
梨自交花粉原位萌發觀察及不親和性強度研究
2018-07-18 02:19:04金子明王國明柯亞琪石蘇利
江蘇農業科學 2018年12期
關鍵詞:生長
金子明,王國明,柯亞琪,石蘇利,吳 磊,谷 超
(南京農業大學梨工程技術研究中心,江蘇南京 210095)
薔薇科梨屬(PyrusL.)植物普遍存在由S基因位點控制的配子體型自交不親和(gametophytic self-incompatibility)現象,大多數品種表現為自花授粉不親和以及低結實率的特征[1]。大量研究表明,梨自花授粉不親和性主要是由花柱分泌的S糖蛋白引起的,這種S糖蛋白能夠分解自花授粉花粉管的RNA,導致花粉管停止生長并進入程序性死亡[2-3]。也有人認為該反應由S位點(S-locus)的1對S等位基因控制,即由雌蕊和花粉的S基因控制,分別為S-RNase、S-locusF-box/S-haplotype-specificF-box基因[4]。當花粉的S基因與雌蕊的1對S等位基因之一的S基因相同時,自花花粉能夠在柱頭上萌發,但花粉管沿花柱向子房伸長過程中會被花柱內的S糖蛋白抑制,而不能完成受精坐果[5],這種普遍存在的機制能夠抑制品種自交,促進異花授粉。在各種自然因素的作用下,自交不親和植物也會發生自交親和性突變,一種是花柱突變,如日本梨品種奧嗄二十世紀是二十世紀(Nijisseiki,S基因型S2S4)的突變體,其中1個花柱S基因,即S42RNase基因發生變異,導致奧嗄二十世紀的花柱不能識別自己的花粉[6]。另一種自交親和性突變就是花粉發生突變,即花粉失去自交不親和(self-incompatibility,簡稱SI)識別功能,而形成花粉SI突變體[5]。如吳華清等研究的金墜梨是鴨梨的芽變品種,金墜梨自花授粉結實率高達76%,自花的花粉管能正常生長至花柱基部,由于花粉S基因發生突變,導致自交不親和性功能的喪失,表現出自花授粉能夠結實[7]。梨屬于配子體型自交不親和果樹,絕大多數品種自花授粉結實率極低,在生產上必須配置授粉樹以及花期人工輔助授粉,才能完成授粉受精,獲得經濟產量,但是同時也增加了生產成本[4]。盡管如此,也常因授粉樹配置不合理、人工授粉不及時或是花期不良的氣候條件等問題影響授粉受精而造成減產[5],而選育自花結實性品種是解決這些問題的重要途徑之一。