外源水楊酸甲酯對高溫脅迫下茶樹光合作用和抗氧化酶的影響
2018-08-16 08:35:04魏吉鵬李鑫王朝陽李洋張蘭沈晨顏鵬張麗平韓文炎
茶葉科學 2018年4期
魏吉鵬,李鑫*,王朝陽,李洋,3,張蘭,沈晨,顏鵬,張麗平,韓文炎*
?
外源水楊酸甲酯對高溫脅迫下茶樹光合作用和抗氧化酶的影響
魏吉鵬1,李鑫1*,王朝陽2,李洋1,3,張蘭1,沈晨1,顏鵬1,張麗平1,韓文炎1*
1. 中國農業科學院茶葉研究所,浙江 杭州 310008;2. 安康市農業科學研究院,陜西 安康 725000; 3. 河北農業大學園藝學院,河北 保定 071000
近年來茶園高溫災害頻發,而關于提高茶樹耐熱性的研究相對較少。本文以龍井43為試驗材料,利用不同濃度水楊酸甲酯(MeSA)噴施茶苗后,在高溫環境下(43℃)處理12?h,隨后測定茶樹葉片的凈光合速率(Pn),Rubisco最大羧化速率(Vmax)、RuBP最大再生速率(max),電解質滲透率(EL),丙二醛(MDA)含量以及抗氧化酶活性等相關指標。結果發現,1?mmol·L-1MeSA能夠有效緩解高溫導致的茶樹Pn降低,維持Vmax和max穩定;高溫導致茶樹葉片EL和MDA含量迅速上升,而適當濃度的MeSA可顯著降低高溫環境下EL和MDA含量。此外,結果表明1?mmol·L-1MeSA能夠提高APX和CAT活性,進而減少H2O2積累,減輕茶樹細胞膜過氧化作用。綜上所述,外源施用MeSA能夠在一定程度上維持高溫條件下植物葉片細胞光合系統穩定,提高茶樹葉片的抗氧化酶活性,緩解氧化脅迫,最終提高茶樹的耐熱性。
水楊酸甲酯;茶樹;高溫脅迫;光合作用;抗氧化酶
茶是世界三大無酒精飲料之一,中國是第一產茶大國[1]。茶樹的生長發育對氣溫和水分要求相對嚴格。在全球氣候變暖的背景下,我國產茶區夏季極端高溫、干旱等事件愈來愈頻繁。高溫干旱影響茶樹正常的生長發育,茶樹葉面溫度過高容易導致氣孔關閉,光合能力下降,葉綠素降解,葉片功能喪失[2];嚴重時導致茶葉萎焉,茶葉品質下降,甚至最終導致茶樹死亡,給茶葉生產帶來巨大的損失[3]。因此研究茶樹應對高溫脅迫的生理機制,進而尋找方法提高茶樹的耐熱性具有重要的科研價值和社會經濟價值。
水楊酸(Salicylic acid,SA)和水楊酸甲酯(Methyl salicylate,MeSA)是植物界普遍存在的酚類化合物,SA易于轉化為MeSA,兩者的生理功能基本一致[4]。研究發現SA在植物氣孔開閉、種子萌發、開花、果實生長等生長發育過程中起著重要作用[5]。20世紀90年代,研究人員發現SA同樣參與植物應對逆境脅迫的生理過程,SA開始作為植物應對脅迫反應的一種信號分子來研究[6]。目前已有關于SA參與植物抗病、抗鹽、抗熱、抗旱、抗寒等生理過程的研究報道[7]。
目前的研究發現,MeSA預處理能夠修復高溫導致的植物葉片光合機構的損壞,維持PSⅡ反應中心結構的穩定,從而在一定程度上維持高溫下植物光合作用的持續進行[8]。同時使用MeSA預處理還能提高高溫下植物葉片抗氧化酶活性,降低MDA含量及細胞膜過氧化程度,減輕高溫強光脅迫導致的氧化損傷[9]。在茶樹方面,盡管學術界已經開展了SA參與茶樹抗蟲性[10],抗寒性[11],抗鹽性[12]等研究,但是關于SA對茶樹抗熱性影響的研究尚未深入開展,尤其是SA對于高溫脅迫下茶樹葉片光合作用和抗氧化酶的影響還沒有研究報道。基于此,本研究選用了龍井43茶苗,通過噴施不同濃度MeSA進行預處理,然后進行高溫處理,隨后測定茶樹光合以及抗氧化酶活性等生理指標,從而探明MeSA對高溫脅迫下茶樹光合作用和抗氧化酶的影響。
1 材料與方法
1.1 試驗材料與處理
