荒漠草原中間錦雞兒灌叢的土壤微生物多樣性
2018-09-11 08:07:08王德凱王躍棠梅續芳解李娜李清芳馬成倉
天津師范大學學報(自然科學版) 2018年4期
王德凱 ,李 穎 ,王躍棠 ,梅續芳 ,解李娜 ,李清芳 ,馬成倉
(1.天津師范大學生命科學學院,天津300387;2.天津師范大學天津市動植物抗性重點實驗室,天津300387)
土壤微生物是生活在土壤中的細菌、真菌、放線菌、藻類的總稱.其組成和結構一方面受土壤性質的影響,如土壤水分、土壤溫度、pH、土壤養分、土壤質地等[1-2];另一方面,土壤微生物群落與植物生長過程緊密相關,植物通過凋落物、根系分泌物、植物生產力等影響土壤微生物群落[3-4].灌木已成為全球草地生態系統的重要組成部分,其密度和蓋度較大且呈逐年增加趨勢.灌叢的形成改變了地表土壤資源的分配,使水分和養分較多地聚集在灌叢內,導致土壤資源具有空間異質性,形成“沃島”[5].“沃島”的形成又會引發土壤有機碳和其他成分的空間異質性,加劇了資源的異質化和斑塊化[6],從而影響土壤微生物群落.灌叢土壤資源異質化又會引起灌叢內外植被的異質化,進一步影響土壤微生物群落[7-8].
錦雞兒屬(Caragana Fabr.)植物是內蒙古草原最常見的灌木,具有很強的適應干旱、寒冷環境和耐貧瘠能力,不僅是牲畜的優質牧草,還能夠防風固沙、保持水土[9].本研究以內蒙古荒漠草原的中間錦雞兒灌叢的土壤微生物為研究對象,通過傳統培養法和分子生物學技術,比較灌叢內外土壤微生物群落的差異,探討中間錦雞兒灌叢形成導致的土壤資源異質化對土壤微生物的影響,進一步理解中間錦雞兒灌叢的沃島效應.
1 材料與方法
1.1 研究區概況
樣地位于內蒙古鄂托克旗的烏蘭鎮(北緯39°1′,東經107°98′),該地區為典型的溫帶大陸性季風氣候,日照充足,四季分明,年平均降水量約為250 mm,降水主要集中在7—9月份.年平均蒸發量為2 400~2 800 mm,是降水量的8到12倍.年平均氣溫為6.5℃,年日照時數約3 000 h.土壤質地以疏松的砂質壤土或沙土為主.
1.2 研究方法
1.2.1 土壤樣品采集
2016年7月在鄂托克旗烏蘭鎮選取3個優勢種為中間錦雞兒的地區作為樣地,每個樣地隨機選取3株中間錦雞兒灌叢,在灌叢內和灌叢外(距離灌叢邊緣 2 m)分別采集 0~10 cm(A 層)、10~20 cm(B 層)、30~40 cm(C層)的土壤樣品約300 g,用自封袋密封.樣品于4℃保存,一部分用于土壤理化性質的測定,一部分用于土壤微生物多樣性的測定.
1.2.2 土壤理化性質的測定
采用水浸提電位法測定土壤pH值;采用烘干法測量含水量;采用重鉻酸鉀容量法測定有機質(Organic matter)含量;采用碳酸氫鈉浸提-鉬銻抗比色法測定速效磷(Available phosphorus)含量;采用醋酸銨浸提-火焰光度法測定速效鉀(Available potassium)含量;使用流動分析儀測定硝態氮(Nitrate nitrogen)和銨態氮(Ammonium nitrogen)含量.將土樣浸提,然后用DDS~LLA型電導儀測定電導率(Electricconductivity)[10].中間錦雞兒灌叢內外的土壤理化性質如表1所示.
