間聯結程度弱,趨于獨立分布。群落正負關聯比為0.81,表明群落結構不穩定,物種間不能穩定共存,群落尚不成熟,處于不穩定的演替階段。聯結系數AC和共同出現百分率PC能體現出那些由χ2檢驗判斷為不顯著種對的聯結性及大小,但共同出現百分率PC夸大了兩物種共同出現次數多時種對間的聯結顯著程度[28],如種對1—2、2—3和3—6等;聯結系數AC夸大兩物種均不出現時種對間的聯結顯著程度,如種對5—11、5—15和10—12等。而點相關系數Ф能降低上述幾種狀況對種群聯結性分析的影響,經過Ф值的測定比較,AC中種對5—12、5—16和8—9與PC中種對1—8、2—3和4—7的種間聯結顯著程度均有明顯降低。因此,種間聯結性分析以χ2檢驗為基礎,以AC、PC值為輔助參數,并經過Ф值的檢驗測定,進行綜合研究,可更科學準確地判定麻櫟群落種間聯結關系[28]。 總體上聯結系數AC、共同出現百分率PC及點相關系數Ф的測定分析結果都表明群落中喬木17個主要種群間關聯較弱,為不顯著負相關,這與χ2檢驗結果一致,但在顯著相關的水平上卻是正相關占優勢。以上結果說明,橋山林區麻櫟群落整體結構不穩定,主要喬木種群間聯結較松散,各物種占據適合自己的生態位,具有一定的獨立性;但群落正處于不斷完善階段,正向更穩定的方向演替,種內競爭和種間競爭一直存在,對生境需求、適應相同或相似的種群相伴出現幾率較大。其中白樺與山楊間呈正相關且所有測定結果都達到顯著相關,共同出現百分率PC值為0.79,達到極顯著相關,說明這兩個樹種對生境需求及生態習性高度相似。白樺和山楊都屬于更新能力強的喜光樹種,適應性較強,對土壤和水分要求不嚴,較耐干旱和瘠薄,是森林演替系列中的過渡階段,往往作為先鋒樹種形成塊狀相間白樺-山楊混交林。在荒山荒地、火災或砍伐破壞后的跡地上能迅速更新成林,形成優勢林分,但經過演替,長勢日漸衰弱,最終又被松、櫟主要成林樹種所更替。群落總體為不顯著負相關,但麻櫟作為群落內的絕對優勢種與群落內的大部分主要樹種呈現微弱正相關,與茶條槭之間有顯著的正相關。這說明麻櫟作為研究區主要地帶性成林樹種,其種群穩定性較好,現階段群落內麻櫟與其他樹種間對環境、資源需求存在競爭,但競爭還不強烈,為了適應環境,相互依賴性在增強。茶條槭耐庇蔭,喜溫暖濕潤環境,多與麻櫟混生,與麻櫟呈顯著正相關。 在群落中,聯結程度弱的種對占總對數比例大時,環境的干擾變化對群落的穩定性具有決定性作用。在本研究中,通過各聯結指標的測定可知橋山林區麻櫟群落中種間聯結程度弱,因此群落的穩定性較差,若在群落演替過程中,受到外界的干擾程度過大,群落的演替速度及方向都極易發生改變[29]。所以在對橋山林區麻櫟群落所在林分進行撫育間伐時,宜采用近自然經營方式,以撫育間伐促進群落正向發育與群落自行演化相結合,減弱人工撫育時對群落的干擾,遵循森林自然屬性,最大限度的保護林地物種多樣性,保障林分穩定持續更新。 種間聯結性反映物種在群落中的分布及對環境的利用與適應情況,因此種間聯結性分析結果可作為生態種組劃分的依據[30]。海拔、坡位、坡向等立地不同體現為林內溫度、濕度、光照等環境條件的不同,物種對不同立地條件的適應使其在群落內分布狀況不同,本研究中根據麻櫟群落17個主要喬木在不同立地條件下的分布進行聚類分析,綜合種間聯結性測定結果和生態習性,把橋山林區麻櫟群落17個主要種群劃分為3個生態種組,生態種組的組成反映研究區內物種與物種、物種與環境之間的關系[31],同一生態種組的樹種生態習性相似,樹種間種對多呈正聯結,說明它們對生境及資源有相似的需求。生態種組Ⅰ中樹種為麻櫟、遼東櫟、茶條槭、槲櫟和油松。在研究區橋山林區內麻櫟、遼東櫟等櫟類林及油松林是相對穩定的植物群落,也是該區群落演替后期的優勢樹種。麻櫟為群落絕對優勢種,槲櫟、栓皮櫟為亞優勢樹種,茶條槭為主要伴生樹種。生態種組Ⅲ中樹種為杜梨、白樺、紅麩楊、山桃、山楊,與生態種組Ⅰ樹種多呈負聯結。此生態種組中樹種多為先鋒樹,為先鋒喬木階段旱生矮生雜林樹種,如杜梨、山桃、白樺、山楊。他們于演替初期在荒山荒地、火災或砍伐破壞后的跡地上能迅速更新成林,形成優勢林分,但后期生長緩慢,壽命較短,在演替的過程中,最終被其它后期主要成林樹種如櫟類、油松等所更替。