不同白羊草居群對干旱脅迫的生理響應及抗旱性評價
2018-07-30 03:29:06郭晉梅董寬虎
草地學報 2018年1期
關鍵詞:植物
鐘 華, 董 潔, 郭晉梅, 董寬虎
(1. 山西農業大學動物科技學院,山西 太谷 030801; 2. 北京市科學技術情報研究所,北京 100044)
干旱是導致植物形態、生理及基因表達變化的世界性問題,也是限制植物生長發育及增產的主要逆境因子之一[1-2]。因此,在淡水資源嚴重匱乏的環境壓力下,開展牧草抗旱性研究的應用價值就顯得格外重要[3]。白羊草[Bothriochloaischaemum(L.) Keng]為禾本科(Gramineae)孔穎草屬(Bothriochloa)多年生植物,具有產量高、營養價值豐富、適口性好、適應性強、再生力強、耐牧、耐踐踏等優良特性,主要分布在我國暖溫帶灌草叢地帶,在水土保持及飼草供應方面具有非常重要的意義[4-6]。白羊草能夠很好的適應其所處生境的水熱條件變化,從而成為響應全球氣候變化的主要植物,也是北方退化草地恢復與碳儲存的重要植物[7]。程林梅等[8]研究表明,白羊草在干旱脅迫下有良好的適應性和忍耐性。此外,適宜的氮磷肥料或適當濃度的CO2有助于增強白羊草對干旱脅迫的耐受性,緩解干旱脅迫造成的不利影響,提高白羊草的抗旱性[9-14]。劉娟等[15-16]發現,野生型白羊草抵御干旱脅迫的能力大于栽培型。然而,遺傳變異普遍存在于自然界中,天然居群由于地理隔離和山脈河流等天然屏障,有限的基因流可能會導致居群間遺傳分化加大,從而增加物種適應劇烈變化環境的潛力[17-18]。因此,不同居群白羊草的抗旱能力必然也會存在巨大差異,充分挖掘不同居群野生白羊草種質資源的抗旱性對其遺傳多樣性的應用具有重要意義。本研究選擇山西省境內10個不同地區的野生白羊草居群進行干旱脅迫,通過對其幼苗的抗旱性指標進行比較評價,旨在篩選出抗旱性較強的白羊草種質資源材料,為野生白羊草的馴化及其新品種選育提供一定的理論依據。
1 材料與方法
1.1 材料來源
