新疆地方綿羊ND2基因多態(tài)性及系統(tǒng)發(fā)育分析
2018-08-01 07:53:22王世鋒米耶斯?fàn)?/span>詹建立張秀英王玉濤
江蘇農(nóng)業(yè)科學(xué) 2018年13期
王世鋒, 米耶斯?fàn)枺?詹建立, 張秀英, 王玉濤
(喀什大學(xué)生命與地理科學(xué)學(xué)院葉爾羌綠洲生態(tài)與生物資源研究重點實驗室,新疆喀什 844000)
綿羊與人類生活密切相關(guān),為人類提供了肉、脂、奶、皮、毛、絨、骨和角等多種多樣的畜產(chǎn)品[1-3]。關(guān)于綿羊的起源仍存在爭議,線粒體DNA(mitochondrial DNA,mtDNA)序列分析已經(jīng)確定了一種多母系血統(tǒng)的普遍現(xiàn)象,即A、B、C、D、E進化世系,這些世系有特定的地理范圍,暗示多重母系起源,羊馴化事件可能是獨立的[4]。目前,研究發(fā)現(xiàn)僅歐洲摩弗倫羊可能是家綿羊的母系(世系B)野生祖先之一。推測亞洲摩弗倫羊與歐洲摩弗倫羊與家綿羊共享最近的母系祖先[5],未發(fā)現(xiàn)其他野生羊?qū)揖d羊有遺傳貢獻的分子證據(jù)。
線粒體是真核細胞的動力車間,mtDNA作為獨立與核DNA的半自主性基因組,是核外遺傳物質(zhì),動物體線粒體基因組編碼的37個基因,包括13條多肽,22個tRNA基因和2個rRNA基因[6],具有分子量小、進化速度快、多挎貝、堿基替換率低、遺傳自主及母系遺傳等特征。
ND2基因是mtDNA的1個蛋白編碼基因,而且是NADH脫氫酶的1個亞基,而NADH脫氫酶是呼吸鏈復(fù)合體1的主要組成,在呼吸鏈中直接參與氫與電子傳遞并通過氧化磷酸化產(chǎn)生ATP,進而參與能量代謝作用。NAD參與生物體內(nèi)的重要反應(yīng),NAD可在糖酵解途徑與三羧酸循環(huán)途徑得到電子形成NADH,NADH又在氧化磷酸化途徑中將電子通過呼吸鏈最終傳遞給氧,促成ATP的形成[7]。在體內(nèi)大多數(shù)代謝脫下的氫,是由NADH呼吸鏈傳遞給氧,糖類碳水化合物、脂質(zhì)和蛋白質(zhì)的分解代謝,脫氫后的氧化反應(yīng)一般是由NADH呼吸鏈傳遞完成的。