加深對梨自交不親和性的研究,不僅有助于更好地了解這一生命現象,還可以為果樹育種和生產提供可靠的理論依據。同時為進一步闡明梨自交親和性突變機制,深入研究自花花粉管被抑制的機制,從形態學角度探明花粉管在花柱中的生長特性,以及不同品種自交不親和強度分析統計,為顯花植物防止自交衰退這一遺傳機制提供一些試驗統計數據。因此,對不同品種的梨樹自交不親和性強度的研究,不僅具有理論價值,而且有潛在的實踐意義。
1 材料與方法
1.1 材料準備
本試驗于2014年3月至2016年6月,分別于江蘇省農業科學院梨資源圃和南京農業大學江浦農場園藝試驗站梨資源圃內進行,共有256個品種進行套袋處理。每個品種隨機選取6朵發育正常的并且處于大蕾期的花朵進行硫酸紙套袋處理。待花開后花藥散粉時,輕輕搖動紙袋或樹枝,使其自交授粉。為了確保每朵花自交授粉成功,從套袋后第2天起,每天搖動紙袋或樹枝2次。5 d后采取套袋花朵,從基部截取花柱并迅速置于FAA(甲醛 、冰乙酸、70%乙醇體積比 5 ∶5 ∶90)固定液中,4 ℃保存。
1.2 花粉管原位熒光顯微觀察并拍照
花粉原位萌發的熒光顯微觀察技術參照吳巨友等的方法[8],并稍有改動:從FAA固定液中取出樣品,用蒸餾水多次清洗后,放入2 mol/L NaOH溶液中軟化,65 ℃水浴40 min,使組織軟化;再次清洗多次后,用0.1%苯胺藍水溶液(用0.1% K3PO4溶液配制)避光室溫染色4~6 h。多次清洗后,滴甘油壓片,用宏觀變倍體式熒光顯微鏡(型號:MVX10,日本OLYMPUS公司)觀察并拍照。
1.3 數據處理
利用Image-ProPlus6.0軟件測量花柱中花粉管的生長長度及其花柱的長度,利用Excel 2007統計軟件計算不同品種花粉管伸長長度占花柱全長的比值,并進行統計分析。
1.4 坐果率調查
于2016年選取篩選后的梨品種進行自花授粉坐果率調查。在開花前1周,對發育一致的花序進行疏花,只留 2~3個生長較好的花蕾,進行套袋處理。盛花期輕輕搖動套袋的花序,進行自花授粉,每天搖動紙袋或樹枝2次。分別在盛花后20、30 d統計坐果率,坐果率大于20%被認為是自交親和[9]。
2 結果與分析
2.1 花粉管生長特性觀察