試驗以采集山西省10個不同地區的野生白羊草為材料,其種子保存于山西農業大學草業科學系種質資源庫,各供試材料詳細信息如表1所示。
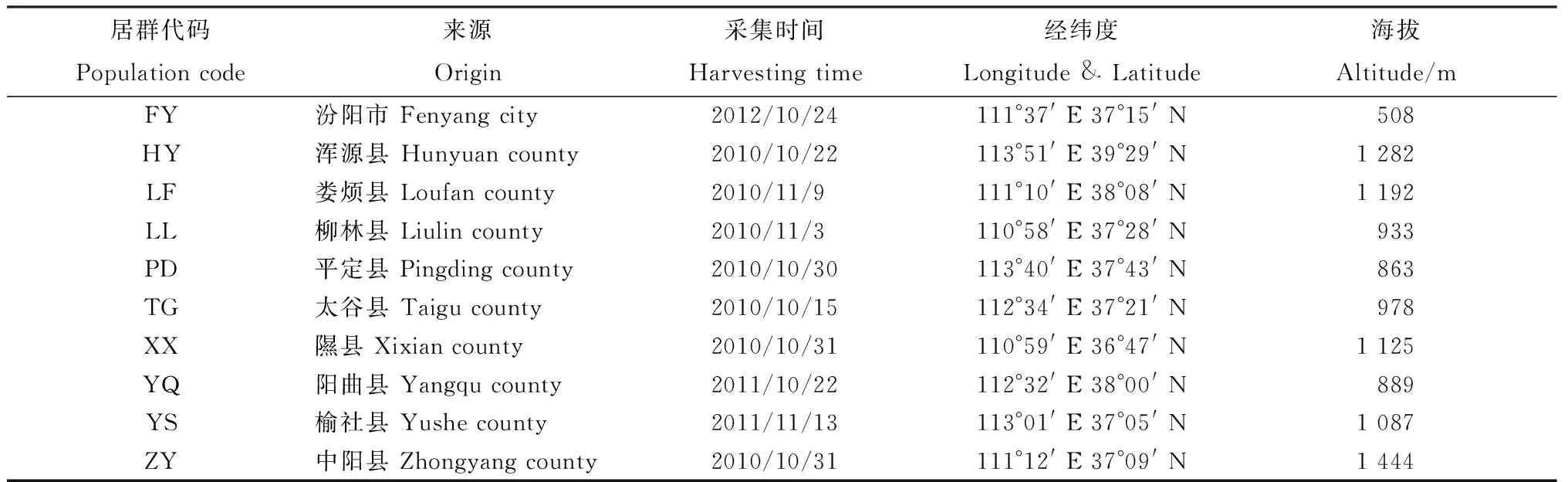
表1 材料來源Table1 Sources of materials
1.2 干旱處理
試驗采用盆栽法于山西農業大學草業科學實驗室日光溫室內進行,盆口內徑25 cm、高20 cm,盆栽用土采用山西農業大學動物科技試驗站牧草試驗田的耕層土(0~20 cm),其田間最大持水量為23.91%,pH為7.5。每盆裝過篩的風干土7 kg,于2013年7月5日播種,每盆播種50粒,播種后在土壤表面覆細沙以防土壤板結,苗期保持充分供水。苗齊后間苗,每盆留苗20株。待白羊草幼苗生長至平均20 cm時開始干旱處理,處理前1次性澆水,使桶內土壤含水量保持在19.13±3.38%(相當于田間持水量的80%)。每個居群設置4個干旱脅迫處理梯度(表2),每個處理設3次重復,共120盆,用稱重法控制土壤含水量。各個處理在達到設定水分梯度后第20天取樣,于-80℃冰箱中保存,用于測定各項指標。
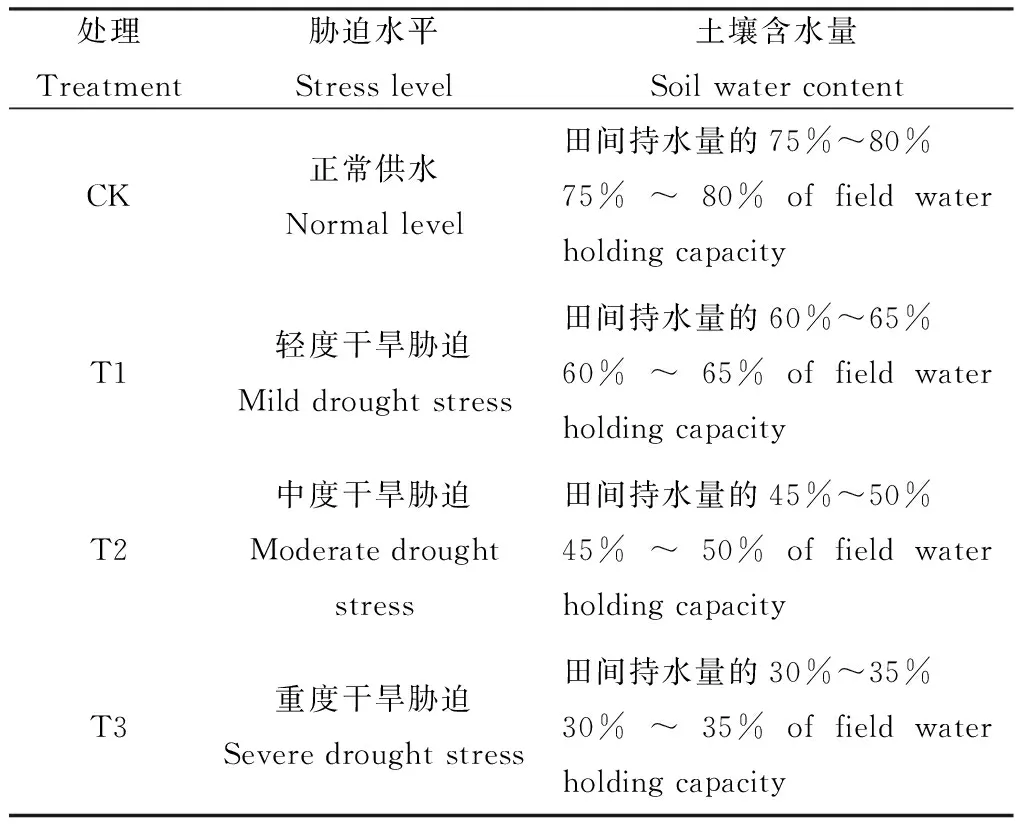
表2 干旱脅迫處理水平Table 2 Level of drought stress treatments
1.3 測定指標
1.3.1生長性能指標 取樣前用直尺測量每盆全部植株的高度(cm),并收集每盆全部植株的地上部分和地下部分,風干后測定其干重(g)作為地上生物量和地下生物量,以計算其根冠比(植株地下生物量/地上生物量)。
1.3.2生理指標 參照李合生的《植物生理生化實驗原理和技術》[19]。采用酸性茚三酮顯色法測定游離脯氨酸(proline,Pro)含量,采用硫代巴比妥酸法測定丙二醛(malondialdehyde,MDA)含量,采用氮藍四唑法測定超氧化物歧化酶(superoxide dismutase,SOD)活性,采用愈創木酚法測定過氧化物酶(peroxidase,POD)活性,采用蒽酮比色法測定可溶性糖(soluble sugar,SS)含量。
