裸果木種群遺傳多樣性特點(diǎn)及與地理氣候因子關(guān)聯(lián)研究
2018-07-30 03:28:56徐振朋張曉明伊衛(wèi)東宛詣超
草地學(xué)報(bào) 2018年1期
關(guān)鍵詞:物種
徐振朋, 宛 濤*, 蔡 萍, 張曉明, 伊衛(wèi)東, 宛詣超
(1. 內(nèi)蒙古農(nóng)業(yè)大學(xué)草原與資源環(huán)境學(xué)院,內(nèi)蒙古 呼和浩特 010019; 2. 錫林郭勒職業(yè)學(xué)院,內(nèi)蒙古 錫林浩特 026000;3. 內(nèi)蒙古農(nóng)業(yè)大學(xué)農(nóng)學(xué)院,內(nèi)蒙古 呼和浩特 010019)
裸果木為石竹科(Caryophyllaceae)裸果木屬(Gymnocarpos)多年生小半灌木,國(guó)家一級(jí)保護(hù)植物,起源于第三紀(jì),古地中海殘遺成分[1-2]。分布于甘肅河西走廊、內(nèi)蒙古西部、寧夏中西部和新疆東部[3]。生境較為單一,主要生長(zhǎng)在海拔為800~2 500 m荒漠區(qū)的干河床、干河道、山前洪積扇及礫石戈壁灘等地。在防風(fēng)固沙、水土保持方面發(fā)揮十分重要的作用,對(duì)維護(hù)當(dāng)?shù)厣鷳B(tài)系統(tǒng)穩(wěn)定具有著不可替代的作用[1]。分布區(qū)年均溫為4.1~9.3℃,≥10℃年積溫在3 000~4 000℃之間,極端最高溫達(dá)40.0℃,極端最低溫達(dá)到-31.6℃;年均日照時(shí)數(shù)為2 900~3 400 h左右;年均降水量50~170 mm之間,集中在7~9月;年蒸發(fā)量2 100~3 300 mm。從植物區(qū)系組成及水熱特征來(lái)看,為典型的戈壁荒漠和荒漠化草原。目前對(duì)于裸果木主要集中于種群生態(tài)學(xué)[4-5]、分類學(xué)[6-7]、繁殖生物學(xué)[8-10]等方面研究。因群落結(jié)構(gòu)簡(jiǎn)單、生態(tài)系統(tǒng)脆弱、自然地理環(huán)境變遷及人類活動(dòng)干擾等因素,使裸果木種群斑塊狀分布格局更趨于明顯,分布區(qū)面積縮小,種群數(shù)量銳減。
遺傳多樣性是生物體攜帶遺傳信息的總和,是生物多樣性的重要組成部分,維持生物繁殖活力及與環(huán)境變化相互適應(yīng)的基礎(chǔ)。物種在受環(huán)境壓力的影響,除逃避和表型可塑性外,主要是通過(guò)改變遺傳基礎(chǔ)來(lái)適應(yīng)環(huán)境[11]。因此,種群或物種遺傳多樣性越高,對(duì)環(huán)境變化的適應(yīng)能力越強(qiáng),越容易擴(kuò)展其分布范圍[12]。簡(jiǎn)單序列重復(fù)區(qū)間擴(kuò)增多態(tài)性(inter simple sequence repeat,ISSR)分子標(biāo)記技術(shù)以簡(jiǎn)便快捷、多態(tài)性高和重復(fù)性好等優(yōu)點(diǎn)在遺傳多樣性評(píng)價(jià)方面得到廣泛應(yīng)用[13-15]。