對梨不同品種花粉管原位熒光拍照觀察發現,花柱中花粉管分布及數量表現為上部較多、中部較少、下部最少的梯度分布(圖1-A、圖1-B)。部分自花授粉的花粉管停止生長時會出現少量的尖端膨大和彎曲等變形現象,與陳迪新等研究觀察結果一致[10](圖1-C、圖1-D)。同時發現自交不親和性較強的品種花粉管雖然有少量穿過柱頭,但不能進一步在花柱內生長,表現為花粉管在柱頭上為畸形狀、扭曲變形、先端膨大、雜亂無章等不親和性現象(圖1-E、圖1-F)。

2.2 梨自交不親和性強度
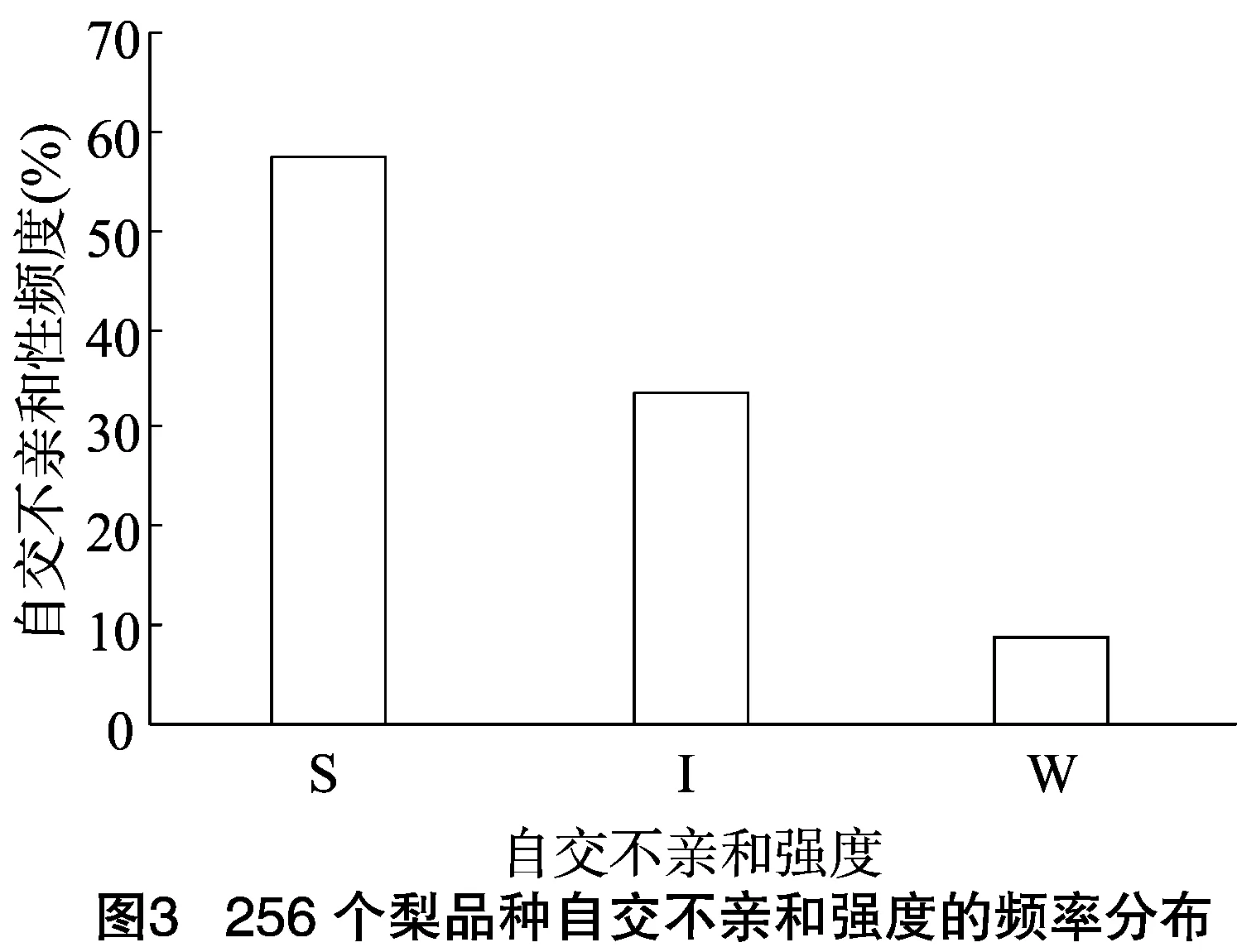
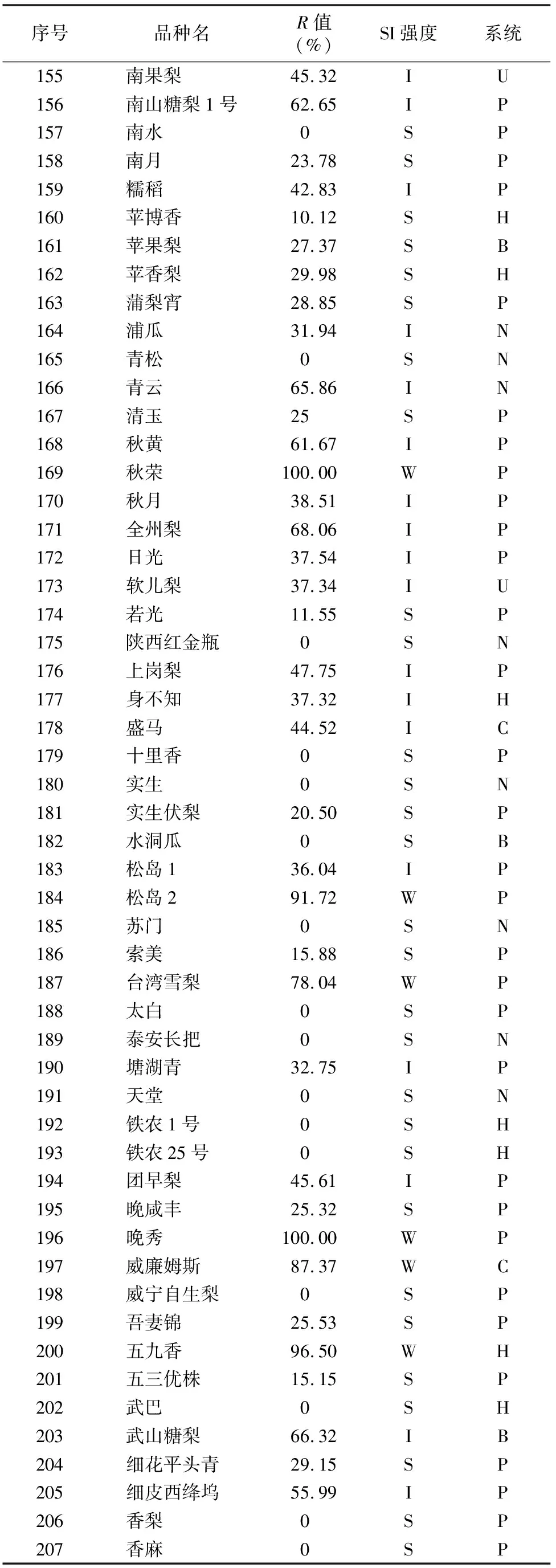
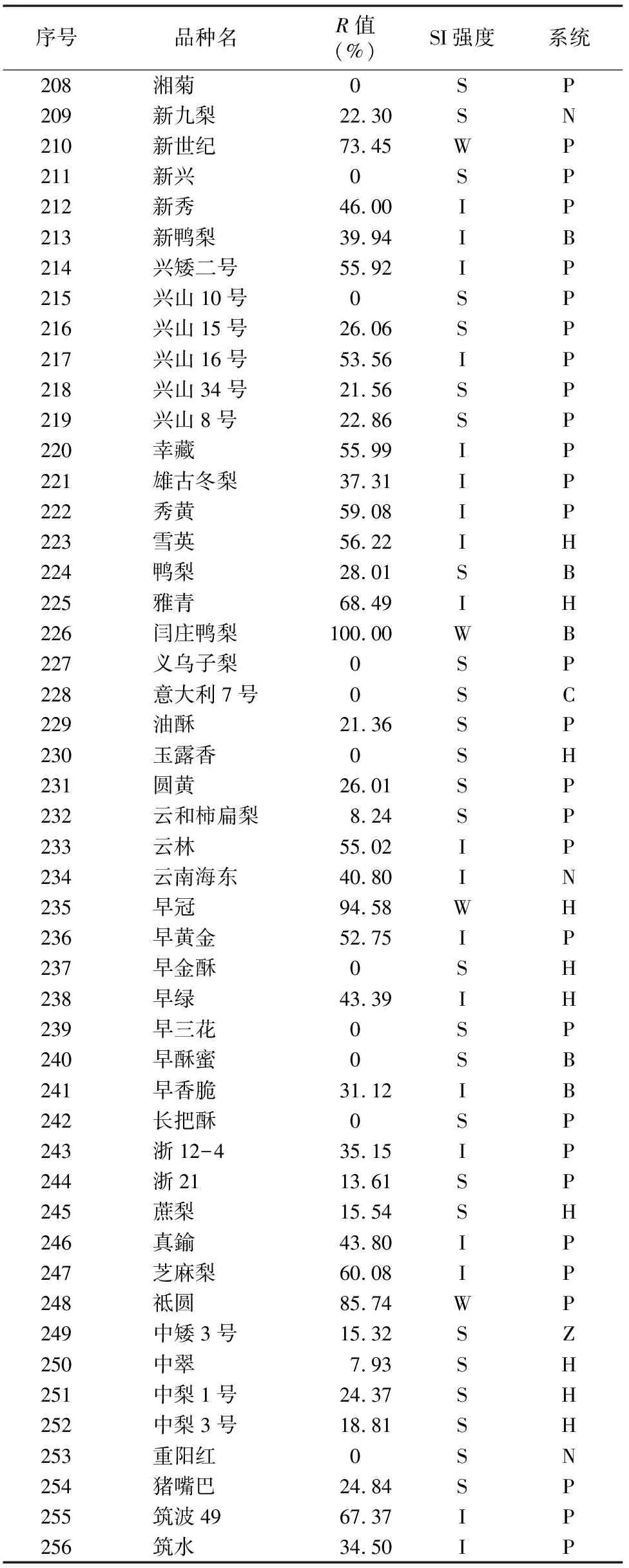
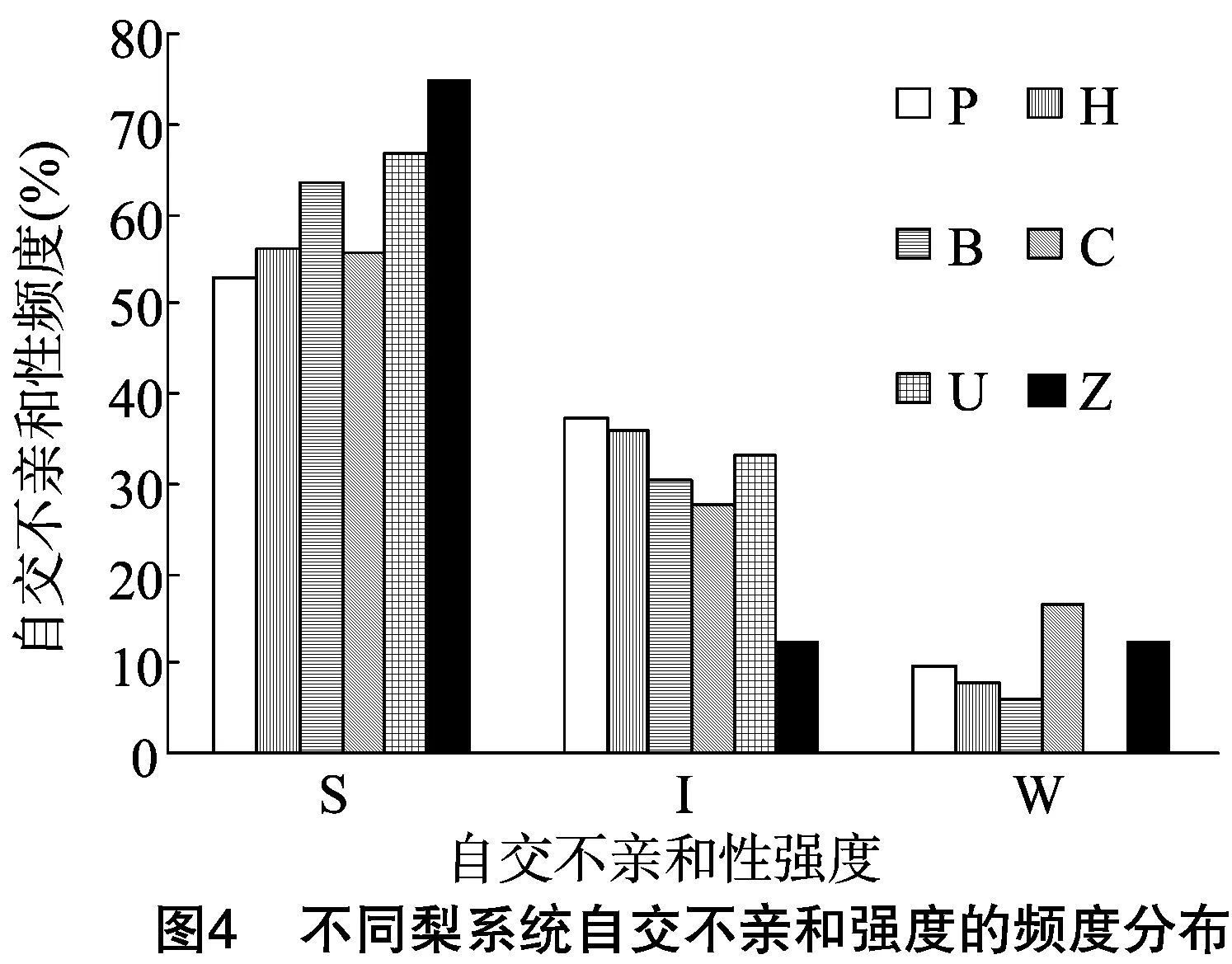
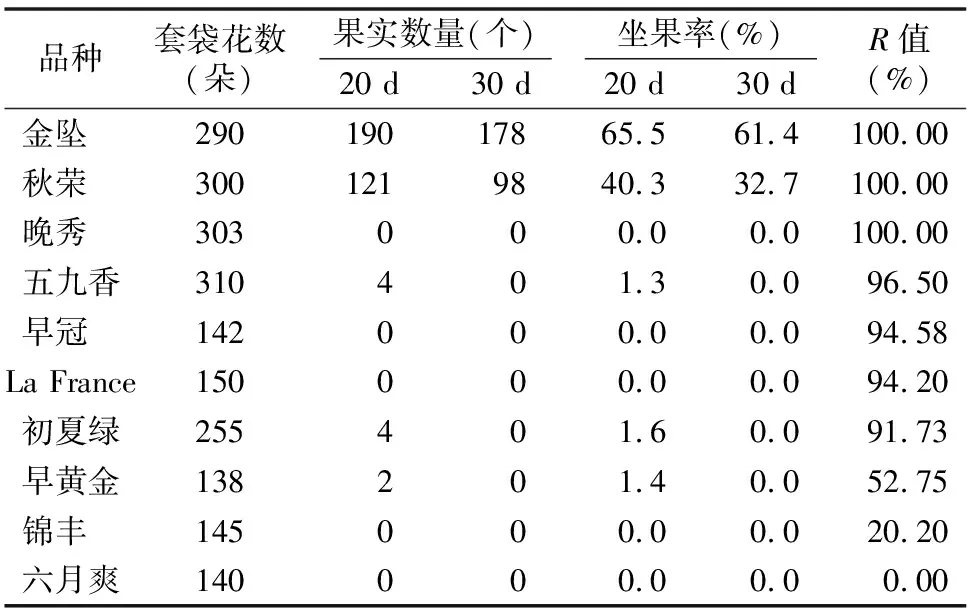
由于不同果樹樹種自交親和與不親和性的判定標準不同,且對于同一樹種不同研究者掌握的尺度也不同。張紹鈴等通過S糖蛋白含量高低來判斷自交不親和強度[11]。還有研究表明,自花授粉的花粉管在花柱中生長長度與其S糖蛋白含量顯著相關[12]。本研究根據自交授粉花柱中花粉管生長長度占整個花柱的比例定義品種的自交不親和強度R值,通過對R值進行分類,將梨供試品種的自交不親和強度分為強(強,0≤R≤30%)、中(中,30% 由于梨自交不親和的花粉能在柱頭上萌發并進入花柱,但在花柱中的生長受到抑制,不能完成受精,本試驗對套袋處理的256個梨品種從基部截取花柱,染色拍照并進行統計分析。結果顯示,不同梨品種自交不親和性強度差異明顯,不親和性強度變幅為0~100%,自交不親和性強度R值強、中、弱參差不齊。花粉管可以完全長到基部,共發現6個極可能為自交親和性的品種,只占調查總數的2.34%。本研究發現大多數品種花粉管不能長到基部(表1),符合梨屬于自交不親樹種的基本屬性,說明梨屬于典型的自交不親和性植物。這6個不同梨品種的R值為100%,其中閆莊鴨梨[13]、秋榮[14]、54S-135[14]、金墜梨[7]、大果黃花梨[15]是已報道的自交親和性品種,而晚秀則是本試驗通過自花花粉管發現的極可能為自交親和性品種。這些品種的花粉管能在自交花柱中生長并長至花柱底部,至于能否完成下一步受精及坐果過程,需對這些品種進行進一步自花套袋座果率調查。