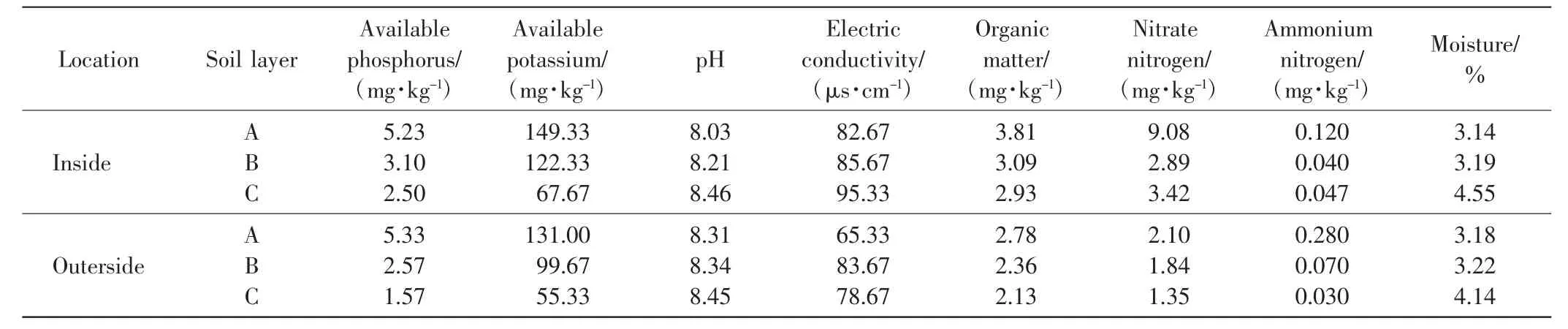
表1 中間錦雞兒灌叢內外土壤理化性質Tab.1 Physical and chemical characteristics of soil inside and outside Caragana intermedia shrubs
1.2.3 土壤微生物群落的多樣性測定
(1)土壤微生物培養:分別采用牛肉膏蛋白胨培養基、馬丁氏培養基和高氏一號培養基法培養土壤中的細菌、真菌和放線菌,采用稀釋平板計數法計數,結果以每克干土所含的微生物菌落數表示.用平板劃線分離法分離純化微生物菌落.
1g干土中的菌落數(cfu/g)=M×鮮土質量/干土質量式中:M=a×u/V,a為培養皿中的平均菌落數,u為稀釋倍數,V為每個培養皿中加入的土壤懸液的體積(本實驗中為0.2 mL).
(2)微生物的分子生物學鑒定:細菌和放線菌DNA采用生工Ezup柱式細菌基因組DNA抽提試劑盒進行提取,真菌DNA采用生工Ezup柱式真菌基因組DNA抽提試劑盒提取,按照試劑盒說明書進行操作.采用通用引物 1492R(5′-GGT TAC CTT GTT ACGACT T-3′)和 27F(5′-AGA GTT TGA TCC TGG CTCAG-3′)擴增細菌和放線菌16S rDNA片段,GeoA2(5′-CCA GTA GTC ATA TGC TTG TCT C-3′)和 Geo11(5′-ACC TTG TTA CTT TTA CTT CC-3′)擴增真菌18S rDNA片段.PCR擴增產物由北京華大基因公司測序,所得堿基序列提交至GenBank數據庫,使用BLAST程序進行比對分析,將菌株鑒定到屬.
采用Shannon-Wiener多樣性指數(H′)分析評價土壤微生物群落的多樣性[11].
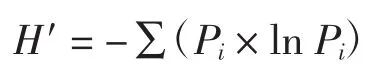
式中:Pi表示群落中第i個物種的個體數占群落中個體總數的比例.
1.3 數據分析
利用SPSS17.0軟件進行單因素方差分析(One-way ANOVAs)(P<0.05),分析灌叢內外不同土層間微生物菌落數量和多樣性的差異.用Tukey HSD做多重比較.采用獨立樣本t檢驗分析灌叢內外的差異.采用CANOCO 4.5軟件對土壤微生物和環境因子之間的關系進行冗余分析.用Monte Carlo置換檢驗分析環境因子對土壤微生物群落影響的統計學意義(P<0.05).
