龍膽苦苷對人卵巢癌細胞株A2780/DDP順鉑化療敏感性的影響及機制探討*
2021-07-14 03:20:46井晶劉夢郭靜田二斌
中醫學報 2021年7期
井晶,劉夢,郭靜,田二斌
鄭州大學第二附屬醫院,河南 鄭州 450003
卵巢癌是一種高度異質類腫瘤,其發病率位于女性惡性腫瘤第2位,而致死率在女性惡性腫瘤中位居首位[1]。卵巢癌早期無明顯痛感和典型癥狀,不易察覺,因此確診時往往已為中晚期,癌細胞已轉移到腹腔,嚴重威脅女性健康[2]。化療技術是中晚期腫瘤治療的主要手段,以鉑類藥物為基礎的化療方法在卵巢癌的治療中普遍使用[3]。雖然該方法可提高患者5年生存率,但是由于卵巢癌復發率高、易耐藥,使得治療效果不明顯[4]。據報道,90%的卵巢癌晚期患者因多藥耐藥死亡[5]。研究表明,初次化療患者中至少存在20%的患者對鉑類化療藥物具有耐藥性,而在卵巢癌復發患者中,對該藥的耐藥率高達70%以上[6]。因此,如何降低腫瘤細胞對化療藥物的耐藥性是目前研究的熱點和難點。龍膽苦苷提取自龍膽科植物龍膽,具有抗炎鎮痛、保肝利膽、健胃等功效[7]。研究表明,龍膽苦苷還有抗腫瘤的作用,可抑制卵巢癌細胞增殖、凋亡和遷移[8]。但是該藥物對耐順鉑卵巢癌細胞順鉑化療敏感性的影響尚未涉及,因此,本研究使用龍膽苦苷處理人卵巢癌細胞株A2780/DDP,研究其對順鉑化療敏感性的影響,并探討其機制,為龍膽苦苷在臨床卵巢癌治療中的應用提供理論依據。
1 材料
1.1 細胞人卵巢癌細胞株A2780/DDP(上海美軒生物科技有限公司,貨號C1015112),-80℃恒溫保存,避免反復凍融。
1.2 藥物與試劑順鉑注射液(江蘇豪森藥業集團有限公司,批號:181204);龍膽苦苷(成都德思特生物技術有限公司,純度≥98%,批號:1808006)。RAMI 1640培養基(美國Gibco公司,批號:20181046);MTT法細胞增殖分析試劑盒(美國Trevigen公司,批號:1811102);吉姆薩染色試劑盒(上海歌凡生物科技有限公司,批號:20181016003);二甲基亞砜(蘇州聯雄化工科技有限公司,批號:20190516);4%多聚甲醛(上海遠慕生物科技有限公司,批號:201904112);RNA提取試劑盒(美國賽默飛公司,批號:1904125);蛋白提取試劑盒(武漢默沙克生物科技有限公司,批號:P1903147);反轉錄試劑盒(日本Takara公司,批號:190415);RT-qPCR染色試劑盒(常州百代生物科技有限公司,批號:1902183);Bcl-2、p53、XIAP、Survivin、p-p53、β-actin一抗和二抗(美國Aviva Systems