1.3.3耐旱隸屬函數值 利用模糊數學中隸屬函數法對不同材料耐旱性進行綜合評價。求出供試材料的各測定指標在不同脅迫程度下的隸屬值Zij,計算公式為:Zij=(Xij-Xjmin)/(Xjmax-Xjmin)。式中:Xij為i材料j指標在某一次采樣下的測定值,Xjmax為該次采樣j指標測定最大值,Xjmin為該次采樣的j指標最小值,如果j為負向指標那么就用反隸屬函數Zij=1-(Xij-Xjmin)/(Xjmax-Xjmin)。最后把j指標在不同處理下的隸屬值累加,在各指標權重相同的前提下求其平均值,該平均值即為i材料j指標的耐旱隸屬函數值[20]。
1.4 數據分析
所有數據均采用Excel 2010錄入及作圖,利用SPSS 18.0統計軟件對各參數進行單因素方差分析(ANOVA)、顯著性檢驗與多重比較(P=0.05)。
2 結果與分析
2.1 干旱脅迫對不同居群白羊草生長性能的影響
不同居群白羊草株高均隨干旱脅迫程度的增加而呈下降趨勢,但各居群下降幅度存在一定的差異(表3),因對不同干旱脅迫程度的響應而異。在各干旱脅迫程度下,LF居群的白羊草株高均最大,顯著(P<0.05)高于其他居群;LL居群的白羊草株高最小,顯著(P<0.05)低于其他居群。在重度脅迫(T3)下,ZY居群相對CK的降低幅度最大,比CK下降50.52%;而XX居群降幅最小,下降20.01%。
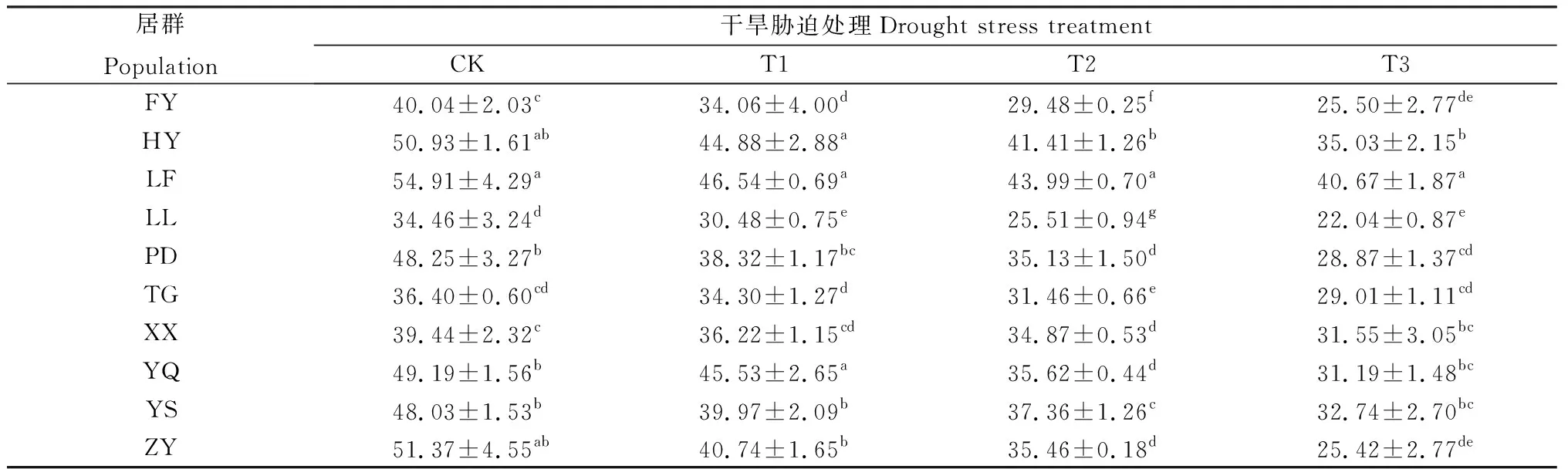
表3 不同干旱脅迫水平下白羊草株高Table 3 Plant height under different drought stress level/cm
注:同列不同小寫字母表示相同脅迫水平不同居群間差異顯著(P<0.05),下同