魯衛(wèi)衛(wèi)等報道雞線粒體ND2基因的異質(zhì)性,并發(fā)現(xiàn)異質(zhì)性變異對生長性狀、屠體指標(biāo)、血清生化等指標(biāo)均有顯著效應(yīng),顯示相關(guān)變異的潛在重要性[7]。鮑海港等在對藏雞的研究中發(fā)現(xiàn),線粒體ND1和ND2基因可被選為藏雞低氧遺傳適應(yīng)的候選基因,因為低氧會抑制與線粒體呼吸功能密切相關(guān)的酶的活性,也會使線粒體產(chǎn)生更多的活性氧。活性氧會破壞膜脂,降低線粒體呼吸酶的活性,從而損害線粒體的呼吸功能。研究發(fā)現(xiàn),3種雞都有但頻率差異較大的2個錯義突變都是ND2基因上的單堿基突變,推測藏雞與低地雞有可能在NADH脫氫酶的功能上存在差異[8]。而有關(guān)高山放牧品種的塔什庫爾干羊(海拔3 000~5 000 m)、和田羊(分為山區(qū)型和農(nóng)區(qū)型,海拔1 440~3 500 m)和多浪羊(海拔1 200 m)線粒體呼吸功能和低氧適應(yīng)研究較少。
本研究通過測定綿羊線粒體ND2基因序列,從而探討新疆南疆地區(qū)綿羊的遺傳多樣性、線粒體呼吸功能及低氧適應(yīng)的分子機制,從而了解并摸清其親緣關(guān)系及低氧適應(yīng)性,為能夠綜合開發(fā)和合理利用綿羊資源提供一定的科學(xué)依據(jù)。
1 材料與方法
1.1 試驗動物
于2014年3月至2015年8月先后在麥蓋提縣種羊場采集33只多浪羊、塔什庫爾干縣麻扎種羊場采集41只塔什庫爾干羊、和田市屠宰場采集17只和田羊共計91只羊的靜脈血液,用檸檬酸葡萄糖(ACD)抗凝,冷凍后置于冰袋中,于實驗室-20 ℃保存?zhèn)溆谩?/p>
1.2 主要試劑
糞便基因組DNA快速提取試劑盒、全血基因組DNA快速提取試劑盒,均購自北京三博遠志生物技術(shù)有限責(zé)任公司;TaqDNA聚合酶、dNTP,均為杭州博日科技有限公司產(chǎn)品;瓊脂糖為BIOWEST公司產(chǎn)品。
1.3 總DNA提取
綿羊總基因組DNA的提取是利用全血基因組DNA快速提取試劑盒(北京三博遠志生物技術(shù)有限責(zé)任公司)。
1.4 PCR擴增及測序