Biology公司,批號:1904126、1904166、1901118、1904187、1902137、1906132、1905157、1907157、1906128、1908142、1905119、1907135);TUNEL凋亡檢測試劑盒(北京索萊寶公司,批號:20181203);BCA蛋白定量檢測試劑盒(美國Sigma公司,批號:20190812004);引物合成委托上海賽百盛基因技術有限公司。
1.3 儀器MCO-18AC CO2細胞培養箱(日本SANYO公司);ELX808酶標儀(美國伯騰儀器有限公司);Q1000 RT-qPCR儀(杭州朗基科學儀器有限公司);DY-1000C蛋白電泳設備(南京貝帝試驗儀器有限公司);CX31型光學顯微鏡(日本奧林巴斯公司);CKX53倒置顯微鏡(日本奧林巴斯公司);BD-YG500熒光顯微鏡(廣州博視達光學儀器有限公司)。
2 方法
2.1 細胞培養、分組和干預人卵巢癌細胞株A2780/DDP使用RAMI 1640培養基置于5%CO237℃恒溫細胞培養箱中培養,待細胞生長至對數期,每孔1×105個細胞加入96孔板中,分為對照組、順鉑組、龍膽苦苷組、順鉑+龍膽苦苷組,每組設置6個重復。順鉑組加入5 mg·L-1順鉑,龍膽苦苷組加入15μmol·L-1龍膽苦苷,順鉑+龍膽苦苷組加入15μmol·L-1龍膽苦苷和5 mg·L-1順鉑,對照組給予等體積生理鹽水,培養48 h。
2.2 細胞形態學觀察藥物干預24 h后,將細胞置于光學顯微鏡下觀察細胞形態。
2.3 細胞增殖抑制率檢測分別于藥物干預24 h和48 h后使用MTT法檢測細胞增殖抑制率。每孔加入20μL MTT溶液,置于細胞培養箱中培養4 h后,棄去培養基,每孔加入150μL二甲基亞砜,避光震蕩10 min,使用酶標儀讀取490 nm處光吸收值。
細胞增殖抑制率=(1-OD實驗組/OD對照組)×100%。
2.4 細胞凋亡指數檢測藥物干預48 h后,按照TUNEL試劑盒說明書檢測細胞凋亡指數。棄去細胞培養基,使用4%多聚甲醛固定細胞2 h,移去固定液,PBS洗3遍,按照TUNEL試劑盒說明書操作加入試劑,用抗熒光淬滅封片劑封片,在熒光顯微鏡下隨機觀察5個視野中凋亡陽性細胞個數,計算細胞凋亡指數。
細胞凋亡指數=(細胞凋亡數/細胞總數)×100%
2.5Bcl-2、p53、XIAP、Survivin的mRNA相對表達量檢測采用RT-qPCR檢測Bcl-2、p53、XIAP、Survivin的mRNA相對表達量。藥物干預48 h后,收集細胞至離心管中,加入Trizol試劑,按照RNA提取試劑盒說明書進行細胞總RNA提取,使用反轉錄試劑盒將RNA反轉錄為cDNA。從NCBI基因庫中查找目的基因mRNA序列并設計出目的基因上下游引物,引物序列見表1。反應條件為:94℃5 min,94℃1 min,56℃30 s,72℃40 s,最后55℃停止。通過配套熒光信號采集系統分析樣品CT值并以β-actin為內參計算各樣品mRNA相對表達量。