根據同一生態種組內樹種生態習性的相似性及不同生態種組間樹種的生態特性,可為研究區造林營林、植被恢復提供依據。如麻櫟與槲櫟、遼東櫟親緣關系相近,生態習性相似,種間競爭較為激烈,在經營管理中選取經營目標樹,以目標樹為中心進行林分撫育管理,適度疏伐干擾木,減緩種間競爭,促進目標林木生長;在造林樹種選擇時,于同一生態種組內選擇搭配樹種,利于林分穩定和發展,選擇生態種組Ⅰ內的后期成林樹種,能使群落跳過先鋒樹種被更替階段,更早達到穩定狀態;在對林分進行改造時,間伐生態種組Ⅲ內的山楊、白樺、山桃等陽性先鋒樹種樹種,補植、保護生態種組Ⅰ內麻櫟、遼東櫟、油松等后期成林樹種,促進群落演替進程,使群落更快達到穩定。 參考文獻(References): [1] Legendre P, Legendre LF. Numerical Ecology. 3rd Ed. Amsterdam: Elsevier, 2012. [2] Gleason H A. The individualistic concept of the plant association. Bulletin of the Torrey Botanical Club,1926, 53(1): 7- 26. [3] 陳玉凱, 楊小波, 李東海, 農壽千, 呂曉波, 呂潔杰, 楊民, 李小成. 海南霸王嶺海南油杉群落優勢種群的種間聯結性研究.植物科學學報, 2011, 29(3): 278- 287. [4] 彭少麟, 周厚誠, 郭少聰, 黃忠良. 鼎湖山地帶性植被種間聯結變化研究. 植物學報, 1999, 41(11): 1239- 1244. [5] 張志勇, 陶德定, 李德銖. 五針白皮松在群落演替過程中的種間聯結性分析. 生物多樣性, 2003, 11(2): 125- 131. [6] 徐滿厚, 劉敏, 翟大彤, 劉彤. 植物種間聯結研究內容與方法評述. 2016, (24): 24- 8233. [7] 郭志華, 卓正大, 陳潔, 吳梅鳳. 廬山常綠闊葉、落葉闊葉混交林喬木種群種間聯結性研究. 植物生態學報, 1997, 21 (5): 424 - 432. [8] 王伯蓀, 余世孝, 彭少麟. 植物群落學實驗手冊. 廣州: 廣東高等教育出版社, 1996. [9] 陽小成, 鐘章成. 綿陽官司河流域防護林優勢種群間的聯結性研究. 西南師范大學學報: 自然科學版, 1994, 19(3): 290- 297. [10] 韓銘哲. 興安落葉松自然更新格局和種群的生態對策. 內蒙古林學院學報, 1994, 16(1): 1- 10. [11] 丁文慧, 李秀珍, 姜俊彥, 黃星, 張運清, 張騫, 周云軒. 崇明東灘南部河口鹽沼植物群落種間關系的數量分析. 應用生態學報, 2016, 27(5): 1417- 1426. [12] 曹旭平, 張文輝. 陜西橋山省級自然保護區綜合科學考察報告. 楊凌: 西北農林科技大學出版社, 2015. [13] 程瑞梅, 王瑞麗, 劉澤彬, 封曉輝, 王曉榮, 肖文發. 三峽庫區栲屬群落主要喬木種群的種間聯結性. 林業科學, 2013, 49(5): 36- 42. [14] 杜道林, 劉玉成, 李睿. 縉云山亞熱帶栲樹林優勢種群間聯結性研究. 植物生態學報, 1995, 19(2): 149- 157. [15] 張明霞, 王得祥, 康冰, 張崗崗, 劉璞, 杜焰玲, 于飛. 秦嶺華山松天然次生林優勢種群的種間聯結性. 林業科學, 2015, 51(1): 12- 21. [16] 李帥鋒, 劉萬德, 蘇建榮, 郎學東, 張志鈞. 滇西北云南紅豆杉群落物種生態位與種間聯結. 植物科學學報, 2012, 30(6): 568- 576. [17] 成向榮, 虞木奎, 葛樂, 張春祥, 王婉. 不同間伐強度下麻櫟人工林碳密度及其空間分布. 應用生態學報, 2012, 23(5): 1175- 1180. [18] 唐羅忠, 劉志龍, 虞木奎, 方升佐, 趙 丹, 王子寅. 