Note:Different lowercase letters within the same column indicate significant differences among different populations with the same drought stress at the 0.05 level. The same as below
在干旱脅迫時,植株通過加強根系生長來提高其對深層土壤水分和養分的吸收利用,進而得以維持其正常生長發育。不同居群白羊草根冠比均隨干旱脅迫程度的加劇而增加,但各居群變化幅度不同(表4)。輕度脅迫(T1)下,ZY居群增幅最大,比CK增加了83.33%;YS居群增幅最小,僅23.53%。中度脅迫(T2)下,LL居群增幅最大,比輕度脅迫(T1)增大了1倍;YS居群仍增幅最小,為23.81%。在重度脅迫(T3)下,ZY居群白羊草的根冠比最大,且其變化幅度也最大,較中度脅迫(T2)增大了3.25倍。
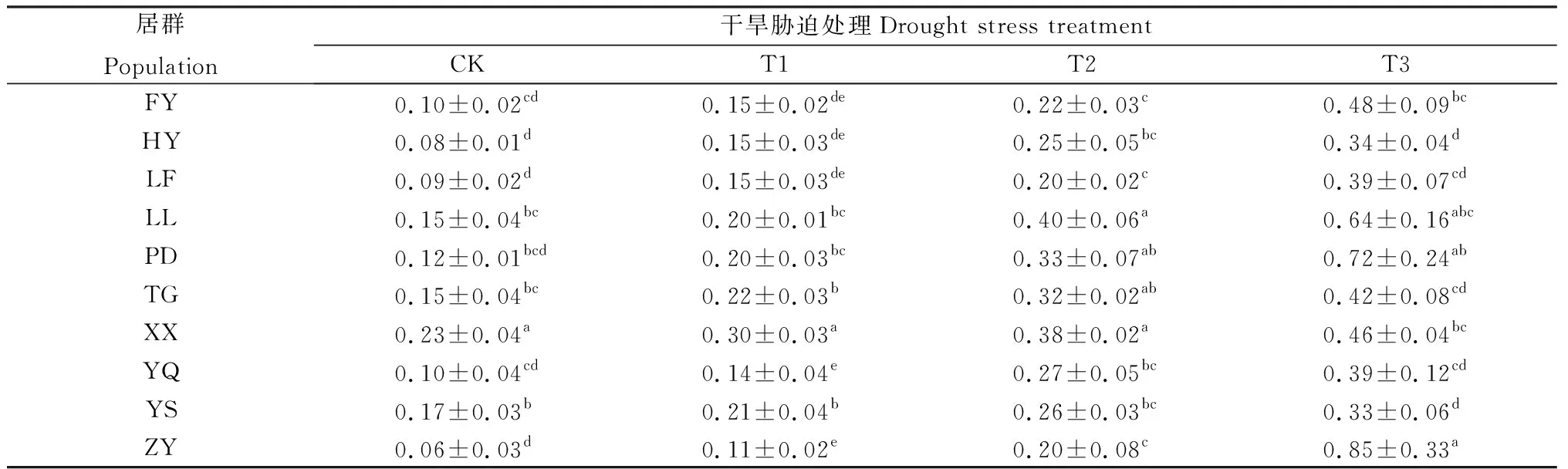
表4 不同干旱脅迫水平下不同居群白羊草根冠比Table 4 Root shoot ratio under different drought stress level
2.2 干旱脅迫對不同居群白羊草游離脯氨酸(Pro)含量的影響
不同居群白羊草Pro含量均隨干旱脅迫程度的增加而呈升高趨勢,但不同居群間變化程度不同(圖1)。在正常供水(CK)及輕度(T1)、中度(T2)脅迫下,LF居群白羊草Pro含量均最大,顯著(P<0.05)高于其他居群;而XX居群白羊草Pro含量均最小,顯著(P<0.05)低于其他居群。在重度脅迫(T3)下,FY居群白羊草Pro含量最高,顯著(P<0.05)高于其他居群,且與CK相比增加幅度最大,此時Pro含量是CK的13.27倍;而YS居群白羊草Pro含量最低,顯著(P<0.05)低于其他居群,且與CK相比增幅最小,僅為CK的4.22倍。
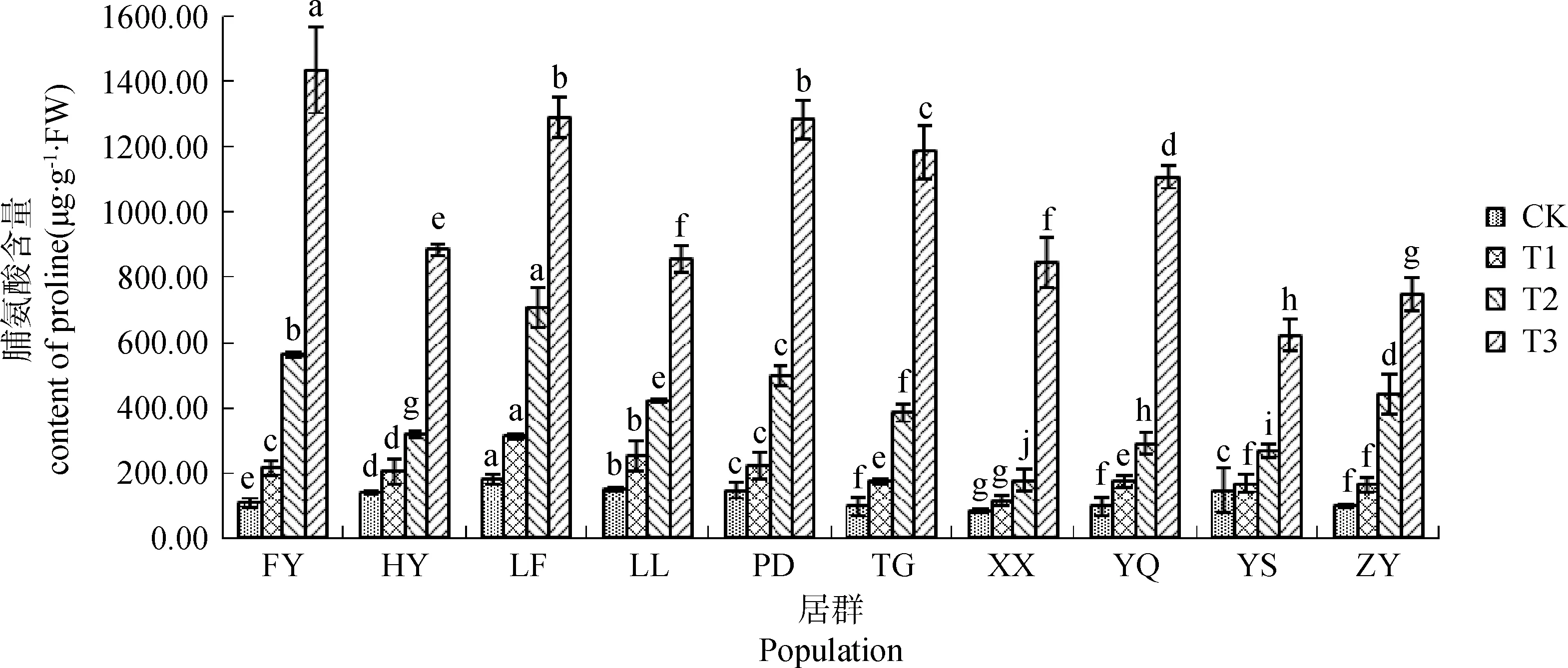