表1 引物序列
2.6 Bcl-2、p53、XIAP、Survivin蛋白表達水平及p53磷酸化水平檢測采用Western blot檢測Bcl-2、p53、XIAP、Survivin蛋白表達水平及p53的磷酸化水平。藥物干預48 h后,收集細胞于離心管中,通過蛋白提取試劑盒提取細胞總蛋白,BCA蛋白定量檢測試劑盒進行蛋白定量。取40μL樣品上樣,蛋白電泳,轉膜后加封閉液孵育,加一抗(1∶1 000)孵育,加二抗(1∶2 000)孵育,ECL化學發光試劑盒曝光、顯色,掃描成像,以β-actin圖像灰度值為對照,計算各組蛋白相對表達量。
2.7 統計學方法統計結果用均數±標準差(±s)表示,采用SPSS 22.0對實驗數據進行統計學分析。使用單因素方差分析對多組間數據進行比較,使用SNK-q檢驗對數據進行兩兩比較,使用t檢驗對組內數據進行比較。P<0.05表示差異具有統計學意義。
3 結果
3.1 龍膽苦苷對A2780/DDP細胞形態的影響對照組A2780/DDP細胞生長狀態良好,細胞排列呈現鋪路石狀,細胞連接緊密且輪廓清晰,偶見死亡懸浮細胞;順鉑組可見少量死亡懸浮細胞,細胞間隙變大,偶見細胞核碎裂;龍膽苦苷組可見部分死亡懸浮細胞,細胞核碎裂較多;順鉑+龍膽苦苷組可見較多細胞處于死亡懸浮狀態,細胞核碎裂嚴重。見圖1。
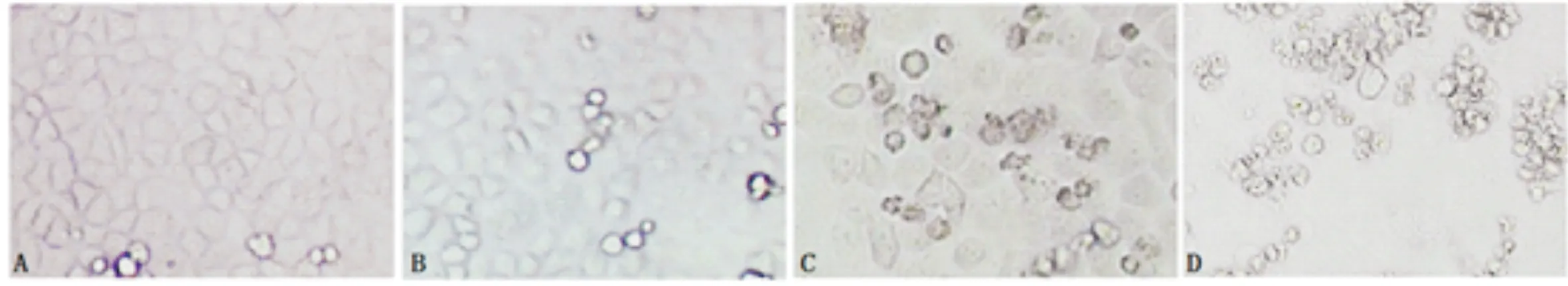
圖1 A2780/DDP細胞形態學觀察(×200)
3.2 龍膽苦苷對A2780/DDP細胞增殖抑制率的影響與對照組比較,藥物干預24h后,各給藥組A2780/DDP細胞增殖抑制率顯著升高(P<0.05);藥物干預48后,各給藥組A2780/DDP細胞增殖抑制率顯著升高,且高于24 h時同組的細胞增殖抑制率(P<0.05)。藥物干預24 h和48 h后,各組增殖抑制率兩兩比較均表現出:順鉑+龍膽苦苷組>龍膽苦苷組>順鉑組>對照組,且各組間比較均有顯著差異(P<0.05)。結果見表2。
表2 A2780/DDP細胞增殖抑制率比較 (±s,%)

表2 A2780/DDP細胞增殖抑制率比較 (±s,%)
注:與同組24 h比較,*P<0.05;與同期對照組比較,a P<0.05;與同期順鉑+龍膽苦苷組比較,b P<0.05;與同期龍膽苦苷組比較,c P<0.05