為深入了解裸果木遺傳基礎(chǔ)、斑塊狀分布的種群遺傳結(jié)構(gòu)、地理氣候因素對(duì)種群遺傳多樣性水平的影響,以分布我國(guó)甘肅河西走廊及內(nèi)蒙古西部地區(qū)裸果木種群為研究對(duì)象,利用ISSR遺傳標(biāo)記技術(shù)對(duì)不同地理種群進(jìn)行遺傳評(píng)價(jià)。從分子水平上探討裸果木種群間的親緣關(guān)系及遺傳多樣性水平,進(jìn)而為其種質(zhì)資源的科學(xué)化管理及保育提供依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料
2015年5月對(duì)分布于內(nèi)蒙古、寧夏、新疆及甘肅河西走廊區(qū)域12個(gè)種群進(jìn)行了考察,每個(gè)種群采集20個(gè)植株健康葉片,每株間隔距離10 m以上共計(jì)采集240株。將采集的新鮮葉片立即放入加變色硅膠自封袋中干燥,帶回實(shí)驗(yàn)室保存于-80℃冰箱中備用。記錄樣地經(jīng)緯度、海拔等地理信息及收集當(dāng)?shù)貧庀蟛块T(mén)提供近10年離樣地最近的氣象數(shù)據(jù)。此外,由于裸果木多生長(zhǎng)在偏僻無(wú)人的地區(qū),部分氣象站點(diǎn)距離樣地較遠(yuǎn)。因此,通過(guò)DIVA軟件進(jìn)行當(dāng)前氣候數(shù)據(jù)提取(精確度為2.5 min),對(duì)樣地氣候數(shù)據(jù)進(jìn)一步校準(zhǔn)及補(bǔ)充,樣地地理及氣候信息如表1所示。
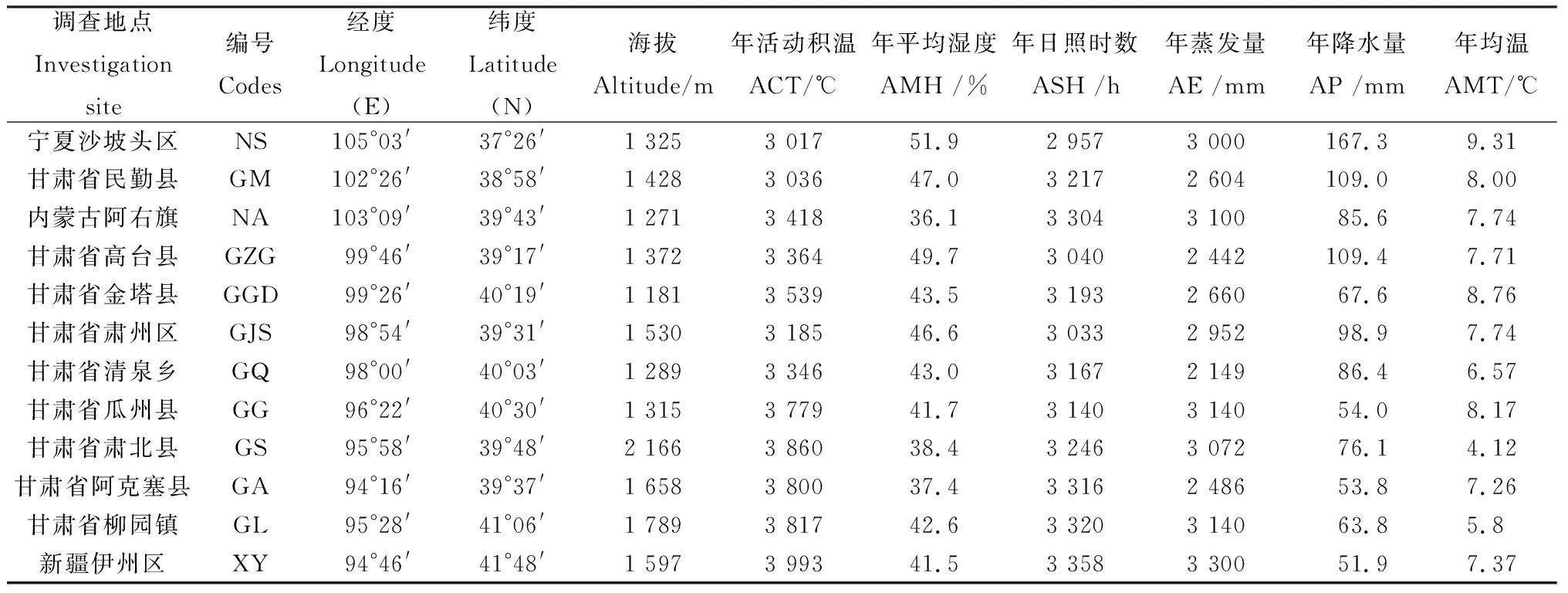
表1 裸果木種群試驗(yàn)區(qū)及樣地地理及氣候信息Table 1 The test site and sample plot’s information of the population of Gymnocarpos przewalskii
1.2 研究方法
1.2.1DNA提取 采用改進(jìn)CTAB法進(jìn)行DNA提取[16];利用紫外分光光度法檢測(cè)DNA濃度,并用1.0%的瓊脂糖凝膠電泳檢測(cè)其完整性,-20℃環(huán)境保存?zhèn)溆谩?/p>
1.2.2引物篩選與PCR擴(kuò)增 采用UBC(GA)8C引物進(jìn)行裸果木PCR擴(kuò)增條件篩選,初步確定ISSR-PCR的最佳擴(kuò)增程序?yàn)椋?4℃預(yù)變性3 min;94℃變性30 s,54℃復(fù)性45 s,72℃延伸1.5 min,35個(gè)循環(huán);72℃延伸10 min,4℃保存。正交試驗(yàn)優(yōu)化ISSR-PCR反應(yīng)體系:在25 μL反應(yīng)體系中dNTP 濃度為0.30 mmol·L-1,引物0.50 μmol·L-1,Mg2+為2.50 mmol·L-1,Taq DNA酶為1.00 U·μL-1,DNA 濃度為40.00 ng·μL-1效果最佳。依據(jù)反應(yīng)體系對(duì)100 個(gè) ISSR引物進(jìn)行篩選,從中選取擴(kuò)增條帶清晰,重復(fù)性較好的引物。
