甜瓜細菌性果斑病抗性基因連鎖的SSR分子標記
2018-09-10 07:11:20俞志杰寧雪飛王賢磊
江蘇農業科學 2018年15期
俞志杰, 寧雪飛, 王賢磊, 李 冠
(新疆大學生命科學與技術學院,新疆烏魯木齊 830046)
甜瓜(CucumismeloL.)是葫蘆科(Cucurbiaceae)甜瓜屬(CucumisL.)1年生蔓性草本植物,基因組大小為 450 Mb[1]。甜瓜在國內外廣泛種植,尤其新疆甜瓜在國內外享有盛譽,遠銷全國各地并出口到東南亞許多國家[2]。但近年來,細菌性果斑病已成為危害甜瓜產業的主要病害之一。國外研究報道,該病最早于1969年在美國福羅里達州被發現,近年來已迅速蔓延至13個國家和地區[3-4]。甜瓜大部分的經濟損失是由細菌性果斑病引起的病害,病發嚴重時細菌性果斑病引起的損失超過90%,甚至絕收[5-6]。我國新疆、內蒙古、海南等9個省份的甜瓜作物曾大規模暴發細菌性果斑病[7]。
1978年Schaad等將甜瓜細菌性果斑病病原菌鑒定為類產堿假單胞菌西瓜亞種(Pseudomonaspseudoalcaligenessubsp.citrulli)[8]。1992年Willems等根據rRNA-DNA和DNA-DNA分子雜交的結果,將該病原菌更名為燕麥嗜酸菌西瓜亞種(Acidovoraxavenaesubsp.citrulli,簡稱Aac)[9]。在國內,趙延昌等最早對新疆甜瓜細菌性果斑病病原菌進行了鑒定,認為引起甜瓜細菌性果斑病發生的病原菌是燕麥嗜酸菌西瓜亞種(Acidovoraxavenaesubsp.citrulli),屬革蘭氏陰性菌[10]。田間發生瓜類細菌性果斑病主要是由于病原菌通過傷口和氣孔侵染寄主,病害遠距離傳播是通過種子帶菌進行傳播[11-12]。目前,由于細菌性果斑病導致瓜類商品的經濟效益嚴重下降,對瓜類產業造成威脅,因此,必須采取有效的措施防治該病害的發生。
目前,田間防治果斑病主要依靠含銅的化學藥劑[13],長期使用化學試劑,不僅導致病原菌對化學試劑產生抗藥性,而且污染環境、威脅人類的健康。而在選育抗果斑病品種方面,目前主要集中在傳統雜交育種上。傳統的雜交育種不僅耗時耗力還在表型選擇上存在局限性,致使育種工作進展緩慢[14]。隨著分子生物學研究的不斷深入,利用分子標記輔助育種具有巨大的潛力,也為抗病育種提供了一項方便、快捷、高效的方法[15]。