圖1 不同干旱脅迫水平下游離脯氨酸(Pro)含量的變化Fig.1 The change of Pro contents under different drought stress level注:相同處理下不同小寫字母表示不同居群間差異顯著(P<0.05),下同Note:Different lowercase letters in the same treatment indicate significant differences among different populations at the 0.05 level. The same as below
2.3 干旱脅迫對不同居群白羊草丙二醛(MDA)含量的影響
MDA是膜脂過氧化產物,它的含量決定了植物的受害程度。在干旱脅迫下,不同居群白羊草的MDA含量均隨干旱脅迫程度增加而呈上升趨勢,但不同居群之間增加幅度不同(圖2)。LF居群變化幅度最大,在正常供水(CK)下,LF居群MDA含量最低,為22.53 μmol·g-1·FW,而在重度脅迫(T3)下含量達到40.25 μmol·g-1·FW,比CK增加了78.65%,增幅最大。重度脅迫下MDA含量最低的是XX和FY居群,且其MDA含量較CK增加幅度最小,分別增加了48.85%和48.97%。
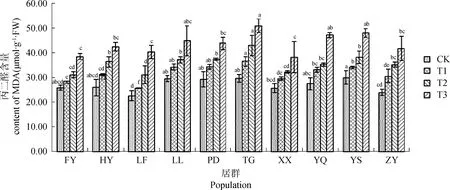
圖2 不同干旱脅迫水平下丙二醛(MDA)含量的變化Fig.2 The change of MDA contents under different drought stress level
2.4 干旱脅迫對不同居群白羊草超氧化物歧化酶(SOD)活性的影響
不同居群白羊草SOD活性均隨干旱脅迫程度增加而呈上升趨勢,且各居群間變化幅度存在差異(圖3)。在各水分條件下,XX居群白羊草的SOD活性最大,均顯著(P<0.05)高于其他居群。隨著干旱脅迫程度的加劇,在中度脅迫(T2)下,LF居群白羊草SOD活性比CK增加了47.71%,在各居群間增幅最大;在重度脅迫(T3)下,YS居群增幅最大,比CK增加了1.05倍。