2 結果與分析
2.1 中間錦雞兒灌叢土壤微生物的數量
中間錦雞兒灌叢內外不同土層的土壤微生物數量如圖1所示.

圖1 灌叢內外土壤細菌、放線菌和真菌菌落總數Fig.1 Quantity of bacteria,actinomycetes and fungi inside and outside the shrubs
由圖1可以看出,土壤微生物總數表現為:細菌>放線菌>真菌.灌叢內土壤細菌數隨著土層深度的增加顯著下降(P<0.05),而灌叢外則呈先緩慢增加后減少的趨勢;同一土層灌叢內土壤細菌總數均顯著高于灌叢外的數量(P<0.05).灌叢內外放線菌總數均表現為A層>B層>C層,且不同土層間的差異具有統計學意義(P<0.05);同一土層灌叢內的土壤放線菌總數均大于灌叢外的數量,A層和B層灌叢內外的差異具有統計學意義(P<0.05),C層灌叢內外數量相近(P>0.05).灌叢內外的土壤真菌數隨著土層深度的增加均呈先增加后減少的趨勢,不同土層之間的差異具有統計學意義(P<0.05);同一土層灌叢內土壤真菌總數均顯著高于灌叢外的數量(P<0.05).
2.2 中間錦雞兒灌叢土壤微生物的群落組成
2.2.1 細菌
本研究從中間錦雞兒灌叢內外土壤中共分離出細菌13屬,如表2所示.無論是灌叢內還是灌叢外,A層的優勢屬均為芽孢桿菌屬(Bacillus)和短小桿菌屬(Curtobacterium Yamada),B層和C層的優勢屬為芽孢桿菌屬、短小桿菌屬和類芽孢桿菌屬(Paenibacillus Ash),優勢屬的相對豐度在不同土層間的差異具有統計學意義(P<0.05).在灌叢內,芽孢桿菌屬的相對豐度隨土層深度增加顯著下降(P<0.05),灌叢外則無顯著差異(P>0.05).無論是灌叢內還是灌叢外,類芽孢桿菌屬的相對豐度均隨土層深度的增加而顯著升高(P<0.05).在灌叢內,短小桿菌屬在B層的數量最多,在灌叢外則是A層數量最多.壤霉菌屬(A-gromyces)、溶桿菌屬(Lysobacter)、鞘氨醇桿菌屬(Sphingobacterium sp)、嗜麥芽窄食單胞菌屬(Stenotrophomonas maltophilia))、金黃桿菌屬(Chryseobacterium))、假單胞菌屬(Pseudoxanthomonas)為灌叢內特有屬,且壤霉菌屬、溶桿菌屬、鞘氨醇桿菌屬、金黃桿菌屬均集中在灌叢內的A層.節桿菌屬(Arthrobacter)僅在A層出現.葡萄球菌屬(Staphylococcus)在各土層均能檢測到,灌叢內各土層間的差異較小(P>0.05),灌叢外則隨著土層深度的增加呈下降趨勢(P<0.05).中華根瘤菌屬(Sinorhizobium Chen)在灌叢內僅在A層檢測到;灌叢外則分布于A層和B層.微桿菌屬(Microbacterium)在灌叢內的各個土層均可檢測到,且A層和C層數量相近,顯著高于B層;灌叢外則出現于B層和C層,且后者數量顯著高于前者(P<0.05).