試驗在中國農業科學院茶葉研究所內進行,試驗材料為龍井43茶樹品種。試驗選擇生長狀態一致,枝條健壯,無病蟲害侵襲的盆栽苗待用。水楊酸甲酯(MeSA)采購自美國Aladdin公司,分別配置成不同濃度(0、0.1、1、5?mmol·L-1)。
在不同濃度的MeSA溶液中加入0.3‰的有機硅后,將其噴施在茶樹葉片上,直至葉片滴水為止。隨后將噴施不同濃度MeSA的盆栽茶苗均分,分別放入兩個不同溫度環境的人工氣候培養箱中。其中常溫處理設置為28℃,高溫處理設置43℃。除溫度不同外,人工氣候箱平均光強均設定為600?μmol·m-2·s-1,光周期為14?h/10?h(晝/夜),相對濕度均控制在80%,CO2體積分數設置為0.04%。連續處理12?h后,測定茶樹葉片的光合氣體交換參數;隨后取一芽及半展葉用于基因表達分析;取一芽二葉樣品稱重后置于液氮中用于后續生理指標的測定。每個處理3個生物學重復。
1.2 測定項目與方法
1.2.1 光合氣體交換測定
利用LI-COR 6400型光合儀(美國LI-COR公司),在光強為600?μmol·m-2·s-1,CO2體積分數為0.04%的條件下測定茶樹頂芽向下第三葉片的凈光合速率(Pn)。參考Voncaemmerer等[13]的方法測定光合作用CO2響應曲線(A/Ci)后,使用Ethier等[14]的方法計算出Rubisco最大羧化速率(Vmax)以及Rubisco的最大再生速率(max)。每個處理設3個重復。
1.2.2 電導率(EL)的測定
準確稱取試樣0.1?g裝入試管,加蒸餾水20?mL,抽氣泵真空抽氣1?h,隨后振蕩混勻并室溫靜置2?h。靜置結束后再次振蕩混勻,隨后用EC215型數字電導率儀測第1次電導率值(1),然后沸水浴1?h,冷卻至室溫后測第2次電導率值(2)。
細胞傷害率=1/2×100%。
1和2需轉化為25℃時的電導率(25)。
25=t[1 +0.02(-25)]式中,t為實測電導率,為實測溫度。
1.2.3 MDA含量測定
采用硫代巴比妥酸法(TBA)法,取茶樹葉片組織研磨后取上清液加入10%三氯乙酸(TCA)和0.65%硫代巴比妥酸(TBA)在95℃加熱30?min,測定OD532和OD600,計算MDA含量。
1.2.4 H2O2含量測定和DAB染色
H2O2含量測定:根據Okuda T等[15]的方法,取葉片組織0.3?g加入3?mL HClO4研磨后離心,用KOH將pH調至6~7。用活性炭吸附色素后,離心取上清液過濾膜,隨后根據Willekens H等[16]的方法測定濾液中H2O2的含量。
DAB染色:配制0.1%二甲基聯苯胺(DAB)溶液(含50?mmol·L-1Tris-HCl,pH3.8)后,取茶樹自頂芽向下第三片葉片放入其中,25℃條件下光照培養直到葉片上出現深色斑點。隨后將葉片放入95%的乙醇中,100℃水浴煮沸15?min,重復這一步驟直到葉片葉綠素完全脫去,進行拍照保存。
1.2.5 APX和CAT活性測定
參考Wang等[17]的方法提取粗酶液,并加以改進。取茶樹葉片組織0.3?g加入3?mL 50?mmol·L-1(含0.2?mmol·L-1EDTA)pH7.8緩沖液研磨。將勻漿轉入10?mL的離心管中,12?000低溫下離心20?min,上清液即為粗酶液。
抗壞血酸過氧化物酶(APX)的活性測定參考Nakano等[18]的方法,并加以改進。取100?μL酶液,加入1?700?μL 25?mmol·L-1PBS(pH7.0,含有0.1?mmol·L-1EDTA),100?μL 20?mmol·L-1H2O2,100?μL 5?mmol·L-1ASA(抗壞血酸)將反應液混合后,測定40?s內OD290的動力學變化,并計算酶活。