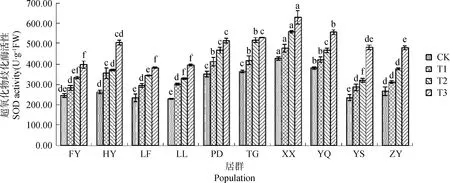
圖3 不同干旱脅迫水平下SOD活性的變化Fig.3 The change of SOD activity under different drought stress level
2.5 干旱脅迫對不同居群白羊草過氧化物酶(POD)活性的影響
隨干旱脅迫程度增加,不同居群白羊草POD活性變化存在差異(圖4)。FY、LF、LL、XX和YQ居群白羊草POD活性呈上升趨勢,而HY、PD、TG、YS和ZY居群白羊草POD活性呈先升后降趨勢,且均在中度干旱脅迫(T2)時達到最高水平。在中度脅迫(T2)下,TG居群白羊草POD活性增幅最大,比CK增加了1.5倍,但隨脅迫程度加劇活性下降。在重度脅迫(T3)下,LL居群白羊草POD活性顯著(P<0.05)高于其他居群,相比CK增幅最大,增加了1.12倍。
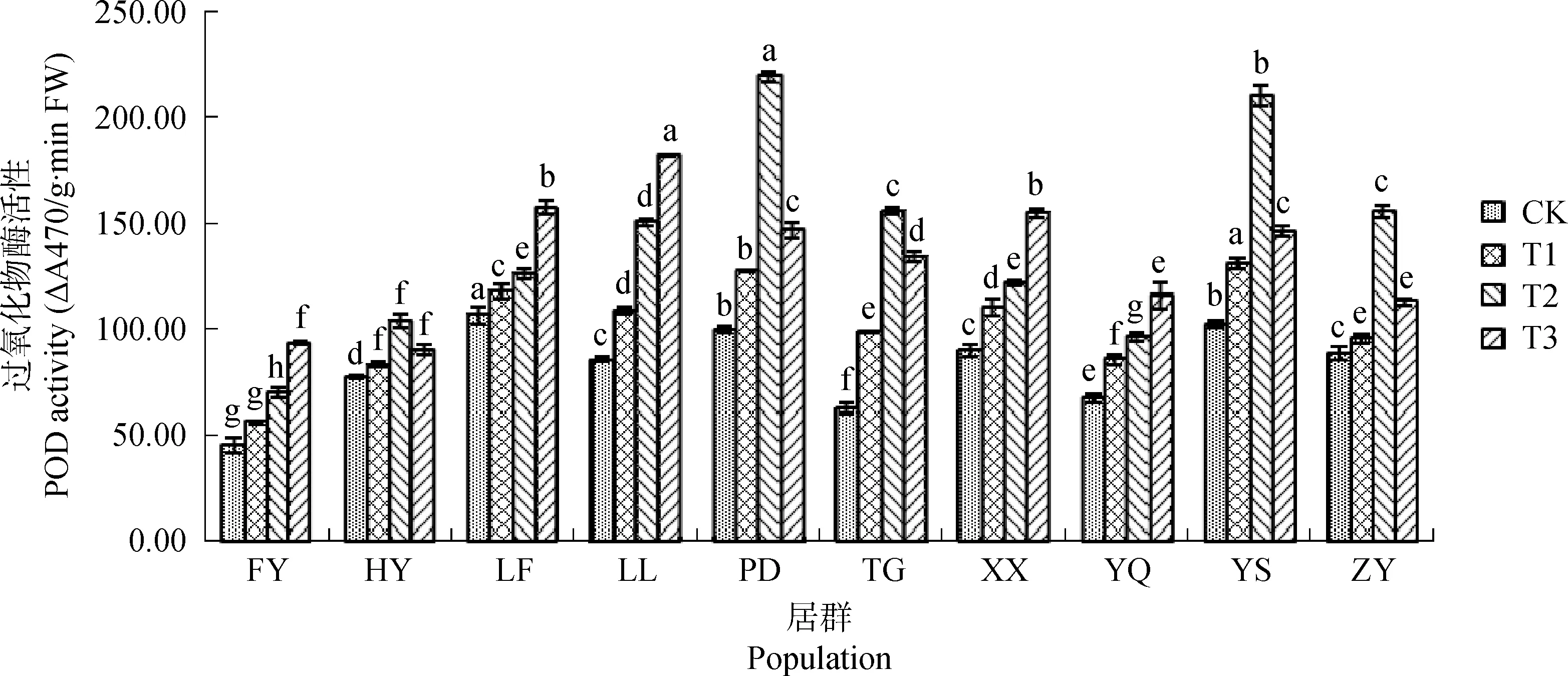
圖4 不同干旱脅迫水平下POD活性的變化Fig.4 The change of POD activity under different drought stress level
2.6 干旱脅迫對不同居群白羊草可溶性糖(SS)含量的影響
不同居群白羊草SS含量均隨干旱脅迫程度增加而呈上升趨勢,且各居群間變化幅度存在差異(圖5)。在正常供水(CK)條件下,FY、LF、LL、PD、YS和ZY居群白羊草SS含量顯著(P<0.05)高于HY、TG、XX和YQ居群,但隨著干旱脅迫程度的增加,到重度脅迫(T3)下,TG居群白羊草的SS含量較CK增加了1.35倍,而ZY居群僅增加了0.62倍。在中度(T2)和重度(T3)脅迫下,FY居群白羊草SS含量顯著(P<0.05)高于其他居群,而YQ居群白羊草SS含量顯著(P<0.05)低于其他居群。