通過鑒定自交不親和強度R值,對后期自交親和性品種進行了初步篩選。并且為后期探索花粉管生長長度與自交不親和強度的深入研究提供了充足的基礎數據。 對256個品種分析發現,其中自交不親和性強度為強的有147個,占所調查總數的57.4%;自交不親和性強度為中的有86個,占33.6%;自交不親和性強度為弱的有23個,占9.0%。從頻度分布角度分析,梨不同品種自交不親和強度主要趨于強與中,花粉管能長到花柱基部的占少數,進一步驗證了梨屬于配子體自交不親和植物(圖3)。對256個品種所屬系統歸類和對不同系統自交不親和性強度的分布頻度進行統計比較,發現每個系統自交不親和性強度R值的強、中和弱分布符合所調查的整體規律,自交不親和性強的占大多數,而自交不親和性弱的僅占少數(表1、圖4)。同時對比發現,西洋梨與砂梨系統自交不親和性強度R值弱的分布頻度較其他系統高(圖4),為發現更多的自交親和性品種提供了方向。 表1 不同梨品種自交花柱中花粉管的生長長度占花柱的比例(R值)、自交不親和性強度分類及所屬系統 續表1 序號品種名R值(%)SI強度系統49蒼梧大沙梨0SP50崇化大梨0SB51初夏綠91.73WP52粗柄酥14.81SP53粗皮西絳塢41.79IP54脆綠7.10SH55大慈梨0SB56大果黃花100.00WP57大果酥梨0SB58大菊水54.99IP59大青67.10IN60大葉雪0SP61碭山酥梨11.90SB62冬果45.30IB63冬蜜19.58SH64凍花蓋0SU65豆梨13.51SZ66鵝梨61.26IB67鵝酥0SB68鄂梨1號69.83IH69鄂梨2號0SH70恩梨24.35SB71二宮白46.81IP72法蘭西0SC73樊山0SB74豐水A0SP75豐月0SN76甘川37.50IP77甘谷香水64.48IU78甘梨早60SH79甘梨早80SH80桂冠21.78SP81桂花梨76.09WP82漢源大白38.35IB83杭紅66.29IP84杭青7.82SP85紅安久0SC86紅巴梨B40.44IC87紅蓓蕾砂9.43SC88紅哈代0SC89紅考蜜斯9.34SC90紅茄梨10.96SC91紅山0SN92紅香26.96SP93紅香蜜0SB94紅香酥0SB95紅霄-10SB96紅星0SC97紅云2號0SP98花蓋22.34SU99花長把57.83IX100華金35.73IH101華酥34.87IP 