過氧化氫酶(CAT)活力測定參照Patra等[19]的方法,并加以改進。取100?μL酶液,加入1?700?μL 25?mmol·L-1PBS(pH7.0,含0.1?mmol·L-1EDTA),200?μL 100?mmol·L-1H2O2。將反應液混合后,立刻測定40?s內OD240的動力學變化,并計算酶活。
1.3 數據分析與處理
試驗中各指標均重復測定3次,取其平均值。數據圖片用Origin Pro 8.0繪制(Microcal軟件公司,美國),并采用SAS 9.0進行差異顯著性分析。
2 結果與分析
2.1 MeSA對高溫條件下茶樹葉片光合作用的影響
高溫脅迫會導致茶樹光合系統受到破壞,光合酶活性喪失,光合能力下降。凈光合速率能夠在一定程度上反應光合能力。如圖1所示,相對于高溫條件,常溫條件下茶樹光合能力維持在較高水平,各處理組凈光合速率(Pn)較高;且常溫條件下不同濃度的MeSA處理對茶樹光合作用沒有顯著影響。在高溫條件下,對照組未使用MeSA處理,其Pn降低了49.89%;而MeSA預處理能夠緩解高溫條件下Pn的下降。高溫條件下0.1、1.0、5.0?mmol·L-1的MeSA預處理的茶樹葉片Pn值比對照分別提高了25.25%、60.58%、和36.02%。其中1?mmol·L-1MeSA能夠最大程度緩解高溫條件下茶樹葉片Pn的抑制作用,與對照組相比效果顯著。此外,研究還發現隨著MeSA濃度的提高,其作用效果并未呈線性增強,高溫條件下施用5.0?mmol·L-1MeSA的茶樹葉片Pn只比對照提高了36.02%,緩解效果弱于1.0?mmol·L-1MeSA處理組。

注:圖中不同字母代表不同處理間差異顯著,P<0.05。下同。
2.2 MeSA對高溫條件下茶樹葉片Rubisco的影響
Rubisco是光合作用C3碳反應中重要的羧化酶,也是光呼吸中不可缺少的加氧酶,是光合作用中決定碳同化速率的關鍵酶。在卡爾文循環中,Rubisco的羧化速率(Vmax)和RuBP的再生速率(max)直接影響到暗反應有機物的積累速率。高溫會破壞Rubisco的蛋白結構,導致酶活性下降。如圖2所示,常溫條件下不同濃度MeSA處理對茶樹葉片V,max和max沒有顯著影響;高溫條件下,對照組Vmax和max分別降低了31.78%和38.32%,使用0.1、1.0、5.0?mmol·L-1的MeSA預處理的茶樹葉片Vmax值比對照組分別提高了7.14%、23.93%、和20.35%,max值比對照組分別提高了12.16%、38.93%、和19.63%。盡管使用MeSA處理均能減少茶樹在高溫條件下V,max值的降低,其中1.0?mmol·L-1MeSA效果最為明顯。同樣的,使用1.0?mmol·L-1的MeSA能夠最大程度的緩解高溫條件下茶樹葉片max值的下降,而0.1?mmol·L-1和5?mmol·L-1的MeSA緩解效果緊隨其后,兩者之間無明顯差異。
2.3 MeSA對高溫條件下茶樹葉片電解質滲透率(EL)和丙二醛(MDA)含量影響
植物細胞的EL和MDA含量均與細胞膜的完整性相關。如圖3所示,在常溫條件下,不同濃度的MeSA處理對茶樹葉片組織的EL和MDA無顯著影響。高溫處理后,茶樹葉片EL值迅速提高,對照組EL提高了2.86倍,達到了28.62%。而施用MeSA能夠在一定程度上降低高溫條件下的EL,0.1、1.0、5.0?mmol·L-1的MeSA預處理的茶樹葉片EL比對照組分別降低了20.72%、33.69%、和38.97%。高溫處理同樣導致茶樹葉片MDA含量升高,高溫下對照組MDA含量提高了1.82倍,達到212.69?μmol·g-1。使用0.1、1.0、5.0?mmol·L-1的MeSA預處理的茶樹葉片MDA含量比對照組分別降低了9.88%、25.91%、和19.09%。同樣1.0?mmol·L-1的MeSA預處理能夠最大程度減少高溫條件下茶樹葉片MDA含量,當濃度達到5.0?mmol·L-1時,其效果有所減弱。