根據(jù)綿羊mtDNA序列(GenBank登錄號為AF010406),采用Primer 5.0軟件設(shè)計引物,擴增綿羊ND2基因,上游引物:5′-AGCACCCACTGATTGCTCAT-3′,下游引物:5′-TTCGTTTTGTGGTTGGGAAT-3′。委托北京三博遠志生物技術(shù)有限責(zé)任公司合成,期望擴增長度約901 bp。
PCR反應(yīng)體系總體積為50 μL:綿羊總基因組4 μL,10×Buffer(含Mg2+)5 μL,上下游引物各2 μL,2.5 mmol/L dNTPs 4 μL,TaqDNA聚合酶1 μL,去離子水29 μL。
PCR反應(yīng)條件:94 ℃預(yù)變性4 min;94 ℃變性30 s,53 ℃退火30 s,72 ℃延伸1 min,進行35個循環(huán);最后72 ℃再延伸10 min,4 ℃保存。
PCR擴增產(chǎn)物直接送北京三博遠志生物公司測序。
1.5 數(shù)據(jù)分析
應(yīng)用Clustal X 1.83軟件進行序列比對并手工校對。DnaSP 5.1軟件計算單倍型多樣度、核苷酸多樣度、多態(tài)位點、核苷酸總變異位點、簡約信息位點和點突變位點等。MEGA 5.1 軟件分析變異位點,計算堿基組成、轉(zhuǎn)換/顛換比(Ts/Tv)、核苷酸差異和序列差異,采用鄰接法(Neighbour-Joining,簡稱NJ)Kimura雙參數(shù)模型構(gòu)建系統(tǒng)發(fā)生樹,對拓撲圖進行重復(fù)抽樣1 000次的自展檢驗(Bootstrap)以確定各分支的置信度。
2 結(jié)果與分析
2.1 ND2基因PCR擴增結(jié)果
ND2基因擴增產(chǎn)物經(jīng)1.5%瓊脂糖凝膠電泳20 min,結(jié)果見圖1。
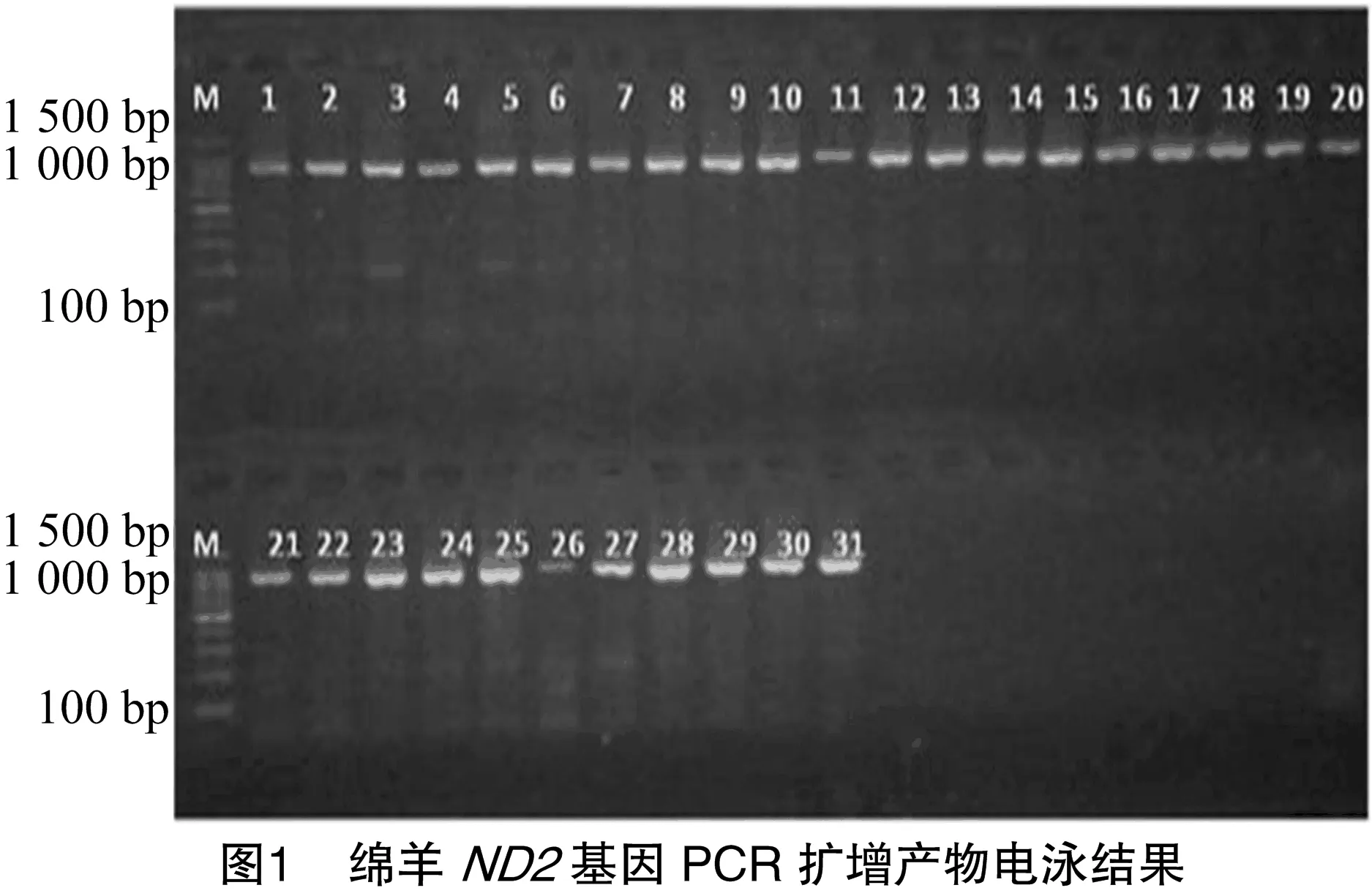
電泳條帶明亮,擴增效果較好,擴增條帶大小為900 bp左右,與預(yù)期擴增的901 bp一致。可以直接測序。
2.2 綿羊mtDNA-ND2全序列堿基分析