續表1 序號品種名R值(%)SI強度系統102華王0SP103黃金梨0SP104黃皮鄂梨0SP105黃茄梨60.26IP106黃色品種西洋梨86.06WC107黃縣長把30.61IB108黃香37.36IB109會理早白0SP110金棒頭32.02IB111金川雪梨0SB112金川野生梨2號40.17IP113金二十世紀0SP114金花梨0SB115金水酥47.22IH116金星0SB117金墜100.00WB118錦豐20.20SB119錦香梨0SH120晉大50.98IN121京白梨10.87SU122酒泉長把0SB123菊水10.02SP124君塚早生(褐皮)0SP125君塚早生(綠皮)0SP126康德0SP127考蜜斯18.60SC128孔德梨43.08IC129庫實0SN130蘭州軟兒梨21.08SU131蘭州長把33.34IX132梨園麻子梨53.06IP133禮縣新八盤43.16IB134麗江面梨0SP135遼陽大香0SU136臨夏尕吊蛋0SX137臨夏黃麻66.46IX138六月黃棕梨0SP139六月爽0SN140六月酥20.00SB141龍泉190SH142龍泉44號51.90IP143龍團梨19.42SP144隆回巨梨32.92IP145綠云28.25SP146麻梨23.35SU147滿豐52.43IP148滿園香0SU149茅盧庫介0SN150美人酥10.22SP151彌渡火把43.30IP152米黃23.26SN153面包梨0SN154明珠0SN 續表1 序號品種名R值(%)SI強度系統155南果梨45.32IU156南山糖梨1號62.65IP157南水0SP158南月23.78SP159糯稻42.83IP160蘋博香10.12SH161蘋果梨27.37SB162蘋香梨29.98SH163蒲梨宵28.85SP164浦瓜31.94IN165青松0SN166青云65.86IN167清玉25SP168秋黃61.67IP169秋榮100.00WP170秋月38.51IP171全州梨68.06IP172日光37.54IP173軟兒梨37.34IU174若光11.55SP175陜西紅金瓶0SN176上崗梨47.75IP177身不知37.32IH178盛馬44.52IC179十里香0SP180實生0SN181實生伏梨20.50SP182水洞瓜0SB183松島136.04IP184松島291.72WP185蘇門0SN186索美15.88SP187臺灣雪梨78.04WP188太白0SP189泰安長把0SN190塘湖青32.75IP191天堂0SN192鐵農1號0SH193鐵農25號0SH194團早梨45.61IP195晚咸豐25.32SP196晚秀100.00WP197威廉姆斯87.37WC198威寧自生梨0SP199吾妻錦25.53SP200五九香96.50WH201五三優株15.15SP202武巴0SH203武山糖梨66.32IB204細花平頭青29.15SP205細皮西絳塢55.99IP206香梨0SP207香麻0SP 續表1 序號品種名R值(%)SI強度系統208湘菊0SP209新九梨22.30SN210新世紀73.45WP211新興0SP212新秀46.00IP213新鴨梨39.94IB214興矮二號55.92IP215興山10號0SP216興山15號26.06SP217興山16號53.56IP218興山34號21.56SP219興山8號22.86SP220幸藏55.99IP221雄古冬梨37.31IP222秀黃59.08IP223雪英56.22IH224鴨梨28.01SB225雅青68.49IH226閆莊鴨梨100.00WB227義烏子梨0SP228意大利7號0SC229油酥21.36SP230玉露香0SH231圓黃26.01SP232云和柿扁梨8.24SP233云林55.02IP234云南海東40.80IN235早冠94.58WH236早黃金52.75IP237早金酥0SH238早綠43.39IH239早三花0SP240早酥蜜0SB241早香脆31.12IB242長把酥0SP243浙12-435.15IP244浙2113.61SP245蔗梨15.54SH246真鍮43.80IP247芝麻梨60.08IP248祗圓85.74WP249中矮3號15.32SZ250中翠7.93SH251中梨1號24.37SH252中梨3號18.81SH253重陽紅0SN254豬嘴巴24.84SP255筑波4967.37IP256筑水34.50IP 