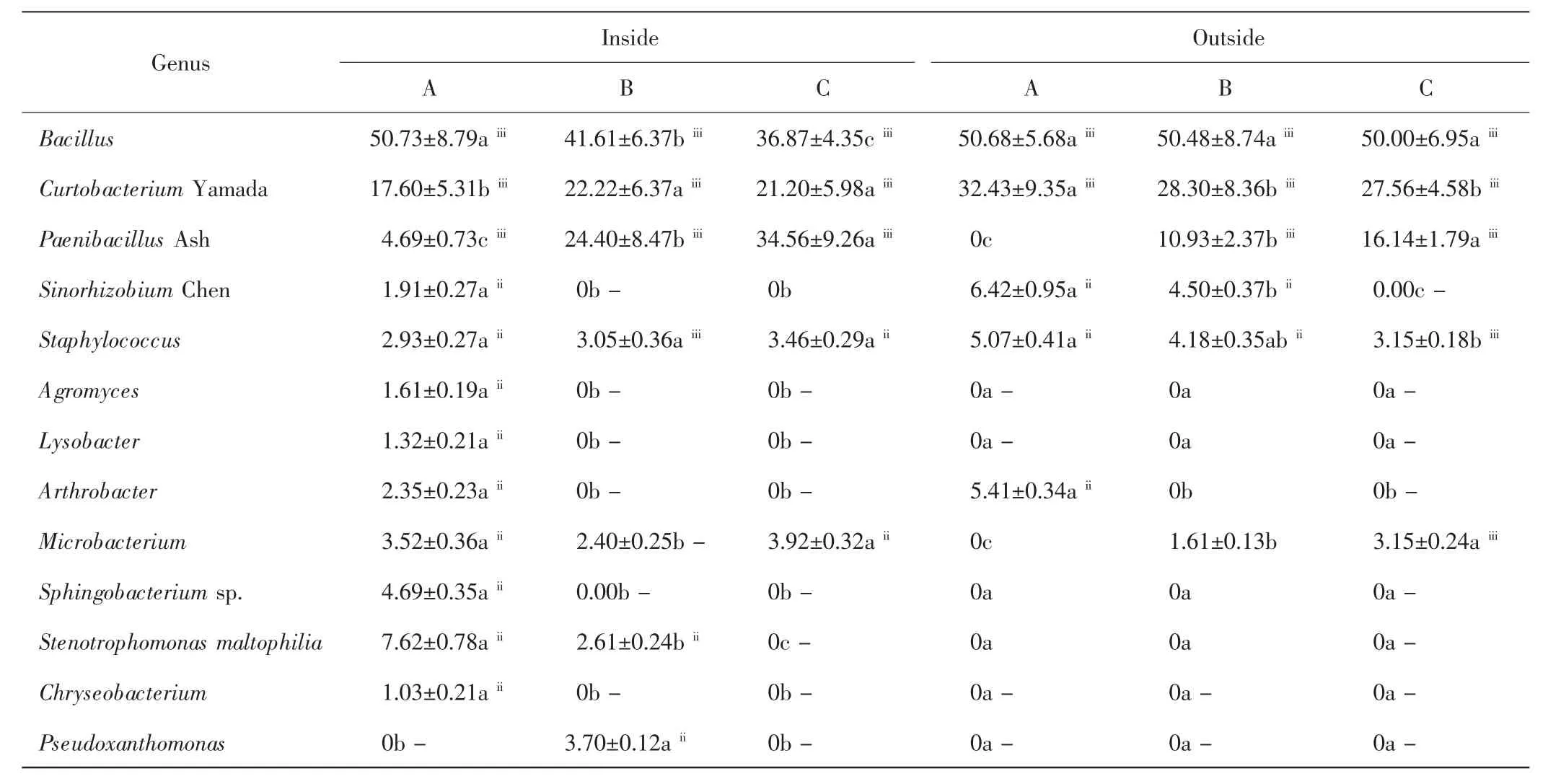
表2 灌叢內外土壤細菌群落組成及相對豐度Tab.2 Community composition and relative abundance of bacteria inside and outside shrubs %
2.2.2 真菌
中間錦雞兒灌叢內外土壤中共分離出真菌12屬,如表3所示.由表3可以看出,曲霉屬(Aspergillus)為灌叢內外各土層的優勢屬,且相對豐度隨著土層深度的增加顯著增加(P<0.05).小囊菌(Microascaceae)為灌叢內B、C層和灌叢外C層的優勢屬.青霉屬(Penicillium)主要集中在灌叢內的A層和灌叢外的B、C層,灌叢內的相對豐度隨著土層深度的增加顯著降低(P<0.05),灌叢外則是B層數量最多.莖點霉屬(Phorna)和隱球菌屬(Cryptococcus)均僅出現在灌叢內外的A層,而鏈格孢霉屬(Alternaria)出現在A層和B層.木霉屬(Trichoderma)、露漆濕斑菌屬(Myrothecium roridum)和小麥葉枯病菌(Septoriatritici)均為灌叢內特有屬,疣孢漆斑菌屬(Myrothecium verrucaria)為灌叢外特有屬.鐮刀菌屬(FusariumLink)、球毛殼菌屬(Chaetomium)和小囊菌(Microascaceae)在灌叢內外的各個土層均能檢測到,通常富集在B層和C層.