尤其是甜瓜基因組序列已于2012年6月公布[16],科研工作者利用基因組序列開展甜瓜相關的分子育種工作變得非常方便。因此,發掘抗細菌性果斑病資源,利用分子標記手段輔助培育高抗果斑病的新品種是解決問題的有效途徑。雖然國外和國內的科學家在抗果斑病種質鑒定方面做出了很多努力,但關于抗果斑病基因的遺傳機制研究較少,對遺傳規律的發掘相對滯后[17],因此,研究甜瓜果斑病抗性基因的遺傳分析及定位已迫在眉睫。本試驗以遼寧神帥抗病品系和皮山奎瑞克感病品系的F2分離群體作為試驗材料,采用集群分離分析法(bulked segregation analysis,簡稱BSA)和簡單重復序列(simple sequence repeat,簡稱SSR)分子標記的方法進行果斑病抗性基因的遺傳規律和定位研究,以期了解抗病品種遼寧神帥所含的抗性基因,為今后的抗果斑病基因精細定位和抗性種質資源應用于甜瓜品種的改良奠定基礎。
1 材料與方法
1.1 試驗材料
用于構建分離群體的抗病親本材料遼寧神帥和感病親本材料皮山奎瑞克,均由國家瓜類工程研究技術中心提供。果斑病病原菌采自國家瓜類工程技術研究中心試驗田(新疆昌吉地區),在新疆大學生物工程研究中心凍存。
1.2 主要試驗試劑
溴化十六烷三甲基銨(cetyltrimethyl ammonium bromide,簡稱CTAB)、β-巰基乙醇、聚乙烯吡咯烷酮(PVP-40)、Tris-base、RNase A等,均購自生工生物工程(上海)股份有限公司;用于PCR相關反應的試劑及Marker,均購自貝思汀生物科技有限公司;SSR引物由華大基因合成。其他分子試劑均為國產或進口分析純。
1.3 試驗方法
1.3.1 病原菌菌懸液的制備 果斑病病原菌的擴繁活化采用LB固體培養基(5 g胰化蛋白胨、2.5 g酵母提取物、2.5 g NaCl、7.5 g瓊脂,用蒸餾水定容至500 mL,pH值為7.4),在28 ℃溫度下培養36 h,并在無菌條件下挑取單菌落于LB液體培養基中,在28 ℃、220 r/min條件下培養24 h,用無菌水將菌懸浮液濃度調至5×108CFU/mL用于接種[18]。