注:S(strong)代表自交不親和強度強的品種,I(intermediate)代表中強度的品種,W(weak)代表弱強度的品種,B代表白梨(P.brestschneideri),C代表西洋梨(P.communis),H代表種間雜交(interspecific hybridization),N(unknow species)代表未知種類,P代表砂梨(P.pyrifolia),U代表秋子梨(P.ussuriensis),X代表新疆梨(P.sinkiangensis),Z(stock species)代表砧木類型。 對于自交親和性強度極弱的,筆者選擇了R>90%的品種(金墜、秋榮、晚秀、五九香、早冠、初夏綠)進一步探索自花授粉結實率,結果發現,金墜、秋榮、晚秀3個品種R值為100.00%,其中以已報道的自交親和品種金墜和秋榮作為對照。同時筆者也對自交不親和強度為中的早黃金、自交不親和強度為強的錦豐和六月爽多個品種進行了自花套袋試驗。表2結果表明,金墜和秋榮在花后20、30 d坐果率都高于20%,而其他品種自花套袋的品種基本沒有坐果。金墜和秋榮自花花粉管長到花柱最低端,并且坐果率大于20%,說明金墜和秋榮為自交親和性品種,同時通過自交不親和強度R值來初步篩選自交親和性品種的可靠性得到了一定的驗證。然而,并不是所有R值為100.00%的品種都是自交親和性品種,本試驗中,初步鑒定晚秀R值為100.00%,但通過自花套袋發現坐果率為0。其他自交親和性強度為強與中的品種自花套袋不結實,或者結實率極低,為自交不親和品種,進一步說明通過R值在海量的資源中初步篩選自交親和性品種方法的優越性及可靠性。 表2 自花授粉后果實坐果率統計 梨是薔薇科中自交不親和物種[16],自花花粉可以在柱頭上萌發,但是很少能生長到花柱底部[17-18]。本研究通過觀察不同品種自花花粉管在花柱內停止生長的位置差異發現,許多品種花粉萌發后生長很短或者剛能穿過柱頭就完全停止生長,表現為強自交不親和性,而大多數品種自花花粉管生長停止在花柱長度的2/3之前,這與前人的報道一致[17]。然而,仍有少量花粉管能夠生長至花柱基部,表現為弱自交不親和性。每個花粉管數量在花柱中的分布從柱頭到基部越來越少,且絕大多數花粉管最終在花柱內停止生長,花粉管末端形態結構改變。有活體試驗表明,不親和性花粉管生長受抑制前,有彎曲、先端膨大變形等現象,而異花授粉的花粉管沒有此現象[10]。