表3 灌叢內外土壤中真菌群落組成及相對豐度Tab.3 Community composition and relative abundance of fungi inside and outside shrubs %
2.2.3 放線菌
中間錦雞兒灌叢內外土壤中共分離出了放線菌5屬,如表4所示.灌叢內和灌叢外各土層的優勢屬均為鏈霉菌屬(Streptomyces),相對豐度占各土層放線菌總數的83%~95%,B層的相對豐度顯著高于A層和 C 層(P<0.05).小單孢菌屬(Micromonospora)僅在灌叢內出現,為灌叢內A層的優勢屬.諾卡氏菌屬(Nocardia)同樣僅出現在灌叢內,為灌叢內C層的優勢屬,且相對豐度隨著土壤深度增加顯著升高(P<0.05).Dyadobacter屬僅在灌叢內的A層檢測到.

表4 灌叢內外土壤中放線菌群落的組成及相對豐度Tab.4 Community composition and relative abundance of actinomycetes inside and outside shrub canopies %
2.3 中間錦雞兒灌叢土壤微生物多樣性
中間錦雞兒灌叢不同土層的土壤微生物多樣性指數計算結果如表5所示.
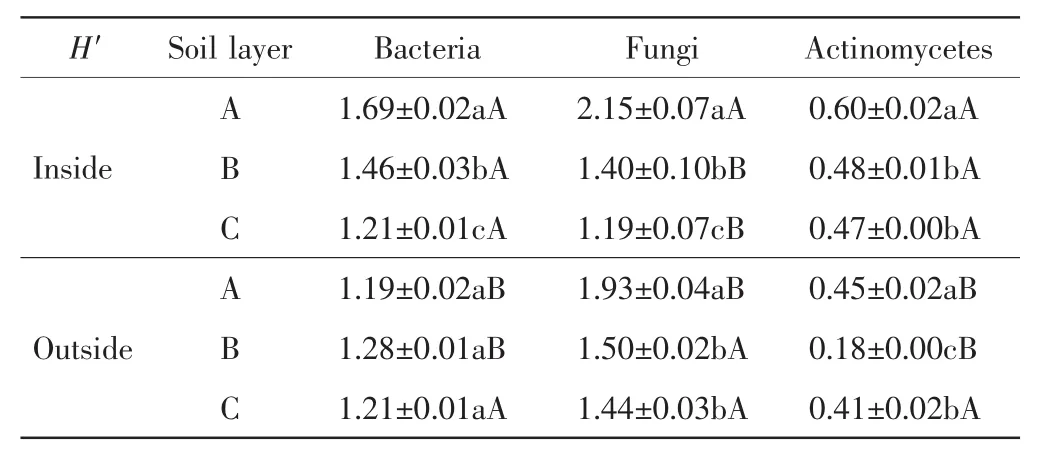
表5 灌叢內外土壤微生物的Shannon-Wiener指數Tab.5 Shannon-Wiener index(H′)of soil microorganisms inside and outside the shrubs
由表5可以看出,中間錦雞兒灌叢內土壤細菌的H′指數隨著土層深度的增加顯著下降(P<0.05),灌叢外則隨著土層深度的增加先增加后下降,但不同土層之間的差異不具有統計學意義(P>0.05);A層和B層灌叢內的細菌H′指數顯著高于灌叢外的數值,C層灌叢內外的數值相同.土壤真菌的H′指數在灌叢內外均隨著土層深度的增加呈下降趨勢;A層灌叢內的數值顯著高于灌叢外的數值,B層和C層則是顯著低于灌叢外的數值(P<0.05).灌叢內土壤放線菌的H′隨著土層深度的增加呈下降趨勢(P<0.05),而灌叢外H′隨著土層深度的增加先降低后增加(P<0.05);A層和B層灌叢內的放線菌H′指數顯著高于灌叢外的數值(P<0.05),C層數值相近(P>0.05).