24 h 48 h對照組組別 n 6 0 0順鉑組 6 19.74±1.28abc 22.15±3.34*abc龍膽苦苷組 6 28.81±2.37ab 31.37±2.66*ab順鉑+龍膽苦苷組 6 34.12±3.84a 45.14±4.90*a
3.3 龍膽苦苷對A2780/DDP細胞凋亡指數的影響采用TUNEL法考察藥物干預48 h后,龍膽苦苷對A2780/DDP細胞凋亡指數的影響。與對照組比較,藥物干預48 h后,A2780/DDP細胞凋亡指數顯著升高(P<0.05),且順鉑+龍膽苦苷組>龍膽苦苷組>順鉑組>對照組,各組間比較均有顯著差異(P<0.05)。結果見圖2,表3。
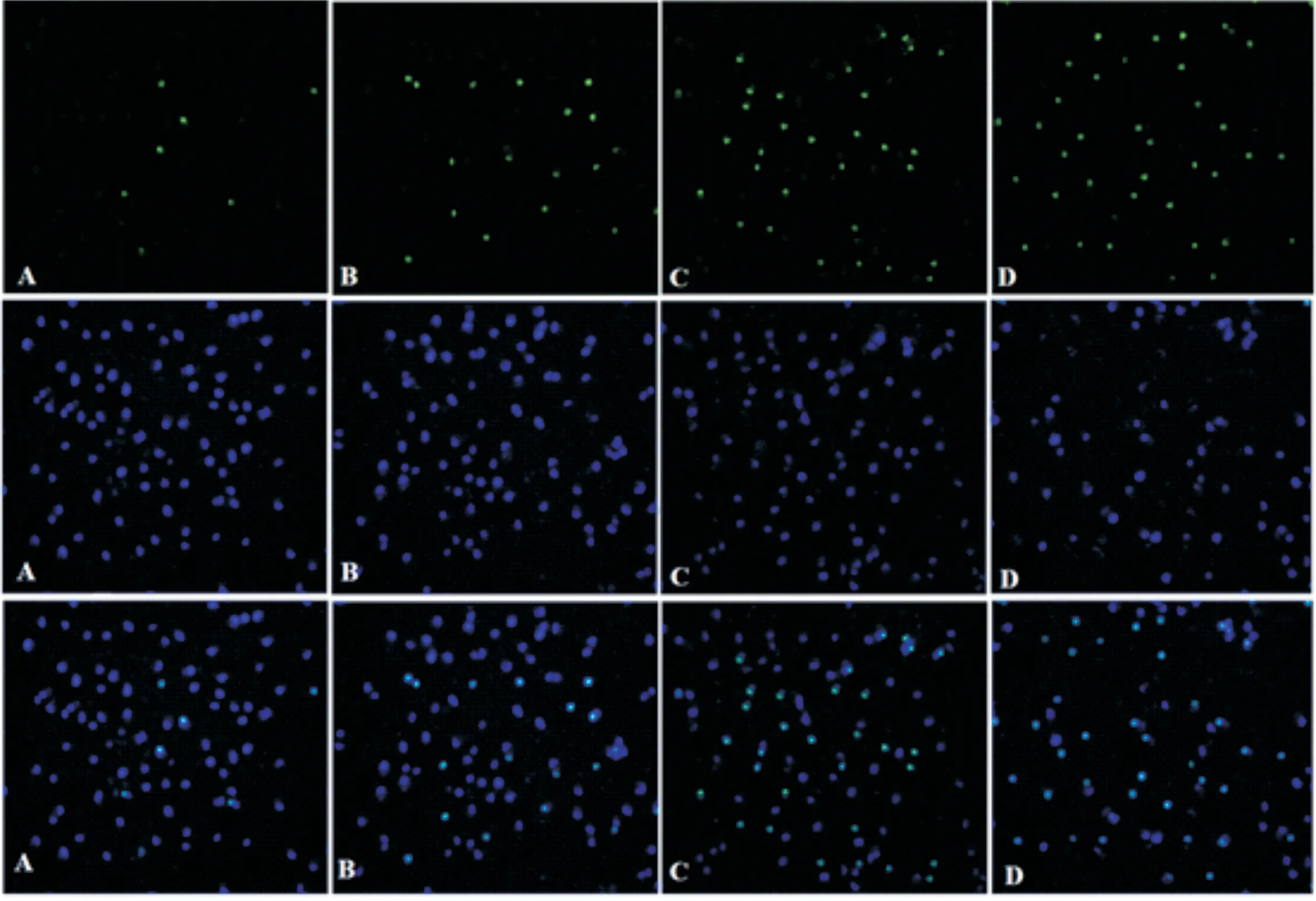
圖2 TUNEL檢測A2780/DDP細胞凋亡(×100)
表3 A2780/DDP細胞凋亡指數(±s,%)

表3 A2780/DDP細胞凋亡指數(±s,%)
注:與對照組比較,a P<0.05;與順鉑+龍膽苦苷組比較,b P<0.05;與龍膽苦苷組比較,c P<0.05
組別 n 細胞凋亡指數對照組5.06±0.52順鉑組 6 19.33±2.38abc龍膽苦苷組 6 36.40±5.31ab順鉑+龍膽苦苷組 6 65.27±10.55 6 a
3.4 龍膽苦苷對A2780/DDP細胞Bcl-2、p53、XIAP、Survivin的mRNA表達的影響與對照組比較,p53 mRNA表達水平顯著升高(P<0.05),且順鉑+龍膽苦苷組>龍膽苦苷組>順鉑組>對照組,各組間比較均有顯著差異(P<0.05);Bcl-2、XIAP和Survivin的mRNA表達水平顯著降低(P<0.05),且對照組>順鉑組>龍膽苦苷組>順鉑+龍膽苦苷組,各組間比較均有顯著差異(P<0.05)。結果見表4。
表4 A2780/DDP細胞中Bcl-2、p53、XIAP、Survivin的mRNA相對表達量 (±s)
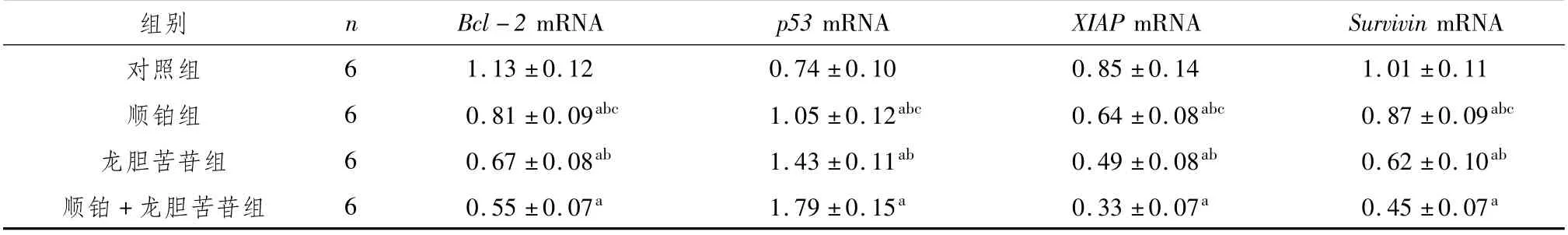
表4 A2780/DDP細胞中Bcl-2、p53、XIAP、Survivin的mRNA相對表達量 (±s)
注:與對照組比較,a P<0.05;與順鉑+龍膽苦苷組比較,b P<0.05;與龍膽苦苷組比較,c P<0.05
mRNA對照組組別 n Bcl-2 mRNA p53 mRNA XIAP mRNA Survivin 6 1.13±0.12 0.74±0.10 0.85±0.14 1.01±0.11順鉑組 6 0.81±0.09abc 1.05±0.12abc 0.64±0.08abc 0.87±0.09abc龍膽苦苷組 6 0.67±0.08ab 1.43±0.11ab 0.49±0.08ab 0.62±0.10ab順鉑+龍膽苦苷組 6 0.55±0.07a 1.79±0.15a 0.33±0.07a 0.45±0.07 a