兩種立地條件下麻櫟人工林地上部分養分的積累和分配. 植物生態學報, 2010, 34(6): 661- 670. [19] 王標, 虞木奎, 孫海菁, 成向榮, 單奇華, 方炎明. 鹽脅迫對不同種源麻櫟葉片光合特征的影響. 應用生態學報, 2009, 20(8): 1817- 1824. [20] Zhang H, Guo W H, Wang G G, Yu M K, Wu T. Effect of environment and genetics on leaf N and P stoichiometry forQuercusacutissimaacross China. European Journal of Forest Research, 2016, 135(4): 795-802. [21] 黃世能, 李意德, 駱土壽, 王伯蓀. 海南島尖峰嶺次生熱帶山地雨林樹種間的聯結動態. 植物生態學報, 2000, 24(5): 569- 574. [22] 張金屯. 數量生態學. 北京: 科學出版社, 2004: 98- 106. [23] 王合玲, 呂光輝, 張輝國. 干旱區典型湖泊濕地主要植物生態種組分析. 生態環境學報. 2012,21(5): 858- 863. [24] 李意德, 許涵, 陳德祥, 駱土壽, 莫錦華, 羅文, 陳煥強, 蔣忠亮. 從植物種群間聯結性探討生態種組與功能群劃分—以尖峰嶺熱帶低地雨林喬木層數據為例. 林業科學, 2007, 43(4): 9- 16. [25] Ward J S, Parker G R, Ferrandino F J. Long-term spatial dynamics in an old-growth deciduous forest. Forest Ecology and Management, 1996, 83(3):189- 202. [26] 黃祥童,王紹先,黃炳軍,尹航,崔凱峰,趙偉,范宇光,顧德峰. 珍稀植物對開蕨與其伴生物種的聯結性及群落穩定性,生態學報,2015,35(1):0080-0090. [27] 王乃江, 張文輝, 陸元昌, 范少輝, 王勇. 陜西子午嶺森林植物群落種間聯結性. 生態學報, 2010, 30(1): 0067-0078. [28] 周先葉, 王伯蓀, 李鳴光, 昝啟杰. 廣東黑石頂自然保護區森林次生演替過程中群落的種間聯結性分析. 植物生態學報, 2000, 24 (3): 332- 339. [29] Greig-Smith P. Quantitative Plant Ecology. 3rd ed. Berkeley: University of California Press, 1983. [30] 王伯蓀, 彭少麟. 南亞熱帶常綠闊葉林種間聯結測定技術研究Ⅰ.種間聯結測式的探討與修正. 植物生態學與地植物學叢刊, 1985, 9(4): 274- 285. [31] 宋永昌. 植被生態學. 上海: 華東師范大學出版社, 2001.
3 結果與分析
3.1 優勢物種組成

3.2 總體相關性
3.3 種對聯間結性




3.4 生態種組的劃分

4 結論與討論
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18當代陜西(2021年17期)2021-11-06 03:21:36保健醫苑(2021年7期)2021-08-13 08:48:02學生天地(2020年36期)2020-06-09 03:12:30小學科學(學生版)(2020年5期)2020-05-25 07:11:32科學大眾(中學)(2019年3期)2019-05-17 10:04:30汽車觀察(2018年10期)2018-11-06 07:05:26學苑創造·A版(2018年11期)2018-02-01 06:29:20讀者(2017年5期)2017-02-15 18:04:18少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