2.4 土壤微生物群落與環境之間的關系
2.4.1 土壤細菌群落與環境的關系
對中間錦雞兒灌叢土壤細菌群落與環境之間的關系進行冗余分析,結果如圖2所示.

圖2 土壤細菌群落和環境因子的冗余分析Fig.2 RDA diagram of the relationship between soil bacterial communities and environmental factors
RDA的直接排序圖顯示,第一軸、第二軸對響應變量的解釋比例分別為56.7%和25.6%,Monte Carlo置換檢驗排序軸均達到顯著水平(第一排序軸:F=9.156,P<0.05;所有排序軸:F=5.147,P=0.002),說明環境因子能夠顯著影響土壤細菌群落.與第一軸關系最密切的是速效鉀(r=0.706 5),與第二軸關系最緊密的是灌叢(r=0.8467).RDA排序圖結果表明,假單胞菌屬、溶桿菌屬、嗜麥芽窄食單胞菌屬、芽胞桿菌屬、壤霉菌屬、溶桿菌屬、金黃桿菌屬和鞘氨醇桿菌屬與土壤有機質、硝態氮含量呈顯著正相關,但與土壤pH呈顯著負相關.類芽孢桿菌屬與土壤含水量呈顯著正相關,與銨態氮含量呈顯著負相關.中華根瘤菌屬與銨態氮含量呈顯著正相關,但與土壤含水量呈顯著負相關.節桿菌屬與速效磷、速效鉀含量呈顯著正相關,與土壤土壤含水量也呈顯著負相關.
2.4.2 土壤真菌群落與環境的關系
對中間錦雞兒灌叢土壤真菌群落與環境之間的關系進行冗余分析,結果如圖3所示.
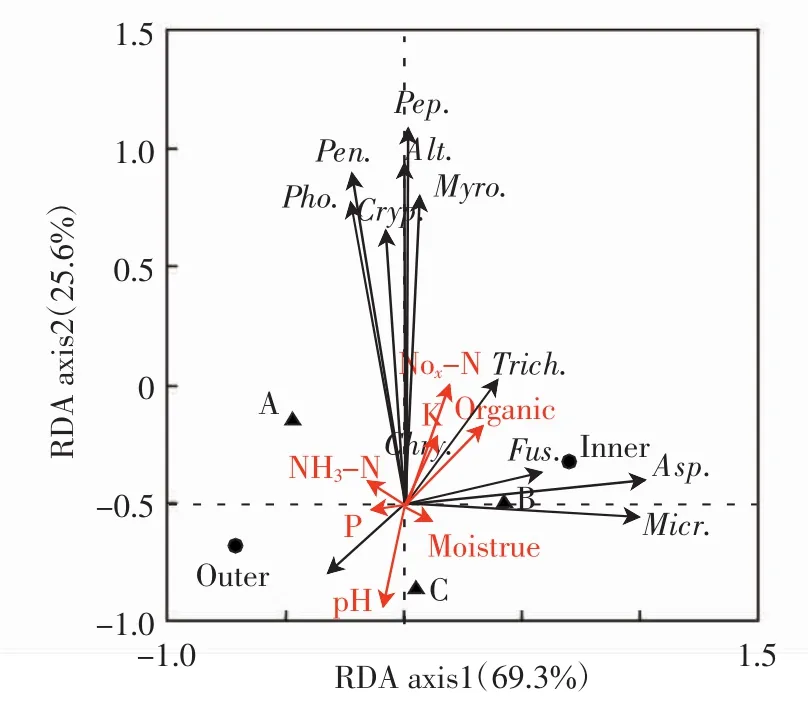
圖3 土壤真菌群落和環境因子的冗余分析Fig.3 RDA diagram of the relationship between soil fungal communities and environmental factors
RDA直接排序圖顯示,第一軸、第二軸對響應變量的解釋比例分別為69.3%和11.3%,Monte Carlo置換檢驗排序軸均達到顯著水平(第一排序軸:F=15.825,P<0.05;所有排序軸:F=8.020,P<0.05).與第一軸關系最密切的是灌叢(r=0.754 7),與第二軸關系最緊密的是硝態氮含量(r=0.863 1).球毛殼菌屬和木霉屬與硝態氮含量、速效鉀含量和有機質含量呈顯著正相關,與土壤pH呈顯著負相關(P<0.05).小囊菌屬與土壤含水量呈顯著正相關,與速效磷含量呈顯著負相關(P<0.05).疣孢漆斑菌屬與速效磷含量含量、土壤pH呈顯著正相關,與土壤有機質含量呈顯著負相關(P<0.05).