也有研究表明,自花授粉的花粉管在花柱內都有“快—慢—快”的動態生長變化過程[10]。在不同的梨品種中花粉管生長受到抑制時,花柱的發生部位及變化不同,徐義流等認為可能是自交不親和基因時空表達的差異造成的[19]。也有研究表明,梨花柱中S糖蛋白的含量具有品種特異性和組織差異性,即品種不同,花柱中S糖蛋白的含量不同,并且S糖蛋白在花柱中分布不均勻導致花粉管停止生長[12,20]。梨的自交不親和性主要表現在自交授粉花柱中花粉管的生長受到抑制,這與S糖蛋白有直接的關系,S糖蛋白的核糖核酸酶活性可通過降解花粉管RNA來抑制花粉管生長[21]。活體和離體試驗均表明,自交不親和性強度與花粉管長度呈負相關[11-12]。從而分析認為,由花粉管生長的長度可以判斷自交不親和強度,自交花柱中花粉管生長長度占花柱的比例將自交不親和強度分為強、中和弱3類。在本研究中256個品種中,自交不親和性強度參差不齊,強、中和弱頻度分布也有很大差距。在離柱頭近的地方S-RNase濃度可能較高,因此自交不親和性為強的頻度分布占多數,為梨不同品種花柱內S糖蛋白總量及其花柱內的分布提供一定的理論基礎。S糖蛋白在花柱內是否具有梯度分布和怎樣的分布有待進一步驗證。本研究同時為后期深入研究自交不親和性強度具體機制提供了充足的基礎數據。 筆者通過對大量自花授粉花粉管生長狀態研究,來初步判斷篩選自交親和性品種。發現金墜、秋榮和晚秀花粉管長到了花柱最底端,自交不親和強度R值為100.00%。進一步自花套袋調查了坐果率,發現金墜和秋榮確實為自交親和性品種。然而遺憾的是,晚秀在鑒定花粉管過程中極可能是自交親和性品種,但最終自花套袋未結實。同時還發現,自交不親和強度呈現中和強的品種,在自花套袋試驗中也不結實。因此,這樣的結果驗證了筆者通過自花花粉管生長狀態來初步篩選自交親和性品種的必要性和可靠性,大大減少了盲目調查坐果率的工作量,并且豐富了自交不親和強度R值數據,為后期進一步研究自交不親和機制提供了充足的數據及一定的理論基礎。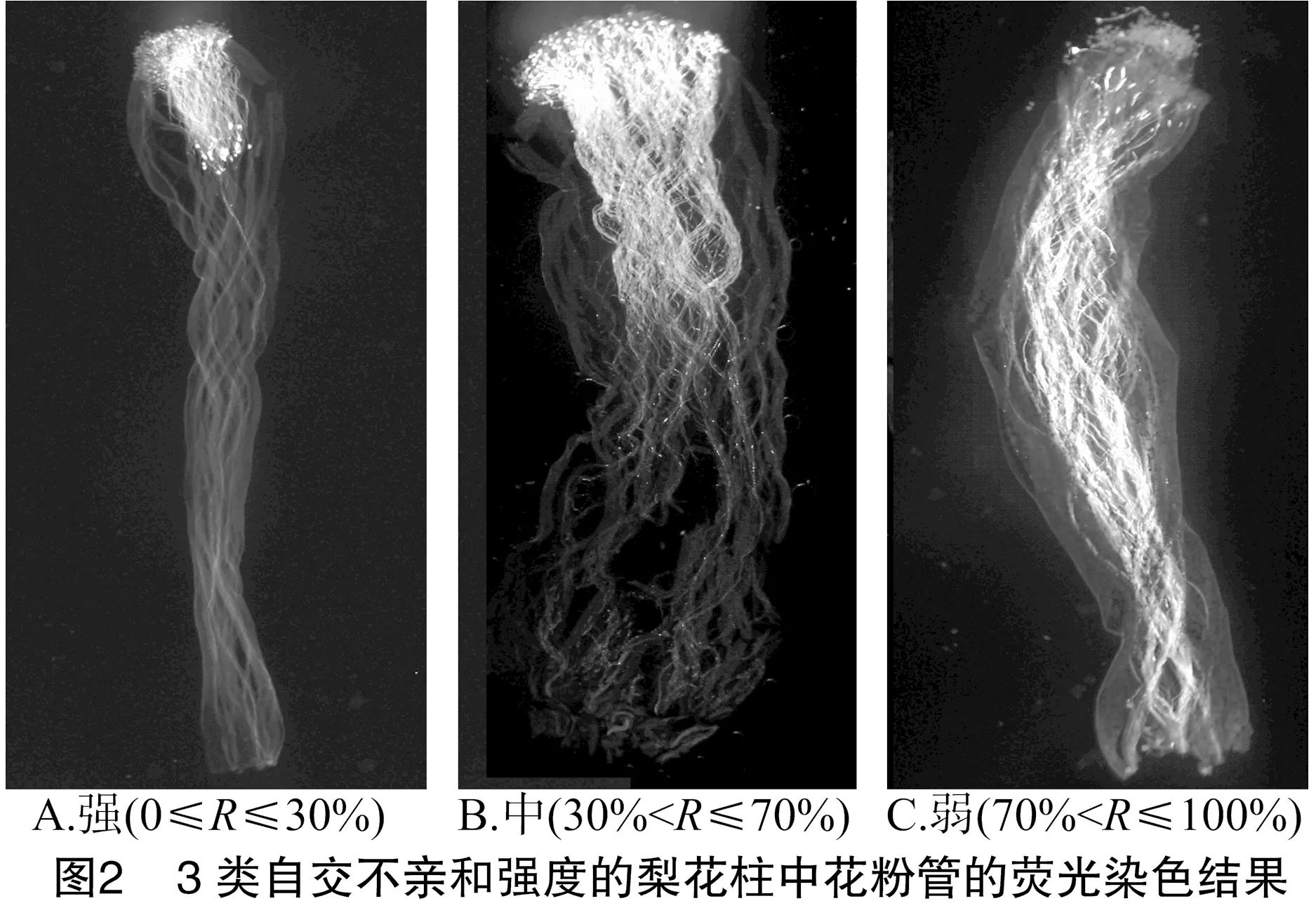
2.3 自交不親和強度R值分析
2.4 不同梨品種自交不親和性強度R值頻度分布

2.5 自花授粉坐果率統計調查
3 討論
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