測序結(jié)果表明,所測出的綿羊mtDNA-ND2基因A、T、C、G含量在綿羊中分布較一致,其中A占36.8%,T占 27.4%,C占27.1%,G占8.7%。綿羊的A+T含量為 64.2%,C+G含量為35.8%,A+T含量高于C+G含量;表明綿羊mtDNA-ND2基因含有大量的堿基A和T,存在一定的堿基偏倚性。堿基組成表明G相對缺乏,其中密碼子第3位上G的含量最低,僅為2.6%;第3位的A含量較高,為49.8%。密碼子的使用存在一定差異,在密碼子的第1位上A高達43.4%;密碼子第2位上T的含量高達45.0%,堿基G的含量為9.1%。
2.3 綿羊mtDNA-ND2序列的多樣性分析
利用DnaSP 5.1軟件進行單倍型統(tǒng)計分析,本試驗分析91個綿羊mtDNA-ND2基因序列,發(fā)現(xiàn)了22個多態(tài)位點,其中有9個單一多態(tài)位點,簡單信息位點(2個堿基)有13個(圖2)。其中438、504、837、873位點核酸變異只出現(xiàn)在塔什庫爾干羊個體中。核酸變異中轉(zhuǎn)換遠大于顛換,轉(zhuǎn)換/顛換比為78.38。

2.4 ND2氨基酸分析
22個多態(tài)位點中的2個位點即299、883位點是非同義突變,分別導(dǎo)致M變?yōu)門和R變?yōu)镚,其余20個位點為同義突變。應(yīng)用SWISS-MODEL(httP://swissmodel.expasy.org/interactive# structure)軟件進行ND2結(jié)構(gòu)在線預(yù)測,多肽空間結(jié)構(gòu)見圖3。
單倍型H1序列未發(fā)生突變,H8序列在299位點和883位點均發(fā)生突變。由圖3可知,突變后空間結(jié)構(gòu)差異不大。但是QMEAN分別為-4.35和-4.38,Cβ分別為-1.72和 -1.82,All Atom分別為-1.27和-13.2,Torsion分別為 -4.32 和-4.33,Solvation均為1.47,未發(fā)生變化。

2.5 mtDNA-ND2基因單倍型分析
共獲得13個單倍型,各單倍型在各品種中分布見表1。
由表1可知,H2和H1為優(yōu)勢單倍型,分別占58.3%和13.2%。綿羊群體單倍型多樣度(Hd)為0.641,核苷酸多樣度(Pi)為:0.357%,平均核苷酸差異數(shù)(k)為3.4。塔什庫爾干羊、和田羊和多浪羊核苷酸多樣度分別為0.330%、0.135%、0.463%。