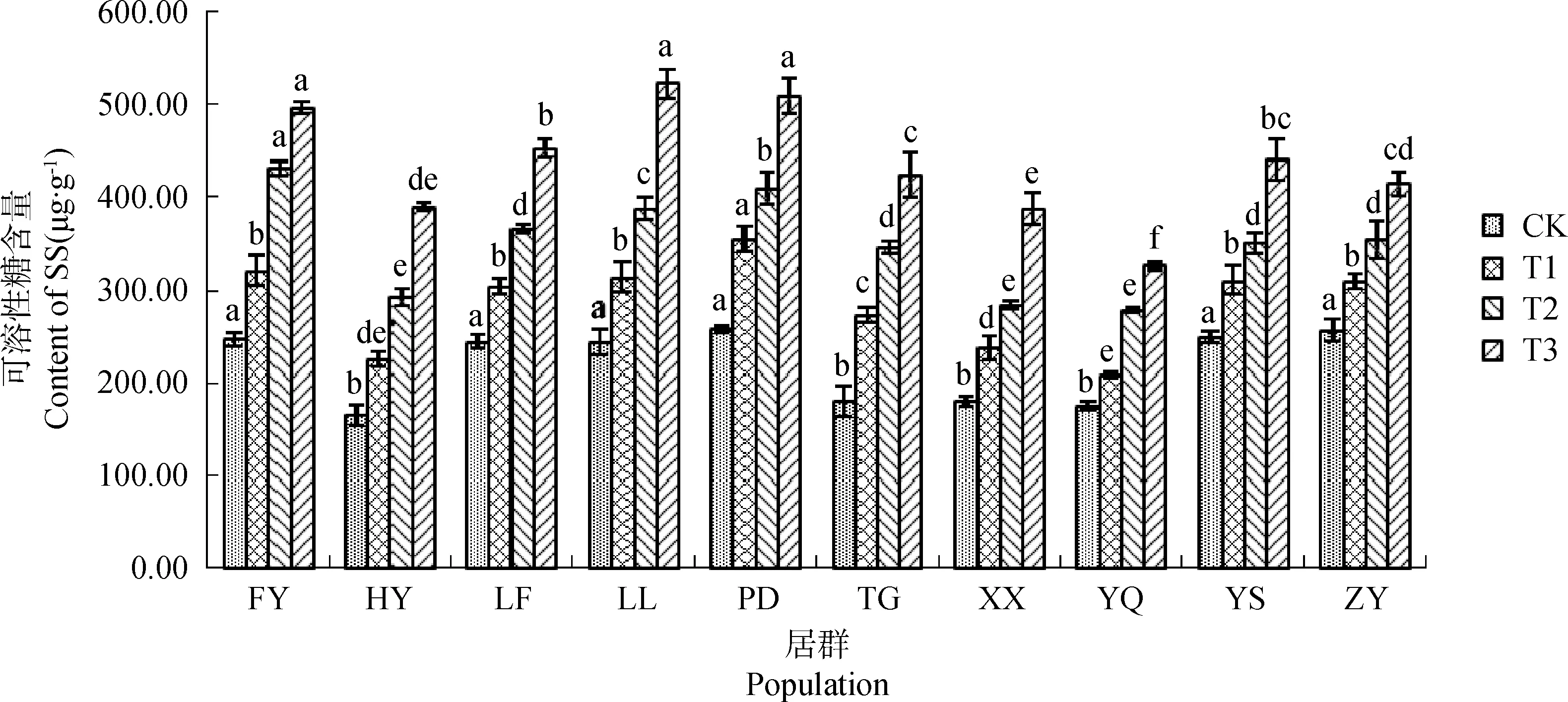
圖5 不同干旱脅迫水平下SS含量的變化Fig.5 The change of SS content under different drought stress level
2.7 隸屬函數分析
利用隸屬函數法對上述7個指標進行綜合分析表明(表5),FY居群白羊草隸屬值最大,為0.618,抗旱性最強;YQ居群白羊草隸屬值最小,為0.383,抗旱性最弱。不同居群白羊草的抗旱性強弱順序為:FY >LF>TG>XX>LL>ZY>YS>PD>HY>YQ。
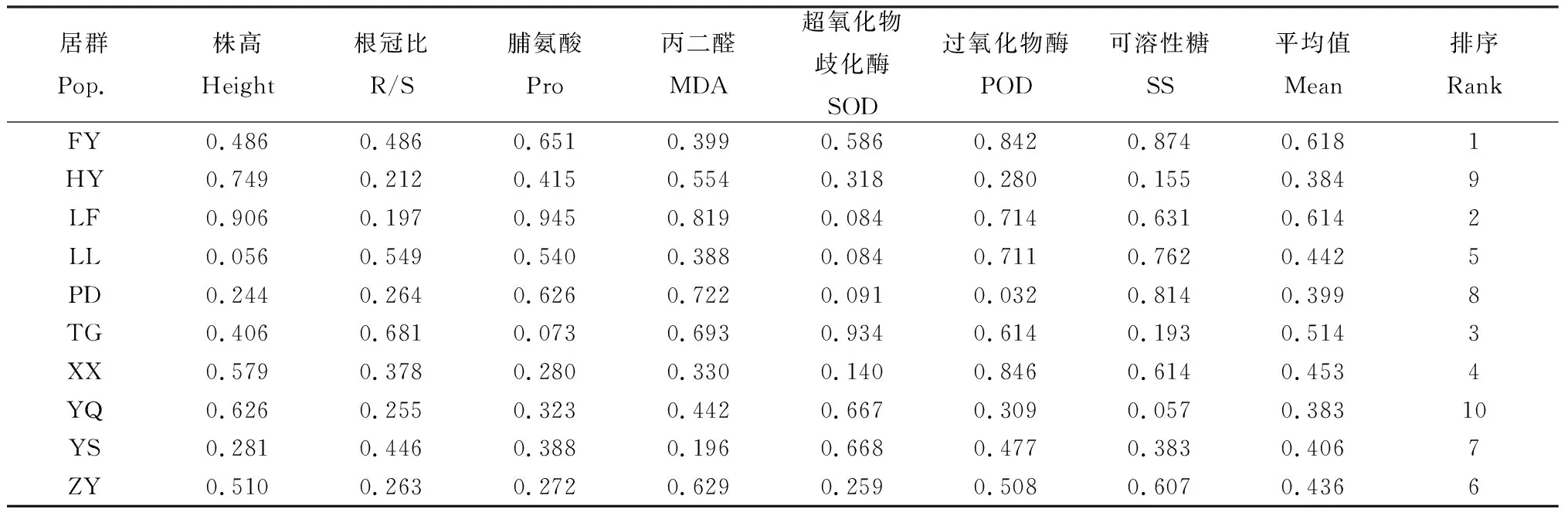
表5 不同居群白羊草抗旱隸屬函數值比較Table 5 Comparison of subordinate function value of different populations of old world bluestem
3 討論
植物在干旱脅迫下會從形態結構、生理生化等多層次上表現出相關的系統適應性[21]。本試驗結果表明,隨干旱脅迫程度的增加,不同居群白羊草的株高均呈下降趨勢,而其根冠比均呈增大趨勢,但是不同居群變化程度存在差異,表明不同居群白羊草對干旱脅迫的適應性存在差異。這與易津等[22]研究賴草屬(Leymus)牧草幼苗耐旱性的結果相一致。
滲透調節物質的積累是植物適應干旱脅迫的基本特征之一,Pro積累可以增加植物對干旱脅迫的耐性[23-24]。本研究表明,不同居群白羊草Pro含量均隨干旱脅迫程度增強而升高,且升高幅度有所不同,表明不同居群白羊草的抗旱能力存在差異。其中FY居群白羊草在不同脅迫程度下Pro含量較高,尤其在重度脅迫(T3)下達到最高,表明Pro的積累有可能提高了該居群的抗旱耐性,因此,隸屬函數分析表明FY居群白羊草抗逆性最強。SS也是植物體內一類重要的滲透調節物質,在植物對水分脅迫的適應性調節中,逐漸積累進行滲透調節,阻止細胞膜解離,增強細胞保水能力,穩定細胞結構,防止細胞脫水[20]。