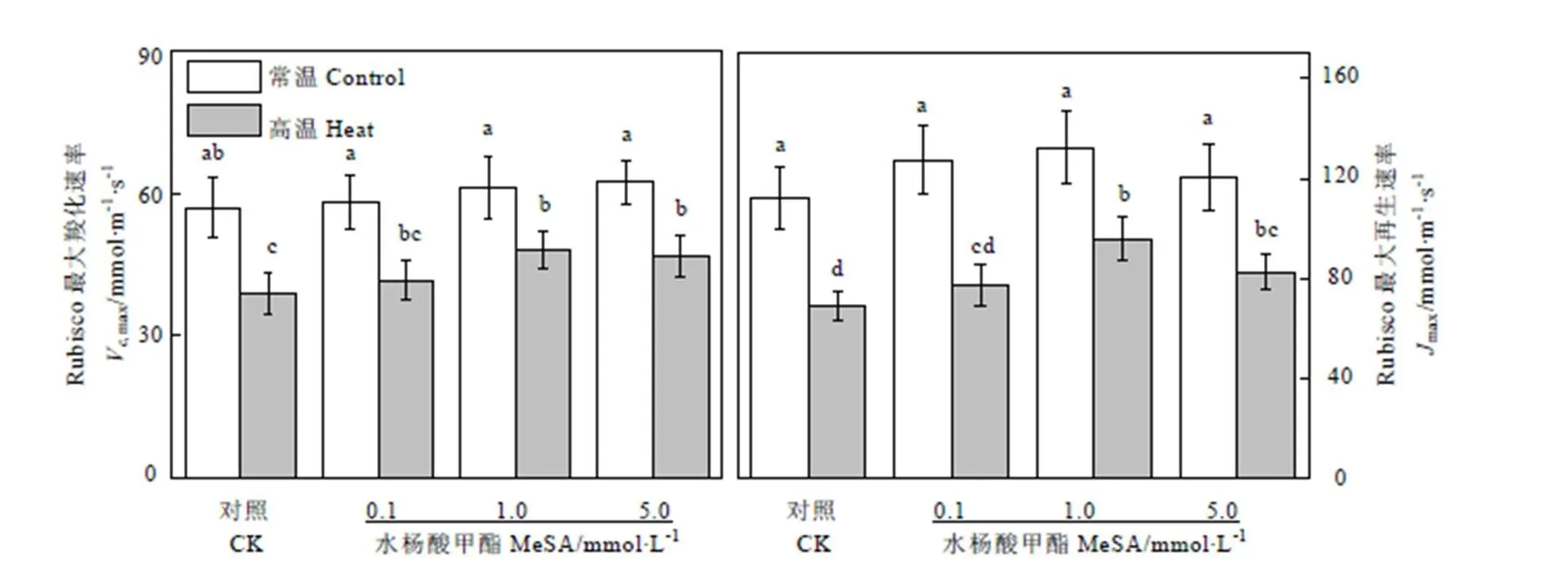
圖2 MeSA處理對高溫脅迫下茶樹Vc,max和Jmax的影響
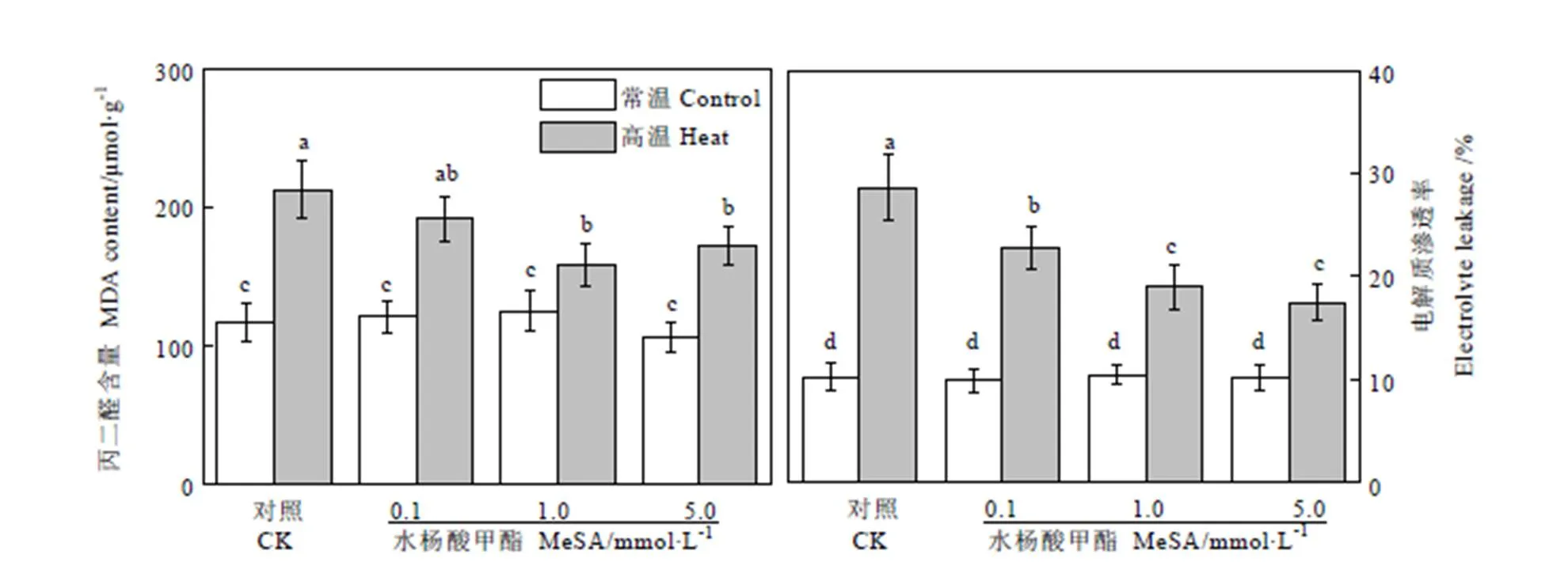
圖3 MeSA處理對高溫脅迫下茶樹MDA和EL的影響
2.4 MeSA對高溫條件下茶樹葉片H2O2含量影響
H2O2是植物細胞在代謝過程中產生的一種活性氧分子,通過DAB染色后,組織細胞內有H2O2的部位生成金黃色的顆粒物。如圖4-A DAB染色所示,在常溫條件下茶樹自頂芽向下第三葉片無明顯的顆粒物,葉片顏色較均勻,呈黃色;在高溫條件下,對照組葉片出現大量顆粒物沉淀,這些顆粒聚集后,葉片變為黃褐色。與對照組相比,高溫條件下使用MeSA預處理的茶樹葉片顆粒物均有所減少。從圖中可以看出,高溫條件下1.0?mmol·L-1MeSA處理的葉片顆粒物最少,5.0?mmol·L-1與之效果相似,0.1?mmol·L-1次之。
通過生物化學手段測定的H2O2含量與染色結果趨勢一致。如圖4-B所示,常溫條件下茶樹葉片組織內H2O2含量較低,噴施不同濃度MeSA的茶樹葉片H2O2含量雖然有所差別,但基本維持在58~87?nmol·g-1之間。常溫條件下對照組H2O2含量最低,為58.18?nmol·g-1,使用1.0?mmol·L-1MeSA處理的葉片H2O2生成量最高,達到86.35?nmol·g-1。在高溫條件下,不同濃度MeSA處理茶樹葉片H2O2含量升高到142~193?nmol·g-1之間。對照組在不使用MeSA處理的情況下,茶樹葉片H2O2含量最高,達到了192.31?mol·g-1,為常溫條件下的3.3倍。隨著MeSA濃度升高,H2O2含量逐漸下降。使用0.1、1.0、5.0?mmol·L-1MeSA處理的茶樹葉片H2O2生成量分別比對照組減少了7.16%、21.54%和26.02%。
2.5 MeSA對高溫條件下茶樹葉片抗氧化酶活性的影響
抗壞血酸過氧化物酶(APX)和過氧化氫酶(CAT)是植物組織細胞內活性氧(ROS)清除系統的重要組成部分,兩者均能與活性氧分子反應。如圖5所示,與高溫條件相比,常溫下APX和CAT活性較低。在常溫條件下,當MeSA濃度為1?mmol·L-1時,APX活性比對照組提高了24.68%,5?mmol·L-1MeSA處理有相似效果,而0.1?mmol·L-1MeSA處理組茶樹葉片APX活性與對照組無明顯差異。在高溫條件下,各處理組APX活性均有提高。高溫下對照組APX活性比常溫條件下提高了43.01%;使用MeSA處理能夠進一步提高茶樹葉片中APX活性,使用0.1、1、5?mmol·L-1MeSA處理的茶樹葉片APX活性分別比高溫處理下對照組分別提高了12.64%、28.98%和11.83%,由此可見1?mmol·L-1MeSA能夠最有效提高高溫下茶樹葉片APX的活性。
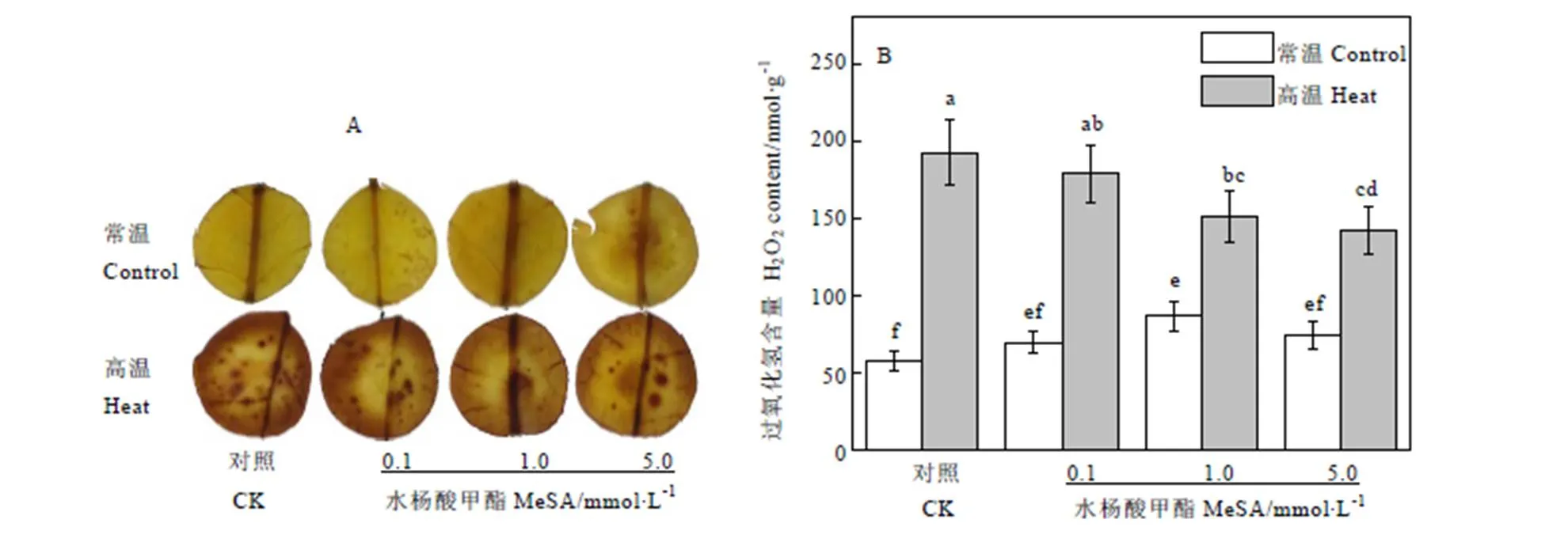
圖4 MeSA處理對高溫脅迫下茶樹H2O2含量的影響

圖5 MeSA處理對高溫脅迫下茶樹APX和CAT的影響