3.5 龍膽苦苷對A2780/DDP細胞Bcl-2、p53、XIAP、Survivin蛋白表達水平及p53磷酸化水平的影響與對照組比較,p53蛋白表達量和磷酸化水平顯著升高(P<0.05),且順鉑+龍膽苦苷組>龍膽苦苷組>順鉑組>對照組,各組間比較均有顯著差異(P<0.05);Bcl-2、XIAP和Survivin蛋白表達量顯著降低(P<0.05),且對照組>順鉑組>龍膽苦苷組>順鉑+龍膽苦苷組,各組間比較均有顯著差異(P<0.05)。見圖3,圖4。

圖3 A2780/DDP細胞中Bcl-2、p53、XIAP、Survivin蛋白表達水平及p53磷酸化水平

圖4 A2780/DDP細胞中Bcl-2、p53、XIAP、Survivin蛋白相對表達水平及p53磷酸化水平
4 討論
隨著我國老齡化日益嚴重,卵巢癌的發病率也逐漸提高[9]。卵巢位于盆腔內,位置隱蔽,發病早期無明顯癥狀,又尚無早期特異性診斷標志物,因此早期難以診斷[10]。未生育、高脂飲食等都是誘發該病的危險因素[11]。目前,化療是中晚期卵巢癌患者治療的重要手段,以順鉑為主的第一代化療藥物具有廣譜抗癌、殺傷力強的特點,廣泛運用于惡性腫瘤的臨床治療[12]。順鉑進入細胞,通過與細胞核中DNA結合從而抑制其轉錄和復制過程,最終誘導細胞凋亡[13]。但是,長期使用順鉑會使腫瘤細胞產生耐藥性,藥效大打折扣。卵巢癌細胞對順鉑的耐藥機制具有多種途徑,阻礙順鉑與DNA結合、干擾細胞凋亡等都是其產生耐藥性的可能機制[14]。因此,尋找加強耐藥細胞對順鉑敏感性的方法,是解決腫瘤細胞耐藥問題的關鍵所在。
龍膽為我國傳統中草藥,具有瀉肝膽火、除下焦濕熱的功效,龍膽苦苷為其主要成分,除保肝利膽,還有抗菌、消炎、止痛、抗腫瘤等功效[15]。研究表明,龍膽苦苷可誘導人肝癌細胞、卵巢癌細胞凋亡[16-17]。本研究使用龍膽苦苷和順鉑作用于對順鉑耐藥的A2780/DDP細胞,24 h后細胞出現了不同程度的形態學改變,其中龍膽苦苷和順鉑聯合用藥組A2780/DDP細胞形態改變最嚴重。細胞增殖抑制率實驗證明,龍膽苦苷可抑制A2780/DDP細胞增殖,并且順鉑+龍膽苦苷組A2780/DDP細胞增殖抑制效果最佳。TUNEL實驗結果再次證實,龍膽苦苷可顯著促進A2780/DDP細胞凋亡,其中順鉑+龍膽苦苷組細胞凋亡指數顯著高于其余各組,表明龍膽苦苷可抑制A2780/DDP細胞增殖,促進細胞凋亡,增強耐藥細胞株A2780/DDP的順鉑敏感性。
