NaCl對(duì)德氏乳桿菌AGDI基因轉(zhuǎn)錄及產(chǎn)腐胺的影響
2018-09-13 06:27:38李蕊婷盧士玲
食品工業(yè)科技 2018年15期
關(guān)鍵詞:生長(zhǎng)
毛 順,李蕊婷,盧士玲
(石河子大學(xué)食品學(xué)院,新疆石河子 832003)
生物胺廣泛存在于發(fā)酵食品中,適量攝入對(duì)人體有益,但攝入過(guò)量則可能導(dǎo)致呼吸、消化、神經(jīng)和高血壓等疾病。生物胺主要由微生物所含有的氨基酸脫羧酶將游離氨基酸脫羧而成[1]。多種微生物如腸桿菌、微球菌、乳酸菌、芽孢桿菌、假單胞菌都能形成生物胺。腐胺作為生物胺的一種,不僅是精胺和亞精胺的前體物質(zhì),而且會(huì)加強(qiáng)組胺、酪胺的毒性作用,并且可以與亞硝酸鹽反應(yīng)生成致癌物質(zhì)亞硝胺[2],攝入過(guò)量可能造成低血壓、四肢疫攣等,2016年Gardini等再次提到高濃度的腐胺可促進(jìn)CT-26結(jié)腸腫瘤細(xì)胞生長(zhǎng),與腫瘤的發(fā)展相關(guān)[3]。
胍基丁胺脫亞胺酶(Agmatine deiminase,AGDI)途徑是微生物合成腐胺的一個(gè)主要途徑,其中精氨酸先脫羧基形成胍基丁胺,然后去亞氨基化成腐胺[4]。負(fù)責(zé)AGDI途徑的基因束包括調(diào)節(jié)基因aguR,分解代謝基因aguB、aguD、aguA和aguC[5-6]共同形成的操縱子aguBDAC。aguR編碼的AguR是一個(gè)跨膜單組分信號(hào)轉(zhuǎn)導(dǎo)蛋白,它能感受到細(xì)胞外的胍基丁胺并啟動(dòng)aguBDAC的轉(zhuǎn)錄從而生合成腐胺[7]。
NaCl會(huì)抑制細(xì)菌的生長(zhǎng)[8],影響微生物的新陳代謝,而且能夠破壞細(xì)菌細(xì)胞膜上參與生物胺合成的酶。許多研究表明,NaCl會(huì)影響編碼生物胺合成相關(guān)酶的基因表達(dá),從而抑制生物胺的產(chǎn)生,如La Gioia等[9]發(fā)現(xiàn)NaCl會(huì)影響嗜熱鏈球菌中編碼酪氨酸脫羧酶的tdcA基因,Rossi等[10]發(fā)現(xiàn)NaCl影響嗜熱鏈球菌中編碼組氨酸脫羧酶的hdcA基因。熏馬腸是深受新疆哈薩克族人民喜愛(ài)的發(fā)酵肉制品,它依靠特有地理環(huán)境所擁有的非人工菌種經(jīng)過(guò)一個(gè)月以上的時(shí)間自然發(fā)酵成熟,期間由于諸多不可控因素導(dǎo)致熏馬腸中存在許多雜菌,這些雜菌中不乏產(chǎn)胺微生物[11]。本實(shí)驗(yàn)使用的德氏乳桿菌是從熏馬腸中分離得到的產(chǎn)腐胺菌,目前國(guó)內(nèi)外對(duì)于NaCl對(duì)德氏乳桿菌產(chǎn)腐胺轉(zhuǎn)錄調(diào)控的影響情況及其原因鮮見(jiàn)報(bào)道,故本實(shí)驗(yàn)從分子生物學(xué)角度,研究了NaCl濃度對(duì)供試菌參與AGDI途徑產(chǎn)腐胺的調(diào)節(jié)基因aguR和操縱子aguBDAC的影響,將反轉(zhuǎn)錄實(shí)時(shí)熒光定量技術(shù)與超高效液相色譜技術(shù)相結(jié)合,分析基因束中調(diào)節(jié)子和操縱子基因的轉(zhuǎn)錄情況與腐胺積累的關(guān)系,明確了NaCl抑制細(xì)菌產(chǎn)生腐胺的機(jī)理,為發(fā)酵食品的安全性提供了理論基礎(chǔ)。
1 材料與方法
1.1 材料與儀器