與APX類似,常溫條件下使用MeSA處理的茶樹葉片CAT的活性與對照組相比均有所增加,其中1?mmol·L-1MeSA處理效果最為顯著。高溫下茶樹APX活性升高,對照組茶樹葉片APX的活性在高溫條件下達到0.24?μmol·mg-1·min-1,是常溫下的1.52倍。高溫條件下使用0.1、1.0、5.0?mmol·L-1MeSA處理的茶樹葉片APX的活性分別比對照組提高了13.61%、40.33%和8.54%。
3 討論
李治鑫等[20]的研究結果表明,持續的高溫導致茶樹葉片顏色改變,甚至枯萎卷曲。將茶樹置于43℃條件下持續48?h后,茶樹葉片葉綠素逐漸降解,茶樹葉片的Fv/Fm呈現明顯的下降趨勢,ΦPSⅡ明顯降低,綜合表現為茶樹葉片光合能力持續下降。而SA能夠提高植物光合作用的研究早有報道。Ananiera等[21]使用大麥苗作為研究材料,發現500?μmol·L-1的SA能提高大麥葉片葉綠素含量與氣孔導度,從而提高麥苗的Pn。王利軍等[22]研究發現SA能夠在一定程度上維持高溫脅迫下溫洲蜜柑葉片氣孔導度,減少Fv/Fm的下降,進而減少Pn值的下降。同樣在本研究中高溫脅迫下不施用MeSA的茶樹Pn比常溫條件下的降低了49.89%,這與李治鑫等的研究結果一致。在使用不同濃度的MeSA處理后,高溫條件下茶樹葉片Pn值均有所提高,且當濃度達到1.0?mmol·L-1時,效果最為明顯。這說明SA對高溫脅迫下茶樹光合機構也能發揮一定的保護作用。
高溫脅迫下植物光合作用的降低,現有的研究認為主要由氣孔因素和非氣孔因素導致[23]。氣孔因素主要包括高溫導致葉片氣孔關閉,光合葉片氣孔導度(Gs)下降,使葉綠體內CO2的供應受阻。而在長期的高溫脅迫過程中,非氣孔因素才是導致植物光合效率下降的主要原因[24]。非氣孔因素主要包括光合相關酶活性降低和光合器官損傷。Rubisco是碳同化代謝的關鍵酶,其初始活力是決定植物光合碳反應能力最直接的內在因素[25]。高溫主要是通過影響Rubisco活性來影響光合作用暗反應的進行。研究發現隨著溫度的升高,Rubisco對CO2的親合性會逐漸減弱,Vmax下降,導致Rubisco更傾向于加氧反應。隨著光呼吸速率增高,光合作用強度減弱,影響了有機物的積累[26]。本研究中使用MeSA處理后,高溫下Rubisco的Vmax和max均有所提升,說明SA能夠在一定程度上緩解高溫對Rubisco活性的抑制,穩定高溫下Rubisco對CO2的親合性,抑制光呼吸,促進有機物的生成,從而增強茶樹在高溫條件下的光合作用。
植物細胞膜的完整性對維持細胞的微環境和正常的代謝起著重要的作用。在常溫條件下細胞膜對物質具有選擇透性能力,植物細胞內電解質含量保持穩定。研究發現當植物遭受包括高溫在內的逆境脅迫時,植物細胞膜脂容易與活性分子發生過氧化作用,生成大量的MDA[27]。同時MDA還能與細胞膜上結構物質反應,進一步破壞細胞膜的完整性。在高溫脅迫下植物細胞膜發生過氧化作用后,細胞膜結構完整性受到破壞,膜透性增大,導致細胞內的電解質外滲,以致植物細胞浸提液的電導率增大[28]。細胞膜完整性越差,膜內電解質外滲越多,電導率越大。因此,MDA含量和電導率的大小能夠在一定程度上反映出細胞膜的完整程度。王開凍等[29]研究發現,使用100?μmol·L-1的SA溶液能夠有效降低高溫條件下南瓜幼苗的EL和MDA含量。呂俊等[30]研究發現0.5?mmol·L-1的SA預處理水稻幼苗,能夠降低其在高溫脅迫下的電導率和MDA含量,說明SA預處理能夠降低膜脂過氧化程度,減緩水稻幼苗電解質的滲出。本研究的試驗結果與前人研究相似,噴施不同濃度的MeSA均能有效降低高溫脅迫下茶樹電導率和MDA含量。說明MeSA能夠在一定程度上穩定高溫條件下茶樹細胞膜脂結構,減少MDA的生成,從而在一定程度上維持了細胞膜的完整性,降低了高溫脅迫下茶樹電導率。而茶樹細胞膜的完整,是進行正常生理代謝的基礎。