1.3 數(shù)據(jù)分析
用POPGENG 32軟件對(duì)種群間和種群內(nèi)遺傳參數(shù)分析,利用NTSYS-pc 2.1軟件采用UPGMA法構(gòu)建種群親緣關(guān)系樹(shù)狀聚類圖;建立不同種群裸果木的地理(經(jīng)緯度)的距離矩陣,利用SAS 9.0軟件對(duì)遺傳距離與地理距離進(jìn)行相關(guān)性檢測(cè)。同時(shí),將遺傳參數(shù)與氣候因子進(jìn)行相關(guān)性分析。
2 結(jié)果分析
2.1 裸果木ISSR標(biāo)記引物篩選
對(duì)100條ISSR引物進(jìn)行篩選,最終篩選出15條擴(kuò)增清晰,重復(fù)性好的引物,用于不同種群裸果木的遺傳多樣性分析,引物擴(kuò)增條帶及退火溫度如表2所示。對(duì)12個(gè)種群的240個(gè)裸果木植株進(jìn)行擴(kuò)增,總計(jì)擴(kuò)增出條帶數(shù)為143,具有多態(tài)為條帶數(shù)127,引物多態(tài)位平均比例為88.81%。
2.2 裸果木種群遺傳多樣性分析
不同種群裸果木遺傳多態(tài)參數(shù)如表3所示,其中多態(tài)位點(diǎn)百分率(PPL)變化范圍為61.54%~81.12%,平均多態(tài)位點(diǎn)百分率(PPL)為69.93%,較高的有甘肅省阿克塞縣(GA)種群(81.12%)及甘肅省肅北縣(GS)種群(72.73%)等,其次分別為寧夏沙坡頭區(qū)(NS)、甘肅省柳園鎮(zhèn)(GL)、甘肅省清泉鄉(xiāng)(GQ)及甘肅省金塔縣(GGD)等種群,最低為內(nèi)蒙古阿右旗(NA)種群(61.54%)。種群中有效等位基因數(shù)(Ne)變化范圍為1.4035~1.4624,較高的為甘肅省柳園鎮(zhèn)(GL)種群,最低為甘肅省金塔縣(GGD)種群。種群間He變化范圍為0.2356~0.2697,平均值為0.2418;種群間I變化為0.3509~0.4057,平均值為0.3629。甘肅省阿克塞(GA)與甘肅省肅北縣(GS)種群的He及I值均高于其他種群,具有豐富的遺傳多樣性,而內(nèi)蒙古阿拉善右旗(NA)及甘肅金塔縣(GGD)2個(gè)種群He及I較低。
注:Na:觀測(cè)等位基因數(shù);Ne:有效等位基因數(shù);He:種群內(nèi)遺傳多樣性;I:Shannon 信息指數(shù);PPL:多態(tài)位點(diǎn);下同
Note:Na: Observed number of alleles;Ne:Effective number of alleles;He: Gene Diversity within Populations;I:Shannon information index;PPL: Percentage of polymorphic loci;the same as below