1.3.2 接種及病情鑒定 抗病鑒定試驗于2016年6—7月在新疆大學生物研究中心進行,將抗病親本、感病親本各10粒、F2分離群體120粒種子種植于營養缽中。當幼苗生長至2~4張真葉完全展開時(約生長25 d)[17],將待用的菌懸液采用噴霧接種的方法接種,并及時用塑料膜保溫保濕。10~15 d充分發病后調查病情等級,本研究參照Hopkins等的分級標準[12],并適當修改(表1)。
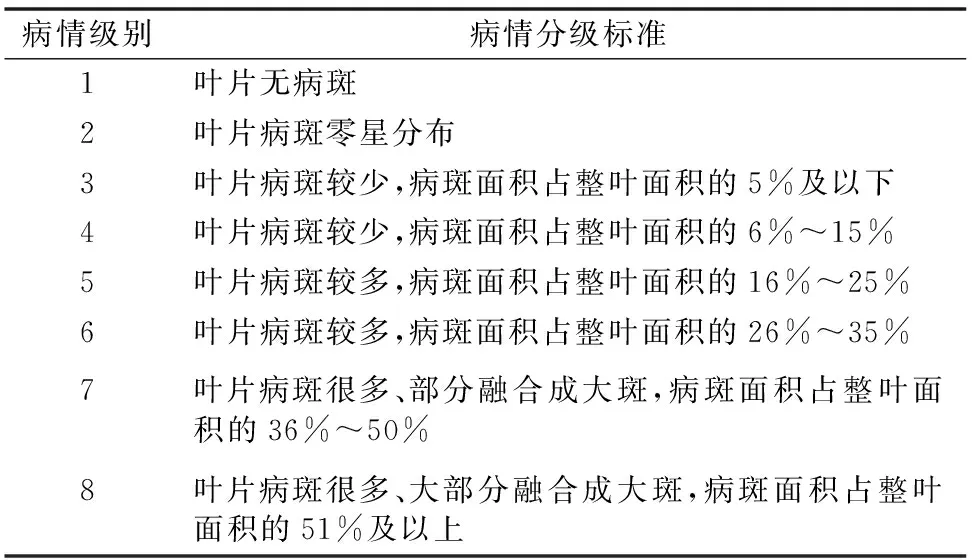
表1 單株幼苗感果斑病分級標準
1.3.3 抗感池的構建和基因組DNA的提取 用病情級別為 0~1級的21個極抗植株構建抗病池,用病情級別為7~8級的11個感病植株構建感病池。采用Porebski等的CTAB法[19]提取各單株的DNA,并將提取的DNA溶液稀釋至 100 ng/μL,-20 ℃保存備用。
1.3.4 抗果斑病基因的初步定位 參照文獻[20-21],均勻選取分布于甜瓜12條連鎖群上的288個SSR標記,對2個基因池進行檢測,篩選在抗病池、感病池之間表現多態性的SSR分子標記。SSR擴增體系為20.00 μL,其中DNA模板0.40 μL,10×PCR buffer(含15 mmol/L Mg2+)2.00 μL,上、下游引物(50 μmol/L)各0.20 μL,10 mmol/L dNTPs 0.40 μL,TaqDNA酶(Biomed,China,5 U/μL)0.24 μL,用ddH2O補足20.00 μL。SSR擴增程序:94 ℃預變性3 min;94 ℃變性 30 s,55 ℃退火30 s,72 ℃延伸30 s,35個循環;72 ℃延伸 5 min;4 ℃保存。SSR擴增產物用6%變性聚丙烯酰胺凝膠電泳檢測,銀染顯色。
1.3.5 數據處理與初步定位的結果驗證 用Excel 97-2003和JoinMap 4.0處理、分析試驗數據。在抗病池、感病池之間有多態性的標記用F2個體驗證,從而確定初步定位的結果。與感病親本的帶型相同記為“A”,與抗病親本的帶型相同記為“B”,F1帶型記為“H”,缺失或模糊不清記為“-”。
2 結果與分析
2.1 F2群體抗性分析
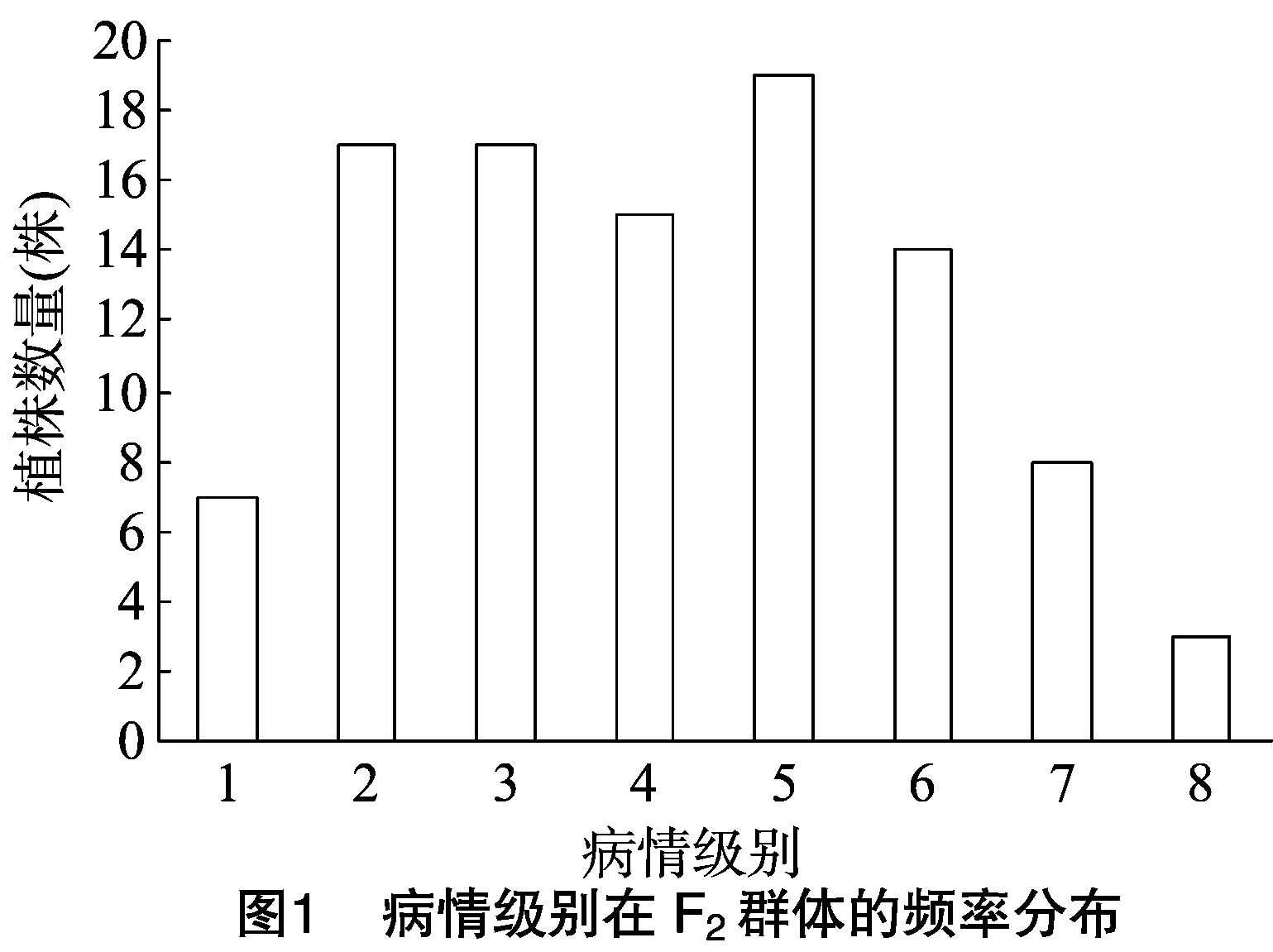
種植的120粒F2分離群體種子,除去未發芽和弱小植株外,對剩余的100株幼苗進行病情調查。由圖1可知,將1~5級記為抗病,6~8級記為感病,F2世代的病情分級呈連續性分布,說明甜瓜抗果斑病基因屬于數量性狀位點(quantitative trait locus,簡稱QTL)。