2.6 綿羊系統(tǒng)進化分析
利用MEGA 5.05軟件構(gòu)建綿羊ND2基因不同單倍型NJ系統(tǒng)發(fā)育樹(圖4)。其中包括GenBank下載的36個序列分別是A世系(HM236174-5)、B世系(AM236176-7)、C世系(AM236178-9)、D世系(HM236180-1)、E世系(HM236182-3)序列、盤羊線粒體序列(藏盤羊JX101654.1、帕米爾盤羊KT781689.1、Darwini盤羊KX609626.1、東方盤羊NC020656.1、HM236188.1、加拿大盤羊NC015889.1、JN181255.1)、Vignei(HM236186.1、HM236187.1、HM236189.1、NC026064.1、KF938361)、摩弗倫羊(HM236185、KF312238.1、KF938360.1、NC026063.1)、新疆地方綿羊阿勒泰羊(KF938320.1)、和田羊(KF938322.1)、吐魯番黑羊(KF938324.1)、巴什拜羊(KF938330.1)巴音布魯克羊(KF938331.1)、多浪羊(KF938332.1)、哈薩克羊(KF938333.1)、塔什庫爾干羊(KF938337.1)、葉城羊(KF938338.1)。
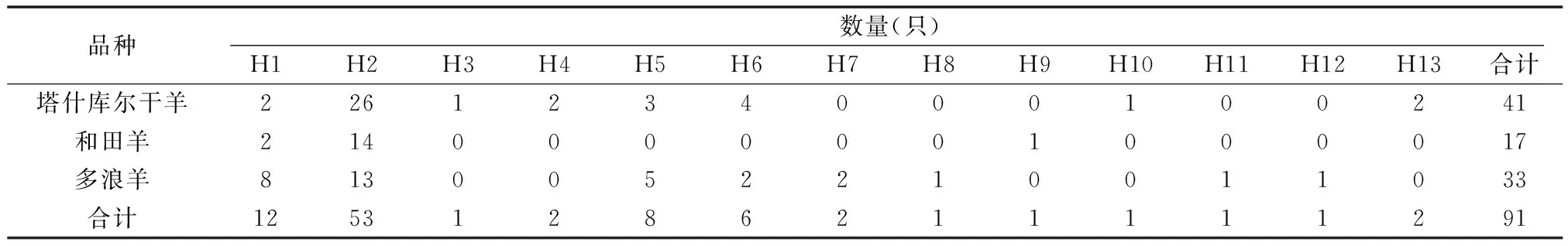
表1 各單倍型在各品種中的分布
由圖4可知,單倍型H2、H3、H9、H10、H11和H13屬于A世系;單倍型H1、H4和H7與歐洲摩弗侖羊和亞洲摩弗倫羊?qū)儆贐世系;H5、H6和H8與塞浦路斯摩弗倫羊?qū)儆贑世系;H12屬D世系。

2.7 3個品種間的遺傳距離及系統(tǒng)發(fā)生樹
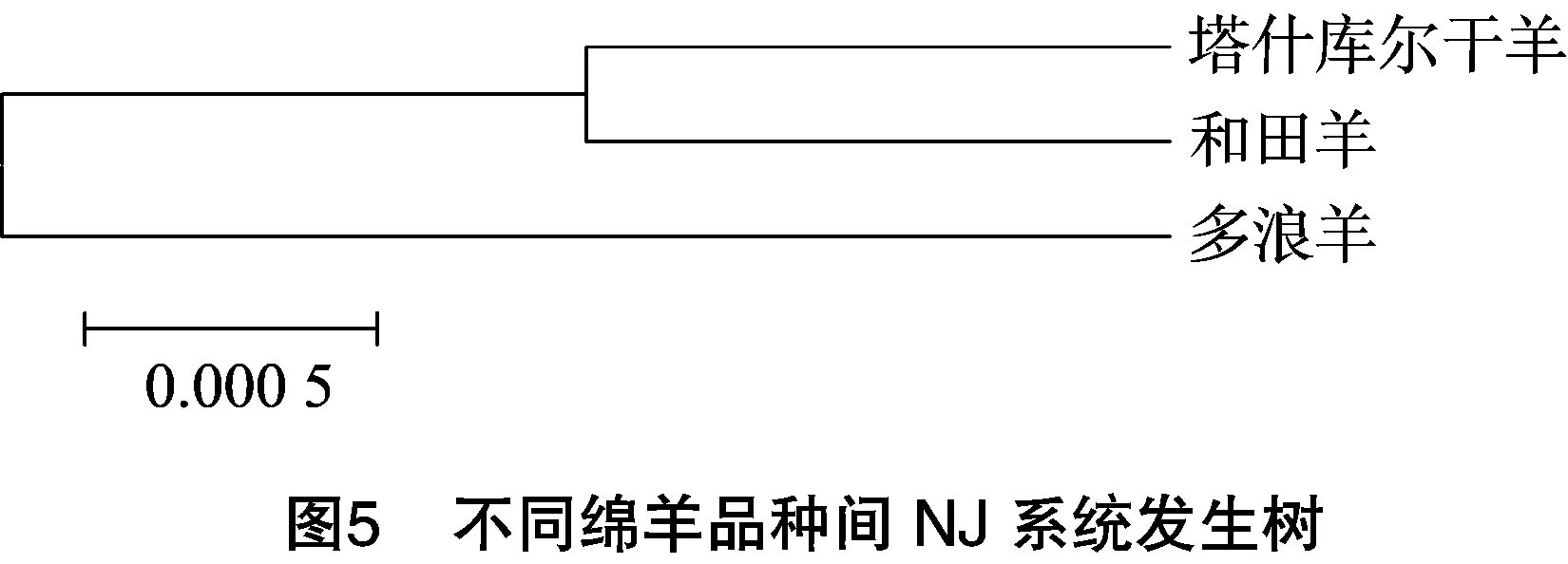
利用MEGA 5.0計算不同品種間遺傳距離(表2),根據(jù)遺傳距離構(gòu)建不同品種間NJ系統(tǒng)發(fā)生樹(圖5)。由圖5可知,塔什庫爾干羊與和田羊關(guān)系較近,其次是多浪羊。
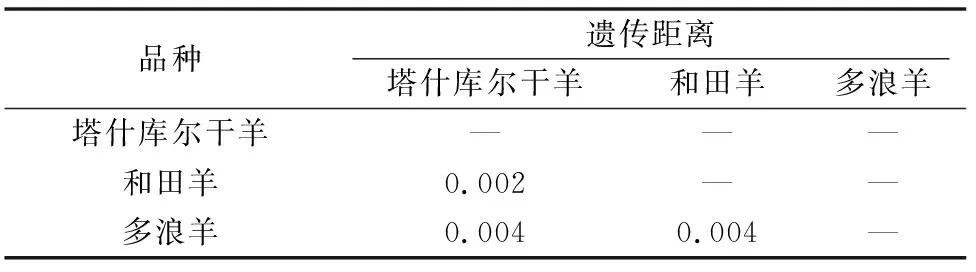
表2 3個品種間的遺傳距離
3 討論