裸果木種群基因多樣性分析如表4所示,其中總的遺傳多樣性(Ht)為0.3452,而種群內(nèi)的遺傳多樣性(Hs)為0.2489,群體間遺傳分化系數(shù)Gst為0.2790,即種群間的遺傳變異占總變異的27.90%,表明裸果木遺傳多樣性主要存在于種群內(nèi),而種群間也存在一定的分化。種群間基因流(Nm)為1.2920,表明種群之間存在著遺傳交換。
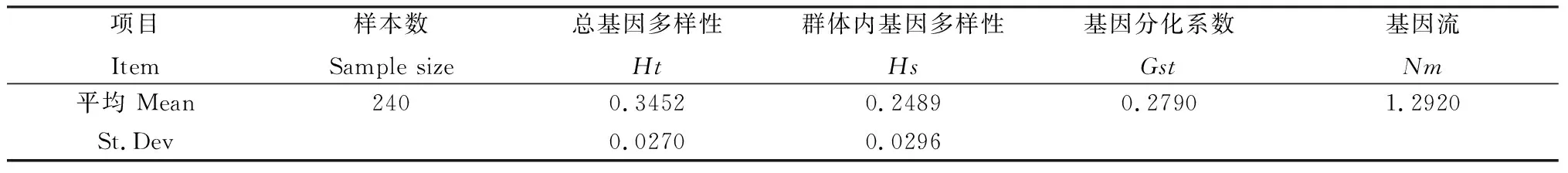
表4 ISSR標(biāo)記對(duì)裸果木種群基因多樣性Nei’s分析Table 4 Nei’s analysis of genetic diversity of Gymnocarpos przewalskii population by ISSR
2.3 遺傳多樣性參數(shù)與地理氣候因子相關(guān)性分析
裸果木地理距離與遺傳距離之間的相關(guān)性分析如表5所示,親緣關(guān)系最近的甘肅省高臺(tái)縣(GZG)和甘肅省肅州區(qū)(GJS)的遺傳一致度為0.9929,其地理距離約為79.19 km,而地理距離約為1004.78。寧夏沙坡頭區(qū)域(NS)和新疆伊州區(qū)(XY)種群遺傳一致度為0.7622,裸果木種群的遺傳距離與地理距離存在一定的相關(guān)性(R2=0.2144)。

表5 裸果木種群遺傳距離和地理距離Table 5 Nei’s genetic distance and geographic distance of Gymnocarpos przewalskii
注:遺傳一致度(對(duì)角線下方)和地理距離(對(duì)角線上方)
Note: Genetic identity (below diagonal)and geographic distance (above diagonal) /km
依據(jù)Nei’s遺傳一致度對(duì)12個(gè)種群裸果木進(jìn)行UPGMA聚類分析,結(jié)果如圖1所示,以河西走廊中西部為中心劃分為4類群:甘肅省高臺(tái)縣(GZG)、甘肅省清泉鄉(xiāng)(GQ)、甘肅省肅州區(qū)(GJS)、甘肅省金塔縣(GGD)為中心區(qū)的東部為一類,與向東和向東南延伸的內(nèi)蒙古阿右旗(NA)、寧夏沙坡頭區(qū)域(NS)和甘肅省民勤縣(GM)親緣關(guān)系較近;甘肅省瓜州(GG)、甘肅省肅北縣(GS)及甘肅省柳園鎮(zhèn)(GL)為中心區(qū)的西部區(qū)種群,與向西南和向西延伸的甘肅省阿克塞縣(GA)和新疆伊州區(qū)(XY)種群親緣關(guān)系較近。
對(duì)裸果木種群ISSR標(biāo)記的各遺傳多樣性參數(shù)與地理氣候因子進(jìn)行相關(guān)分析,結(jié)果如表6所示,ISSR標(biāo)記的裸果木遺傳多樣性與地理、氣候因子存在一定的相關(guān)性,遺傳參數(shù)Ne、He及I與經(jīng)度呈顯著負(fù)相關(guān)(P<0.05),與海拔呈現(xiàn)極顯著正相關(guān)(P<0.01),與年均溫呈現(xiàn)顯著負(fù)相關(guān)(P<0.05)。
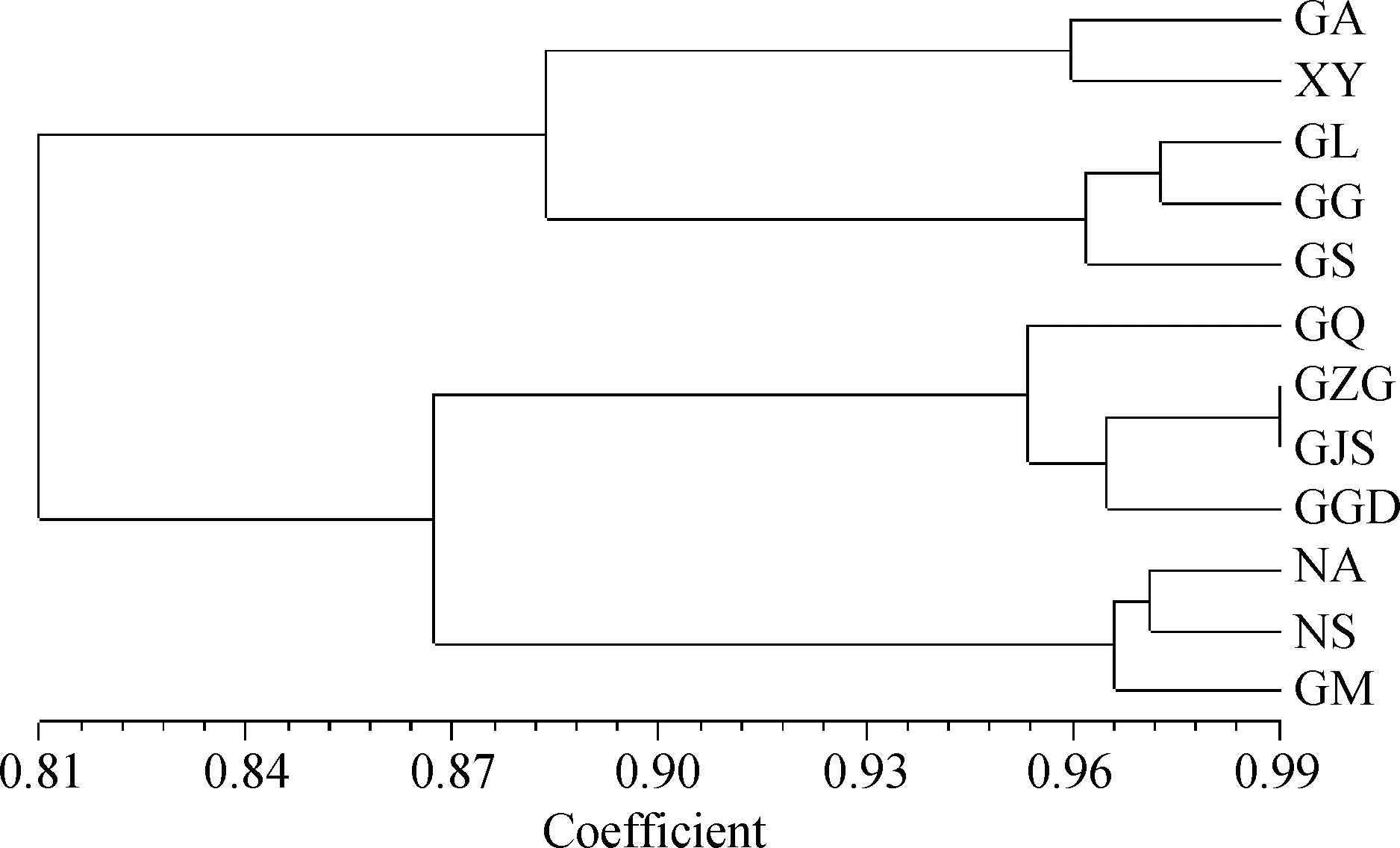