2.2 抗病池、感病池引物的篩選
用288對SSR引物分別對抗病池、感病池進行PCR,經過多次重復后,由圖2可知,有7對引物在抗病池、感病池之間能夠穩定地擴增出多態條帶,多態性比率為2.43%,依次為EMAGN73(95)、ECM122(96)、GCM336(101)、ECM53(102)、ECM231(106)、GCM246(111)、CMAGN61(114),其多態性引物序列如表2所示。7對標記都分布于連鎖群(linkage group,簡稱LG)Ⅳ上,因此將抗果斑病基因初步定位到LGⅣ上。
2.3 初步定位結果驗證
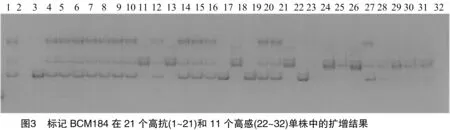
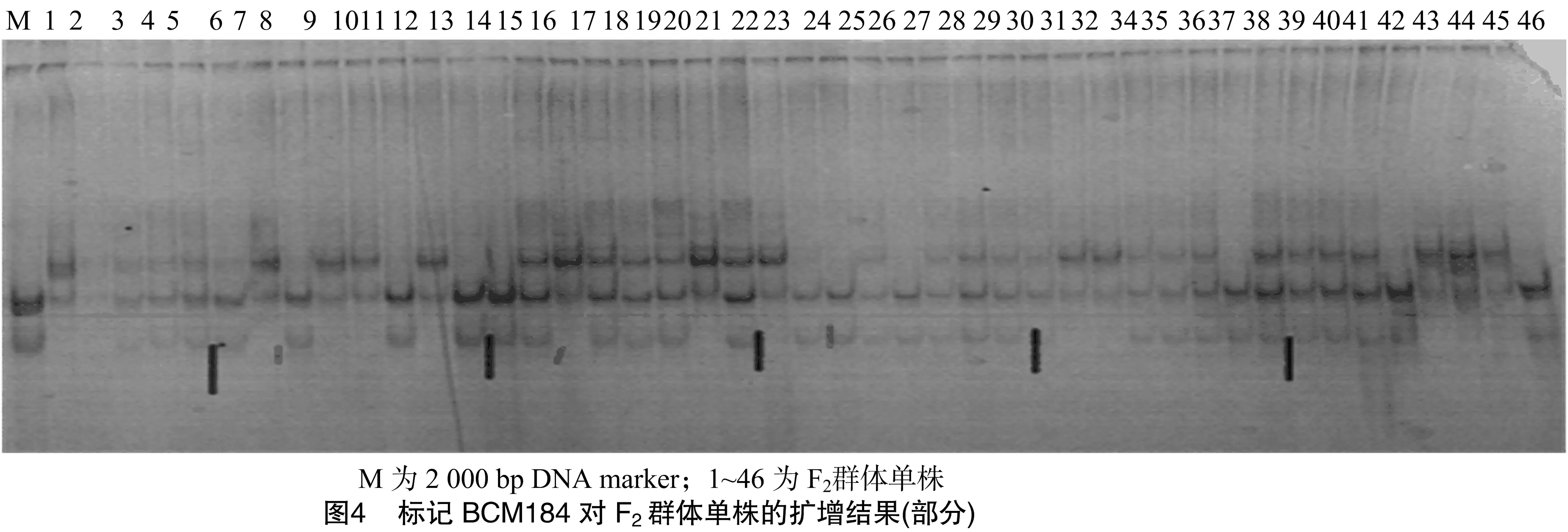
篩選出的7對引物分別在100個F2群體的單株中進行擴增、檢測,其中BCM184標記與抗性基因連鎖最為緊密。由圖3可知,BCM184標記在21個高感單株中擴增,除個體2無擴增條帶外,個體11、13、18均擴增出感親多態性條帶。在11高感單株中,個體24沒有擴增出條帶,個體23擴增出抗親多態性條帶。由此說明,BCM184標記與甜瓜果斑病抗性基因具有連鎖關系。用100個F2分離群體進行個體驗證(結果如圖4),BCM184標記與抗果斑病基因的連鎖距離為12.4 cM。

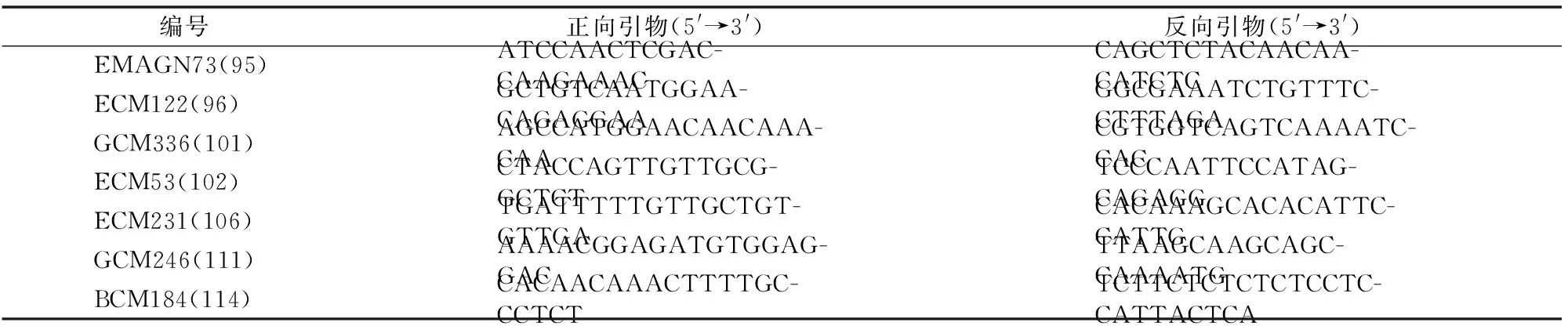
表2 抗感基因池間表現多態性的SSR引物
3 討論與結論
關于果斑病的報道,國內外主要圍繞瓜類細菌性果斑病病菌的分離檢測、致病機制、遺傳多樣性及防治等方面的研究[13]。筆者所在實驗室多年來一直從事甜瓜抗病研究,積累了較為豐富的經驗,筆者認為果斑病已成為威脅甜瓜產業的主要病害之一,但其抗性基因的定位之所以鮮有報道,原因是抗病種質資源有待發掘、抗性基因的遺傳機制符合QTL遺傳等[22-23],導致后續定位難度增加。