綿羊的A+T含量為64.2%,C+G含量為35.8%,A+T含量高于C+G含量;表明綿羊mtDNA-ND2基因含有大量的堿基A和T,存在一定的堿基偏倚性,與筆者前期研究結(jié)果[9-10]一致。
基因變異發(fā)生在密碼子第1、2、3位置分別占9.09%、4.55%、86.36%,與前期研究結(jié)果[9]一致。
轉(zhuǎn)換顛換比78.38遠大于36.81[9]和轉(zhuǎn)換顛換比的臨界值2,說明新疆南疆地區(qū)地方綿羊ND2基因序列突變可能未達到飽和狀態(tài),轉(zhuǎn)換隨著遺傳變異增加趨于飽和。
馬麗娜等在蒙古羊和小尾寒羊上發(fā)現(xiàn)770位點上發(fā)生了T到C的轉(zhuǎn)換[11],本研究未發(fā)現(xiàn)此變異。
438、504、837、873位點核酸變異只出現(xiàn)在塔什庫爾干羊個體中,單倍型H3、H4、H10和H13也只分布于塔什庫爾干羊中,是否與其NADH脫氫酶功能差異和低氧適應(yīng)有聯(lián)系,仍需進一步研究。另外單倍型H8序列在299位點和883位點均發(fā)生突變。突變后ND2亞基空間結(jié)構(gòu)差異不大,但是多肽部分性質(zhì)發(fā)生改變,是否影響線粒體呼吸功能有待進一步探討。
單倍型H12只分布于多浪羊中,與D世系聚在一起,表明多浪羊存在世系D,與基于細胞色素b的分析結(jié)果[9]一致。和田羊未發(fā)現(xiàn)世系C,可能與研究樣品數(shù)量偏少有關(guān)。
3個品種平均核苷酸多樣度(Pi)為0.357%,塔什庫爾干羊、和田羊和多浪羊核苷酸多樣度分別為0.330%、0.135%、0.463%,低于趙倩君等[12]、王昕等[13]、張傳生等[14]基于細胞色素b研究的0.805%、0.602%、0.850%。但與筆者基于細胞色素C氧化酶亞基Ⅰ(COⅠ)基因分析塔什庫爾干羊的0.421%接近。
基于ND2基因的系統(tǒng)發(fā)生樹表明摩弗倫羊可能是家綿羊世系B的野生祖先,與前人研究觀點[5]一致。但是塞浦路斯摩弗倫羊與是世系C聚在一起,可能對世系C有遺傳貢獻,此觀點有待進一步驗證。另外塔什庫爾干羊與和田羊關(guān)系較近,其次是多浪羊,此觀點與湯存?zhèn)サ然谖⑿l(wèi)星的分析結(jié)果[15]一致。