圖1 基于ISSR的12個(gè)裸果木種群的UPGMA聚類圖Fig.1 UPGMA dendrogram of 12 Gymnocarpos przewalskii clusters based on ISSR

表6 裸果木遺傳參數(shù)與地理氣候因子相關(guān)性分析Table 6 Correlation analysis between geographic climatic factors and genetic diversity of Gymnocarpos przewalskii
注: *表示P<0.05水平;**表示P<0.01水平
Note: *indicates the diffence at the 0.05 level;** indicates the diffence at the 0.01 level
3 討論與結(jié)論
3.1 裸果木種群遺傳多樣性特點(diǎn)
遺傳多樣性水平高低與多因子有關(guān),如分布區(qū)、生境的變化、繁育系統(tǒng)、個(gè)體細(xì)胞突變、奠基者效應(yīng)、更新世產(chǎn)生的物種遺留區(qū)等[17]。荒漠瀕危植物因繁殖方式、分布大小及生長(zhǎng)環(huán)境的限制,如綿刺(Potaniniamongolica)、蒙古扁桃(Amygdalusmongolica)等常常表現(xiàn)為相對(duì)較低的遺傳多樣性水平[18-19]。然而也有研究發(fā)現(xiàn)瀕危或分布狹窄的物種由于區(qū)系的古老性表現(xiàn)出較高的遺傳多樣性水平,如四合木(Tetraenamongolica)及長(zhǎng)葉紅砂(Reaumuriatrigyna)等[20-21]。Nybom等認(rèn)為期望雜合度(He)和Shannon信息指數(shù)(I)更為重要,注重物種在種群水平的遺傳多樣性[22],而Hamrick等認(rèn)為多態(tài)位點(diǎn)百分率(P)和期望雜合度(He)比較重要,注重物種水平的多樣性[23]。本研究結(jié)果表明不同種群裸果木平均多態(tài)位點(diǎn)百分率(PPL)為69.93%。種群間He平均值為0.2418;I平均值為0.3629。裸果木總的遺傳多樣性(Ht)為0.3452,而種群內(nèi)的遺傳多樣性(Hs)為0.2489,綜合分析裸果木遺傳多樣性較高,維持著較高的遺傳變異,這與賈舒雯等對(duì)中國(guó)西北干旱區(qū)裸果木遺傳多樣性分析結(jié)果相同[24]。一方面裸果木以有性繁殖為主,世代周期較長(zhǎng),為異花授粉植物,具蟲(chóng)媒及風(fēng)媒傳粉特點(diǎn)[8]。另一方面裸果木為古地中海孑遺種,起源于第三紀(jì),末次間冰期裸果木可能分布范圍較廣,但受冰期的影響使得當(dāng)時(shí)大部分植物消失[25],然而部分遺留下的裸果木在歷史的變遷過(guò)程中保存至今,種群為適應(yīng)環(huán)境的變化,積累豐富了遺傳變異,從而可能使得裸果木存在較高的遺傳多樣性水平,推斷裸果木存在較高的遺傳多樣性可能與其生物學(xué)特性、演化歷史和分布區(qū)生態(tài)因子的特殊性分不開(kāi)。
3.2 裸果木種群遺傳結(jié)構(gòu)與分化特點(diǎn)