國內外果斑病接種方法包括浸種法、噴霧法、瓜期接種法、離體法等。浸種法是將種子在菌液中浸泡10~12 h,雖然該方法能夠快速接種,但會造成幼苗迅速死亡,損失率高達80%[24],因此不適合本試驗接種。瓜期接種法一般選擇開花授粉15 d后大小一致、健康、坐果節相同的甜瓜果實[7],此方法的缺點是不適合實驗室等小規模試驗,且周期較長。離體法具有簡便、準確可靠、易于人工控制溫度、濕度等外界環境因素,但此法與瓜期接種法的缺點相同。本試驗采用的接種方法是噴霧法,李俊閣用浸種、噴霧、針刺、離體等4種方法對種質資源進行抗性種質的篩選,其中用噴霧法接種病原菌后對甜瓜葉片可溶性蛋白、過氧化物酶、多酚氧化酶、苯丙氨酸解氨酶等生理指標進行測定,結果發現抗病材料和感病材料有明顯的差異[7]。通過苗期噴霧接種鑒定,調查植株所有葉片病斑面積大小統計病情指數,以此確定不同甜瓜材料之間的抗病指數,可以較準確地反映不同材料之間存在的差異[17]。
SSR標記相比于其他的分子標記如擴增的限制性內切酶片段長度多態性(amplified fragment length polymorphism,簡稱ARLP)、隨機擴增多態性(randomly amplified polymorphism,簡稱RAPD)、相關序列擴增多態性(sequence-related amplified polymorphism,簡稱SRAP)等,明顯的優勢在于分布廣、共顯性、操作簡單、高度可重復等[25-26]。因此,SSR標記被廣泛應用于遺傳圖譜的構建、基因定位、分子輔助育種、種子純度鑒定、親子鑒定、遺產多樣性等研究中[27-30]。近年來,利用SSR分子標記定位的甜瓜基因包括裂葉基因pll、矮化基因mdw1、短側枝基因slb、抗白粉病基因Pm-2F、Pm-Edisto47-1、Pm-AN等[31]。鑒于SSR分子標記的諸多優點以及前人采用SSR分子標記技術在甜瓜多項研究中取得的良好試驗結果,本試驗采用SSR分子標記技術對甜瓜抗細菌性果斑病進行定位研究。
本試驗中病情調查結果顯示,F2抗病植株和感病植株分別有75、25株。因此,將抗病植株細化分為5個等級,而感病品種分為3個等級。BCM184標記在高感21個單株中擴增,擴增出17個抗親多態性條帶和F1多態性條帶,3個感親多態性條帶。在高感單株中,擴增出2個抗親多態性條帶和F1多態性條帶,8個感親多態性條帶。這一結論與表型性狀相吻合,說明抗果斑病基因受部分顯性基因控制。本試驗對抗性品種遼寧神帥所含的抗性基因的遺傳規律及所屬連鎖群的研究,為進一步開展甜瓜抗細抗菌性果斑病基因定位克隆和抗性甜瓜的選育奠定了基礎。