NaCl 純度99.5%,天津市盛奧化學(xué)試劑有限公司;改良MRS培養(yǎng)基(半乳糖代替葡萄糖作為碳源) 北京奧博星生物試劑公司;胍基丁胺(純度97%)、丹磺酰氯(純度99%)、腐胺標(biāo)準(zhǔn)品(純度97%) 美國(guó)Sigma公司;甲醇、乙腈 均為色譜純,天津福晨化學(xué)試劑廠;Trizol 美國(guó)Invitrogen公司;cDNA合成試劑盒 北京全式金生物技術(shù)有限公司;Bestar? SybrGreen qPCR mastermix、ROX Reference Dye 德國(guó)DBI公司;供試菌:德氏乳桿菌(Lactobacillusdelbrueckii) 菌株號(hào)為MSR5-6a,GenBank登錄號(hào)為MF661779,由本實(shí)驗(yàn)室從熏馬腸中分離保存,最后送往華大基因測(cè)序所得。
LDZX-30KBS型立式壓力蒸汽滅菌鍋 上海申安醫(yī)療器械廠;EON型多功能酶標(biāo)儀 美國(guó)伯騰儀器有限公司;ND2000C型微量核酸測(cè)定儀 香港基因有限公司;5417R型高速冷凍離心機(jī) 德國(guó)艾本德公司;M×3000P型熒光定量PCR儀 美國(guó)安捷倫科技公司;H-Class型超高效液相色譜儀 美國(guó)沃特世公司。
1.2 實(shí)驗(yàn)方法
1.2.1 菌懸液的制備 用滅菌的接種環(huán)將在-18 ℃斜面保藏的德氏乳桿菌MSR5-6a挑取一環(huán),在MRS固體培養(yǎng)基中劃線,37 ℃恒溫培養(yǎng)24 h。挑取單菌落于MRS液體培養(yǎng)基中,37 ℃培養(yǎng)后將菌懸液稀釋至104~105CFU/mL,備用。
1.2.2 NaCl對(duì)供試菌生長(zhǎng)的抑制作用 在每支試管中移入5.0 mL含有0.4%胍基丁胺[12]與0.005%的磷酸吡哆醛的MRS液體培養(yǎng)基,然后分別加入0~6%的NaCl,滅菌冷卻后接入供試菌液。37 ℃恒溫培養(yǎng)48 h,每4 h取一次樣,采用多功能酶標(biāo)儀測(cè)定菌液的OD600值,使用含相應(yīng)NaCl濃度的空白培養(yǎng)基作對(duì)照,并用pH計(jì)測(cè)定pH。
采用平板菌落計(jì)數(shù)法測(cè)定活菌數(shù)。吸取1.0 mL待測(cè)菌液,按照1∶10稀釋到適宜的梯度,加入100 μL菌懸液在含1%~6% NaCl的MRS平板上,均勻涂布,另設(shè)一組不添加NaCl的培養(yǎng)基作為對(duì)照。37 ℃恒溫培養(yǎng)24~48 h,菌落總數(shù)不再顯著增加時(shí)進(jìn)行計(jì)數(shù)。每個(gè)處理取三個(gè)重復(fù),結(jié)果取平均值來(lái)計(jì)算菌落數(shù)和抑菌率。以未被抑制的德氏乳桿菌活菌總數(shù)為指標(biāo),不僅可以體現(xiàn)不同濃度NaCl對(duì)其數(shù)目的抑制效果,而且能觀察到被抑制后德氏乳桿菌的生長(zhǎng)情況。
抑菌率(%)=(CK組菌落數(shù)-處理組菌落數(shù))/CK組菌落數(shù)×100(其中,CK組為不含NaCl組,處理組分別為含1%~6% NaCl組)
花粉產(chǎn)生于花粉囊,是植物的雄性生殖細(xì)胞,功能與動(dòng)物精子相似。植物花粉的傳播途徑大致分為蟲(chóng)媒與風(fēng)媒,而引起廣大患者過(guò)敏的罪魁禍?zhǔn)桩?dāng)屬風(fēng)媒花。它們往往數(shù)量極大,可以憑借風(fēng)的力量擴(kuò)大傳播范圍。在遼寧地區(qū),主要致敏花粉有蒿屬、豚草、葎草等等。
1.2.3 RT-qPCR測(cè)AGDI相關(guān)基因表達(dá)量