種群的遺傳結(jié)構(gòu)是一個(gè)物種最基本的特征,基因流被認(rèn)為是種群遺傳結(jié)構(gòu)均質(zhì)化的主要因素之一。基因流與種群擴(kuò)散、遺傳有效性種群擴(kuò)散有關(guān),具有較高基因流的物種往往比基因流較低物種遺傳分化小[26]。Slatkin認(rèn)為Nm<1,表明種群間差異顯著,基因流不足以抵制種群內(nèi)因遺傳漂變而引起的種群分化,若Nm>1表明種群之間有一定頻率的基因流,遺傳物質(zhì)的交換可以降低種群內(nèi)的遺傳漂變,可以在一定程度上減少種群間的強(qiáng)烈分化[27]。研究發(fā)現(xiàn)裸果木的基因流為1.2920,種群間存在較高的基因流,種群間的遺傳分化小,遺傳變異在種群間均衡分布。馬松梅采用cpDNA標(biāo)記法對(duì)裸果木的遺傳結(jié)構(gòu)進(jìn)行研究表明,中國(guó)西北地區(qū)的干旱和沙漠的擴(kuò)展造成裸果木片段化[28]。本研究發(fā)現(xiàn)引發(fā)裸果木基因流較大的原因可能有以下幾方面:從繁殖生物學(xué)角度看,裸果木繁育系統(tǒng)屬兼性異交類型,異交為主,自交為輔,異交導(dǎo)致裸果木的基因流增大;從傳粉生態(tài)學(xué)角度看,裸果木兼有風(fēng)媒和蟲(chóng)媒傳粉,盛花期裸果木花期開(kāi)花量較大[8],傳粉昆蟲(chóng)較多,使得基因在較多個(gè)體間交流。此外有學(xué)者對(duì)喜馬拉雅紅豆杉(Taxuswallichiana)[29]和桃兒七(Sinopodophyllumhexandrum)[30]的遺傳特性進(jìn)行研究,結(jié)果表明高原隆升形成的主要山系阻礙植物群體間的基因流,促進(jìn)種群間植物異域分化。本研究分布區(qū)位于河西走廊等地,種群沿著祁連山分布,不存在山系間的隔離,各種群間地理距離較近,種群間的地理隔離因子不強(qiáng)烈,存在較大的基因交流。
3.3 地理氣候因子對(duì)裸果木遺傳多樣性的影響
植物種群進(jìn)化過(guò)程中,突變與選擇都是重要的進(jìn)化驅(qū)動(dòng)力,然而一些極端環(huán)境或生態(tài)異質(zhì)較大環(huán)境中,環(huán)境選擇壓力和變異方向尤為重要。地理和環(huán)境因子共同作用可加強(qiáng)植物種群的遺傳分化[31]。如Wu等對(duì)穗狀狐尾藻(Myriophyllumspicatum)遺傳格局進(jìn)行研究發(fā)現(xiàn),區(qū)域尺度主要受環(huán)境要素制約,地理要素顯得更重要[32]。遺傳多樣性與物種多樣性在緯度梯度分布上的耦合暗示著可能存在一些共同的驅(qū)動(dòng)因素[33],導(dǎo)致遺傳多樣性與物種多樣性在緯度梯度上協(xié)同變化。對(duì)荒漠孑遺植物紅砂(Reaumuriasoongarica)的遺傳學(xué)研究表明,自2.96萬(wàn)年以來(lái),亞洲季風(fēng)氣候促進(jìn)了生境片斷化和物種分化,同時(shí)冬季風(fēng)促進(jìn)了物種緯向的基因流[34]。