順鉑可誘導腫瘤細胞凋亡,但其作用機制尚未完全明確。目前,已知p53、Bcl-2蛋白在順鉑誘導的細胞凋亡中起重要作用[18]。p53為腫瘤抑制蛋白,研究表明,p53基因的激活可誘導腫瘤細胞的凋亡,也是順鉑誘導細胞凋亡的關鍵所在,p53磷酸化水平與其穩定、激活有直接關系[19]。順鉑可激活p53,誘導p53磷酸化,從而發揮其誘導細胞凋亡的功能,而p53基因突變可使卵巢癌細胞株對順鉑的敏感性下降,誘導順鉑耐藥[20],因此,激活p53是提高順鉑化療敏感性的重要環節。Bcl-2家族蛋白位于線粒體中,對細胞凋亡具有調控作用,其中Bcl-2具有抑制凋亡作用,Bcl-2的過表達可導致細胞存活延長,凋亡抑制[21]。研究表明,順鉑耐藥腫瘤細胞中可觀察到Bcl-2的過表達,而降低Bcl-2表達水平可提高順鉑化療敏感性[22]。XIAP和Survivin屬于凋亡抑制蛋白,對細胞周期、信號傳導、細胞凋亡均有調節作用。凋亡抑制蛋白可以通過阻礙細胞凋亡,促進腫瘤細胞的耐藥性[23]。XIAP的過表達可導致存活因子如caspase-3、caspase-7、caspase-9等蛋白磷酸化并失活,從而阻斷順鉑誘導細胞凋亡。研究表明,順鉑可以抑制敏感細胞株中XIAP蛋白的表達并誘導caspase-3和caspase-9活化,引起細胞凋亡,而耐藥細胞株中XIAP蛋白高表達則細胞對順鉑化療敏感性降低[24],因此,抑制XIAP基因和蛋白表達可增加癌細胞對順鉑化療的敏感性。Survivin在惡性腫瘤中存在過表達,而正常細胞中表達量極低。研究表明,向人卵巢癌細胞中轉染Survivin基因,誘導Survivin表達量升高,可導致其順鉑耐藥性顯著增加,化療敏感性顯著降低[25]。本 研 究 對A2780/DDP細 胞 株 中p53、Bcl-2、XIAP、Survivin的表達量進行檢測,結果顯示,p53 mRNA和蛋白及其磷酸化水平表達增加而Bcl-2、XIAP、Survivin的mRNA和蛋白表達量均降低,表明龍膽苦苷可通過上調p53表達水平和磷酸化水平,下調Bcl-2、XIAP和Survivin表達水平,增強A2780/DDP細胞株順鉑敏感性,促進A2780/DDP細胞凋亡。
綜上所述,龍膽苦苷可抑制細胞株A2780/DDP增殖、促進腫瘤細胞變形、凋亡,且可增強耐順鉑A2780/DDP細胞對順鉑的敏感性,推測其機制是通過上調p53表達水平及磷酸化水平,下調Bcl-2、XIAP和Survivin表達水平。本研究結果對克服卵巢癌順鉑耐藥性具有臨床意義,但是龍膽苦苷對正常細胞是否具有細胞毒性尚未討論,其臨床應用可行性也尚未涉及,因此有待后續深入研究。