本研究發現,不同居群白羊草SS含量隨干旱脅迫程度增加而升高,這與武燕奇[20]、史玉煒等[25]的研究結果相符。此外,不同居群白羊草SS含量增加幅度存在差異,說明不同居群白羊草的抗旱能力不同。其中TG居群白羊草的SS含量隨脅迫程度的加劇而增加幅度最大,表明其具有較強的滲透調節能力,抗旱性也較強。所以隸屬函數分析表明TG居群白羊草的抗旱性能位居第3位。
干旱脅迫會破壞植物細胞內氧自由基產生與清除的平衡,從而導致細胞膜脂質過氧化,使植物受到傷害,并產生MDA,MDA含量越大,表明植物受干旱脅迫越嚴重[26]。本試驗表明,不同居群白羊草MDA含量隨干旱脅迫程度加劇而升高,表明白羊草幼苗受到的傷害逐漸增大,這與朱慧森[27]、郭郁頻[28]的研究結論一致。FY居群白羊草的MDA含量較低,且其增加的幅度較小,因而其抗旱能力最強。干旱脅迫時,植物體內SOD和POD可以清除過量積累的活性氧,從而保護植物免遭傷害[29]。因此,干旱脅迫下,抗氧化酶酶活性越高,消除活性氧的能力越強,植物的抗逆性也越強[30]。本研究中,不同居群白羊草SOD和POD活性均隨干旱脅迫程度加劇而升高,這表明更多的活性氧被清除[31-32]。然而,HY、PD、TG、YS和ZY居群白羊草的POD活性隨干旱脅迫程度加劇而呈先升后降的趨勢,并在中度脅迫(T2)時達到最高水平,可能與POD活性在重度干旱脅迫下遭到破壞有關,這與季楊等[3]在干旱脅迫對鴨茅(Dactylisglomerata)的POD活性的影響的結果一致。
4 結論
干旱脅迫下,10個野生居群白羊草株高均隨脅迫程度的增加而降低,其根冠比均隨脅迫加劇而增加,其Pro、MDA和SS含量及SOD和POD活性均隨脅迫程度加劇而增加。采用隸屬函數法綜合分析得出其抗旱性強弱順序為:汾陽居群>婁煩居群>太谷居群>隰縣居群>柳林居群>中陽居群>榆社居群>平定居群>渾源居群>陽曲居群。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