放線菌冗余分析Monte Carlo置換檢驗未達到顯著水平,說明環境因子對放線菌的影響不顯著.
3 討論與結論
本研究對內蒙古荒漠草原中間錦雞兒灌叢內外的微生物多樣性進行調查,結果發現,土壤可培養細菌的數量最多,其次是放線菌,真菌數量最少,這與多數研究[12-13]結果一致.土壤微生物的數量和群落多樣性存在明顯的垂直分布異質性.如苦豆子、芨芨草、油蒿等的土壤微生物數量隨土層加深呈下降趨勢[14-16];小興安嶺的原始紅松林、硬闊葉林和白樺林3種林地的土壤微生物多項多樣性指數隨著土層深度的增加顯著下降[17].細菌和真菌的豐富度隨土壤深度的增加而下降,而放線菌的豐富度隨土壤深度的增加而增加[18-19].本研究結果表明,中間錦雞兒灌叢的土壤可培養細菌、真菌和放線菌的數量及H′均表現為表層土或次表層土最大,這主要是因為土壤表層覆蓋有較多的凋落物,凋落物的分解提高了土壤表層養分含量,有利于表層土中土壤微生物的生長.另外,土壤性質隨著土層深度的變化而變化,從而影響土壤微生物群落.本研究通過RDA分析發現土壤速效鉀和硝態氮均與土壤可培養細菌和真菌的關系密切,說明速效鉀和硝態氮是影響中間錦雞兒土壤細菌和真菌的主要環境因素.RDA分析還表明土壤可培養放線菌數量與環境因子之間的相關性不顯著,說明環境變化對放線菌群落的影響不大.中間錦雞兒灌叢內的土壤細菌、真菌、放線菌數量和多樣性均大于灌叢外,這是由于中間錦雞兒灌叢的生長發育引起土壤資源空間異質性,灌叢內土壤養分聚集,形成比周圍環境更適宜的微生境[20-21],從而有利于土壤微生物生長.與灌叢外相比,灌叢內的植物多樣性更高,這也會增加土壤有機質含量并調節土壤含水量,即改善了土壤微生物的生存環境[22-23].
在土壤細菌中,Lysobacter含有較高的G+C含量和生物分解活性,能分泌各種抗生素、酶和生物活性物質,并抑制有害細菌的滋生,控制植物疾病.中間錦雞兒灌叢中,Lysobacter僅出現在灌叢內表層,這對中間錦雞兒灌叢具有保護作用.諾卡氏菌能夠在一定程度上指示土壤肥力的高低,諾卡氏菌越多,土壤肥力越高.本研究結果表明諾卡氏菌屬只出現在灌叢內,也說明了灌叢內的土壤肥力高于灌叢外的肥力.