植物細胞在高溫脅迫下會產生過量的H2O2等活性氧類(ROS)物質。過量的H2O2能與植物體內的蛋白質、膜質、DNA及其他細胞組分發生反應,對植物細胞造成傷害,影響生理功能[31]。因此通過降低高溫條件下植物H2O2的含量能夠提高植物在高溫條件下的抗性。Dat等[32]研究發現使用SA處理能夠降低芥菜(L)幼苗在高溫條件下H2O2的累積量,并顯著改善高溫下幼苗的生長發育狀況。本研究通過生化手段測定了高溫脅迫下茶樹H2O2的累積量,發現高溫同樣導致茶樹產生過量的H2O2。研究發現使用MeSA預處理能夠降低高溫下H2O2的累積量,這很有可能是通過增強茶樹體內的抗氧化系統來實現的。
當植物在高溫下產生過量的ROS分子時,植物體會通過抗氧化系統來清除過量的ROS分子[33]。植物體內主要有兩類抗氧化防御系統,即酶類抗氧化和非酶類抗氧化防御系統,其中酶類抗氧化防御系統包括APX和CAT在內的一系列抗氧化酶[34]。APX通過催化抗壞血酸與H2O2分子反應,從而消除過量的H2O2分子,達到保護細胞的作用[35];CAT是一種以鐵卟啉為輔基的酶類清除劑,它能夠促進H2O2迅速分解為分子氧和水,約占過氧化物酶體40%[16]。Fan等[36]研究發現,外源施用SA后,將榆樹置于35℃的高溫條件下,分析差異表達基因,發現SA能夠上調控制APX和CAT合成相關的基因,促進高溫條件下兩者的生物合成。杜朝昆等[37]研究發現,使用300?mmol·L-1SA溶液預處理的玉米幼苗,能夠提高其體內APX和的CAT活性,從而提高玉米幼苗對高溫抗性。而在本研究中,我們發現通過噴施1?mmol·L-1的MeSA能夠顯著提高茶樹體內APX和CAT的酶活,從而提高茶樹在高溫下對H2O2的清除能力,由此進一步提高茶樹對高溫的抗性。
在本研究中,我們使用0.1、1、5?mmol·L-1的MeSA處理茶樹后進行高溫處理,隨后檢測了各項生理生化指標,通過綜合評價我們認為1?mmol·L-1的MeSA能夠最為有效維持高溫條件下茶樹葉片內部Rubisco的活性,提高高溫條件下Vmax和max。由此在一定程度上維持茶樹在高溫條件下暗反應的順利進行,從而減少茶樹Pn的下降。同時通過研究發現,SA能夠通過提高包括APX和CAT在內的抗氧化酶活性,提高高溫條件下茶樹清除H2O2的能力。這在一定程度上減少了ROS對細胞膜的破壞,減輕膜脂過氧化作用,進而降低高溫條件下茶樹MDA的生成,緩解葉片組織El的升高。因此MeSA能夠在一定程度上維持高溫條件下茶樹細胞膜完整性,維持茶樹葉片細胞正常代謝,從而提高茶樹對高溫的抗性。
盡管MeSA能夠提高茶樹對高溫的抗性,但是具體的信號途徑尚不清晰。Slaymaker等[38]研究煙草發現SA能與靶細胞膜上一種受體結合蛋白(SABP)結合,隨后影響靶細胞內H2O2含量和Ca2+含量[39]。H2O2和Ca2+作為信號分子進而介導下游的一系列抗性反應。但是這一介導過程的信號途徑尚未明確。而Horváth等[40]研究發現,過高濃度的SA本身導致植物大量合成ROS分子,會使植物產生氧化脅迫,反而削弱了植物對于高溫等逆境脅迫的抗性。這也可能是導致在本研究中5?mmol·L-1的MeSA處理效果反而不如1?mmol·L-1的MeSA效果明顯的原因。由于SA在茶樹體內的作用機制研究并不透徹,關于其在茶樹高溫抗性上的研究相對較少,因此相關問題亟需進一步研究。
[1] 梅峰. 國際茶市現狀與中國茶產業發展[J]. 中國茶葉加工, 2009(1): 3-6.