1.2.3.1 RNA的提取 分別取1.2.2中0、1%、2%、3% NaCl濃度下(考慮到加入4%及以上濃度的NaCl時(shí)細(xì)菌增長(zhǎng)數(shù)量少,RNA提取困難,因此不提取)[13]生長(zhǎng)至穩(wěn)定期的菌液2.0 mL,于無(wú)酶離心管中離心得菌體,采用Trizol法[14]提取細(xì)菌總RNA。RNA提取完成后,使用微量核酸測(cè)定儀測(cè)定其濃度以及在260、280 nm的光吸收值,直至OD260/OD280的值為1.8~2.0。由1%瓊脂糖凝膠電泳檢測(cè)清晰可見(jiàn)23S rRNA、16S rRNA、5S rRNA三條條帶且無(wú)基因組污染,表明RNA的完整性和純度均符合反轉(zhuǎn)錄的要求,方可進(jìn)行下一步。
1.2.3.2 cDNA的合成 使用cDNA合成試劑盒將提取出的細(xì)菌總RNA反轉(zhuǎn)錄成cDNA,其中20μL體系中,總RNA添加量為500 ng。cDNA合成完畢后置于-80 ℃冰箱貯存?zhèn)溆谩?/p>
1.2.3.3 RT-qPCR對(duì)基因表達(dá)定量 RT-qPCR反應(yīng)體系制備如下:Bestar? SybrGreen qPCR mastermix 12.5 μL,上下引物各0.5 μL,cDNA模板1 μL,補(bǔ)充ddH2O至終體系為25 μL,加入ROX Reference Dye使其在整個(gè)體系的終濃度為0.1×,最后用微型離心機(jī)以6000 r/min的速度離心2 min以充分混勻反應(yīng)液。
反轉(zhuǎn)錄實(shí)時(shí)熒光定量所用引物見(jiàn)表1。其中aguR和aguB為目的基因,tufA和rpoA為內(nèi)參基因,每組基因各做三組平行。RT-qPCR擴(kuò)增程序如下:Stage 1:預(yù)變性 95 ℃ 2 min;Stage 2:PCR反應(yīng)40個(gè)循環(huán) 95 ℃ 10 s,55 ℃ 34 s,72 ℃ 30 s;Stage 3:融解曲線。以3% NaCl濃度下的基因表達(dá)量為對(duì)照,使用2-ΔΔCt的計(jì)算方法[16]計(jì)算其余NaCl濃度下的aguR和aguB基因表達(dá)量。

表1 反轉(zhuǎn)錄實(shí)時(shí)熒光定量所用引物
1.2.4 UPLC監(jiān)測(cè)菌液中腐胺含量
1.2.4.1 腐胺標(biāo)準(zhǔn)溶液的配制 準(zhǔn)確稱(chēng)取腐胺標(biāo)準(zhǔn)品5 mg,加入0.4 mol/L的高氯酸定容至5 mL,分別吸取25、50、100、125、250、500 μL于5 mL棕色容量瓶中,加入0.4 mol/L的高氯酸使終體積為1 mL,作為腐胺標(biāo)準(zhǔn)溶液備用。
1.2.4.2 樣品的制備 以1.2.2中0、1%、2%、3% NaCl濃度下的菌液為研究對(duì)象(考慮到加入4%及以上濃度的NaCl時(shí)細(xì)菌增長(zhǎng)數(shù)量少,腐胺幾乎不產(chǎn)生,因此不做研究)[13],每4 h取一次樣。每次取菌液后經(jīng)12000 r/min離心10 min,吸取上清液1 mL,加入等體積的0.4 mol/L高氯酸,充分混勻制成菌液樣品處理液備用。
1.2.4.4 色譜條件 ACQUITY UPLC? HSS T3柱(2.1 mm×50 mm,1.8 μm),流動(dòng)相A為水,流動(dòng)相B為乙腈;流速0.4 mL/min;每次進(jìn)樣量5 μL;柱溫30 ℃;選用波長(zhǎng)254 nm。梯度洗脫程序見(jiàn)表2。

表2 洗脫程序