本研究發(fā)現(xiàn)裸果木種群經(jīng)度相對(duì)與緯度對(duì)遺傳多樣性影響較大,樣品采集中經(jīng)度的變化范圍為94°~105°,經(jīng)度跨越較大,可能引發(fā)不同地理間的差異較大,進(jìn)而影響裸果木種群的遺傳多樣性水平。裸果木遺傳與地理距離關(guān)系存在一定的相關(guān)性,從裸果木的分布區(qū)上可以看出,甘肅省阿克塞、甘肅省肅北縣位于裸果木分布的中心地帶,其種群西北的分布主要沿著天山山脈延伸分布于新疆部分區(qū)域,而向東部分布沿著祁連山分布于河西走廊東部及陰山山脈的內(nèi)蒙古西部。從中心區(qū)向外不斷擴(kuò)展使得種群間的基因流變小,遺傳水平隨著地理間距離向兩邊延伸導(dǎo)致遺傳水平降低。全球變化下溫度成為一個(gè)直接的選擇因子,這種選擇壓力也表現(xiàn)在物種的遺傳變異上,即和溫度相關(guān)聯(lián)位點(diǎn)(適應(yīng)性基因)的頻率發(fā)生相關(guān)變化。如Jump等對(duì)不同年齡級(jí)個(gè)體歐洲水青岡(Fagussylcatica)的遺傳多樣性進(jìn)行研究發(fā)現(xiàn),其中一個(gè)位點(diǎn)的頻率變化和溫度有較高的相關(guān)性,表明植物為適應(yīng)環(huán)境而產(chǎn)生變化[35]。Semagn等對(duì)Phytolaccadodecandra遺傳分化研究,發(fā)現(xiàn)種群與溫度、海拔以及生態(tài)地理因素顯著相關(guān),提出遺傳變異是適應(yīng)以上特征的基礎(chǔ)[36]。本研究發(fā)現(xiàn)裸果木遺傳多樣性與海拔及溫度有關(guān),高海拔地區(qū)溫度高,降水低,風(fēng)力較大,在一定程度加速花粉及果實(shí)的散布,加強(qiáng)種群間基因交流,導(dǎo)致遺傳多樣性水平升高。環(huán)境與植物作為一個(gè)整體系統(tǒng),各種因子之間相互聯(lián)系、共同作用,本文僅分析地理及氣候因子與遺傳多樣性的關(guān)系,有關(guān)多因子的綜合作用還有待進(jìn)一步深入探究。
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2024年12期)2024-12-02 00:00:00
英語(yǔ)世界(2023年10期)2023-11-17 09:18:18
中學(xué)生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(shuō)(2021年8期)2021-11-03 10:55:16
學(xué)苑創(chuàng)造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(shù)(快樂(lè)歷史地理)(2020年9期)2020-03-19 05:10:56
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車(chē)觀察(2018年10期)2018-11-06 07:05:26