[2] 韓冬, 楊菲, 楊再強, 等. 高溫對茶樹葉片光合及抗逆特性的影響和恢復[J]. 中國農業氣象, 2016, 37(3): 297-306.
[3] 何辰宇, 李蓓蓓, 楊菲. 高溫干旱對茶葉生產的影響及應對措施[J]. 江蘇農業科學, 2016, 44(4): 215-217.
[4] Paterson J R, Srivastava R, Baxter G J, et al. Salicylic acid content of spices and its implications [J]. Journal of Agricultural & Food Chemistry, 2006, 54(8): 2891-2896.
[5] Klerk G J D, Marinova S, Rouf S, et al. Salicylic acid affects rooting of apple microcuttings by enhancement of oxidation of auxin [J]. Acta Horticulturae, 1997, 447(447): 247-248.
[6] Ogawa D, Nakajima N, Sano T, et al. Regulation of salicylic acid synthesis in ozone-exposed tobacco and[J]. Phyton, 2005, 45(4): 169-175.
[7] 王利軍, 戰吉成, 黃衛東. 水楊酸與植物抗逆性[J]. 植物生理學報, 2002, 38(6): 619-624.
[8] An C F, Mou Z L. Salicylic acid and it’s function in plant immunity [J]. Journal of Integrative Plant Biology, 2011, 53(6): 412-428.
[9] Wang L J, Li S H. Salicylic acid-induced heat or cold tolerance in relation to Ca2+homeostasis and antioxidant systems in young grape plants [J]. Plant Science, 2006, 170(4): 685-694.
[10] 苗進, 韓寶瑜. 外源水楊酸甲酯(MeSA)處理茶樹對茶園主要害蟲及其天敵的影響[J]. 生態學雜志, 2011, 30(3): 564 -568.
[11] 楊恕玲, 單守明, 鞏傳銀, 等. 水楊酸對休眠期茶樹光合作用和抗凍性的影響[J]. 中國農學通報, 2009, 25(15): 121-124.
[12] 周旋, 申璐, 肖霄, 等. 外源水楊酸對鹽脅迫下茶樹生長及抗氧化酶活性的影響[J]. 西北農業學報, 2014, 23(6): 127-133.
[13] Caemmerer S V, Farquhar G D. Some relationships between the biochemistry of photosynthesis and the gas-exchange of leaves [J]. Planta, 1981, 153(2): 376-387.
[14] Ethier G J, Livingston N J. On the need to incorporate sensitivity to CO2transfer conductance into the Farquhar-von Caemmerer-Berry leaf photosynthesis model [J]. Plant Cell and Environment, 2004, 27(2): 137-153.
[15] Okuda T, Matsuda Y, Yamanaka A, et al. Abrupt increase in the level of hydrogen peroxide in leaves of winter wheat is caused by cold treatment [J]. Plant Physiology, 1991, 97(3): 1265-1267.
[16] Willekens H, Chamnongpol S, Davey M, et al. Catalase is a sink for H2O2and is indispensable for stress defence in C3 plants [J]. The EMBO Journal, 1997, 24(16): 4806-4816.
[17] Wang S D, Zhu F, Yuan S, et al. The roles of ascorbic acid and glutathione in symptom alleviation to SA-deficient plants infected with RNA viruses [J]. Planta, 2011, 234(1): 171-181.
[18] Nakano Y, Asada K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts [J]. Plant and Cell Physiology, 1981, 22(1): 867-880.
[19] Patra H K, Kar M, Mishra D. Catalase activity in leaves and cotyledons during plant development and senescence [J]. Biochemie Und Physiologie Der Pflanzen, 1978, 172(1): 385-390.
[20] 李治鑫, 李鑫, 范利超, 等. 高溫脅迫對茶樹葉片光合系統的影響[J]. 茶葉科學, 2015, 35(5): 415-422.
[21] Ananiera E A, Alexieva V S, Popoval LP. Treatment with salicylic acid decreases the effects of paraquat on photosynthesis [J]. Journal of Plant Physiology, 2002, 159(7): 685-693.
[22] 王利軍, 劉允芬, 劉琪璟, 等. 高溫干旱脅迫下水楊酸和鉀對溫洲蜜柑光合作用和葉綠素熒光的影響[J]. 江西科學, 2003, 21(1): 202-205.
[23] 王利軍, 黃衛東. 高溫脅迫及其信號轉導[J]. 植物學通報, 2000, 17(2): 114-120.
[24] 云建英, 楊甲定, 趙哈林. 干旱和高溫對植物光合作用的影響機制研究進展[J]. 西北植物學報, 2006, 26(3): 641-648.
[25] Andersson I, Backlund A. Structure and function of Rubisco [J]. Plant Physiology and Biochemistry, 2008, 46(3): 275-291.
[26] Stidham M A, Uribe E G, Williams G J. Temperature dependence of photosynthesis in. II. Contribution from electron transport and photophosphorylation [J]. Plant Physiology, 1982, 69(4): 929-934.
[27] 陳培琴, 郁松林, 詹妍妮, 等. 植物在高溫脅迫下的生理研究進展[J]. 中國農學通報, 2006, 22(5): 223-226.
[28] 高京草, 王慧霞, 李西選. 可溶性蛋白、丙二醛含量與棗樹枝條抗寒性的關系研究[J]. 北方園藝, 2010(23): 18-20.
[29] 王開凍, 顏志明, 馬衛軍, 等. 水楊酸對高溫脅迫下南瓜幼苗生理生化的影響[J]. 浙江農業科學, 2009(1): 42-45.
[30] 呂俊, 張蕊, 宗學鳳, 等. 水楊酸對高溫脅迫下水稻幼苗抗熱性的影響[J]. 中國生態農業學報, 2009, 17(6): 1168-1171.
[31] Salin M L. Toxic oxygen species and protective system of the chloroplast [J]. Physiology Plant, 1987, 72(1): 681-689.
[32] Dat J F, Lopez-Delgado H, Foyer C H, et al. Parallel changes in H2O2and catalase during thermotolerance induced by salicylic acid or heat acclimation in mustard seedlings [J]. Plant Physiology, 1998, 116(4): 1351-1357.
[33] 范雙喜, 谷建田, 韓瑩琰. 園藝植物高溫逆境生理研究進展[J]. 北京農學院學報, 2003, 18(2): 147-150.
[34] Mittler R. Oxidative stress, antioxidants and stress tolerance[J]. Trends in Plant Science, 2002, 9(7): 405-410.
[35]Willekens H, Langebartels C, Tiré C, et al. Differential expression of catalase genes in(L.) [J]. Proceeding of the National Academy of Sicence of the Unitied States of America, 1994, 91(22): 10450-10454.
[36] Fan M H, Sun X, Xu N J, et al. Integration of deep transcriptome and proteome analyses of salicylic acid regulation high temperature stress in[J]. Scientific Reports, 2017, 7(1): 11052. doi:10.1038/s41598- 017-11449-w.
[37] 杜朝昆, 李忠光, 龔明. 水楊酸誘導的玉米幼苗適應高溫和低溫脅迫的能力與抗氧化酶系統的關系[J]. 植物生理學通訊, 2005, 41(1): 19-22.
[38] Slaymaker D H, Navarre D A, Clark D, et al. The tobacco salicylic acid-binding protein 3 (SABP3) is the chloroplast carbonic anhydrase, Which exhibits antioxidant activity and plays a role in the hypersensitive defense response [J]. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(18): 11640-11645.
[39]Polidoros A N, Scalios J G. Role of hydrogen peroxide and different classes of antioxidants in the regulation of catalase and glutathione S-transferase gene expression in maize (L.) [J]. Physiologia Plantarum, 1999, 106(1): 112-120.
[40] Horváth E, Szalai G, Janda T. Induction of abiotic stress tolerance by salicylic acid signaling [J]. Journal of Plant Growth Regulation, 2007, 26(3): 290-300.
Effects of Exogenous Salicylic Acid on Photosynthesis and Antioxidant Enzymes of Tea Plants under HighTemperature
WEI Jipeng1, LI Xin1*, WANG Zhaoyang2, LI Yang1,3, ZHANG Lan1, SHEN Chen1, YAN Peng1, ZHANG Liping1, HAN Wenyan1*
1. Tea Research Institute, Chinese Academy of Agricultural Sciences, Hangzhou 310008, China; 2. Ankang Academy of Agricultural Sciences, Ankang 725000, China; 3. The Horticulture College of Hebei Agricultural University, Baoding 071000, China
In recent years, heat stress was more frequently occurred in tea gardens.However, few studies were focused on the approaches towards the improvement of heat tolerance in tea plants. In this study, Longjing 43 was used as experimental material to investigate the effects of various concentrations of MeSA on the net photosynthetic rate (Pn), the maximum carboxylation rate of Rubisco (V,max), the maximum RuBP regeneration rate (max), electrolyte leakage (EL), MDA content and antioxidant enzyme activities in tea leaves under heat stress. Results showed that 1?mmol·L-1MeSA could significantly increase Pn,Vmaxandmaxin tea leaves under heat stress. EL and MDA content in tea leaves increased significantly after heat stress, while the application of MeSA attenuated heat-induced increases in EL and MDA. In addition, the application of 1?mmol·L-1MeSA stimulated the activities ofAPX and CAT, leading to an efficient scavenging of hydrogen peroxide (H2O2) in tea leaves. In summary, we revealed that the applications of MeSA could improve photosynthetic capacity, strengthen the antioxidant system, reduce reactive oxygen species accumulation and lipid peroxidation in tea leaves under heat stress, and thus improve the tolerance of tea plants under heat stress.
salicylic acid, tea plant, heat stress, photosynthesis, antioxidant enzyme
S571.1;Q945.11
A
1000-369X(2018)04-353-10
2017-07-31
2017-10-09
國家重點研發計劃政府間國際科技創新合作重點專項(2017YFE0107500)、中國農業科學院科技創新工程(CAAS-ASTIP-2014-TRICAAS)、中國農業科學院茶葉研究所基本科研業務費專項項目(1610212016025)
魏吉鵬,男,碩士研究生,主要從事茶樹栽培與生理生化研究。*通訊作者:hanwy@tricaas.com
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