1.2.5 數(shù)據(jù)分析與處理 所有實(shí)驗(yàn)數(shù)據(jù)使用Microsoft Excel 2013建立數(shù)據(jù)庫(kù),Origin 8.5 作圖,并用SPSS 23進(jìn)行差異性分析。
2 結(jié)果與分析
2.1 NaCl對(duì)供試菌株生長(zhǎng)的抑制作用
如圖1所示,與CK組相比,不同濃度NaCl中細(xì)菌的延滯期和對(duì)數(shù)期會(huì)不同程度地延長(zhǎng),1%~6% NaCl六組的細(xì)菌OD600始終低于CK組,說(shuō)明NaCl可有效抑制供試菌的生長(zhǎng),減少菌體數(shù)量,且濃度越大,抑制作用越大,但NaCl并沒(méi)有改變供試菌的生長(zhǎng)趨勢(shì),只是延緩了其穩(wěn)定期的到來(lái)。

圖1 NaCl對(duì)德氏乳桿菌生長(zhǎng)的抑制作用
各組供試菌在MRS固體培養(yǎng)基生長(zhǎng)48 h后,通過(guò)計(jì)算不同NaCl濃度下的抑菌率,發(fā)現(xiàn)增大NaCl的濃度對(duì)德氏乳桿菌抑制效果顯著(p<0.05)增強(qiáng)(圖2)。說(shuō)明NaCl在細(xì)菌細(xì)胞中具有良好的滲透性,且分散效果良好,能夠使細(xì)胞內(nèi)的水分滲出從而有效抑制細(xì)菌生長(zhǎng)。1% NaCl組的抑菌率為5.79%,說(shuō)明此濃度的NaCl對(duì)德氏乳桿菌抑制作用較小,而6% NaCl組的抑菌率為96.71%,說(shuō)明此濃度下供試菌基本無(wú)活性。通過(guò)測(cè)定微生物的生長(zhǎng)情況,初步確定了不同濃度NaCl對(duì)德氏乳桿菌具有顯著的抑制效果。

圖2 NaCl對(duì)供試菌的抑菌率(IR)
2.2 不同濃度NaCl對(duì)菌株生長(zhǎng)過(guò)程中pH的影響
由圖3可知,從接入供試菌開(kāi)始,CK組的pH相對(duì)于其它組的下降更為迅速,1%~6% NaCl六組的pH趨于穩(wěn)定后均高于CK組,原因是NaCl抑制了德氏乳桿菌的增長(zhǎng),所以1%~6% NaCl六組相比于CK組培養(yǎng)液中所含的有機(jī)酸較少。3% NaCl組與CK、1% NaCl兩組pH變化曲線的趨勢(shì)不同,CK與1% NaCl兩組的pH先快速下降再緩慢下降最后平緩,而3% NaCl組pH先緩慢下降再快速下降,繼而趨于平緩,可能是因?yàn)?%濃度的NaCl顯著(p<0.05)抑制了供試菌的增長(zhǎng),所以從接入供試菌開(kāi)始,細(xì)菌增長(zhǎng)緩慢,細(xì)菌數(shù)量少產(chǎn)酸少導(dǎo)致pH下降緩慢,至32 h左右細(xì)菌總數(shù)達(dá)到最高峰,所以3% NaCl組在32 h左右進(jìn)入穩(wěn)定期時(shí),pH依舊下降較快。

圖3 不同NaCl濃度下德氏乳桿菌生長(zhǎng)過(guò)程中pH的變化
2.3 不同NaCl濃度下的aguR基因表達(dá)
圖4中,德氏乳桿菌MSR5-6a在0、1%、2%濃度的NaCl下aguR基因的表達(dá)量分別為1.62、1.18、1.49,均不到3% NaCl組的兩倍,說(shuō)明不同濃度NaCl情況下aguR基因表達(dá)量差別不大,即AGDI的調(diào)節(jié)基因aguR的轉(zhuǎn)錄不受NaCl濃度的影響,這與Del等[13]對(duì)于乳酸乳球菌的研究結(jié)果相近。AGDI基因束第一個(gè)基因編碼的AguR是一種跨膜蛋白,它作為信號(hào)轉(zhuǎn)導(dǎo)系統(tǒng)的一部分可以感受到培養(yǎng)基中的胍基丁胺濃度從而調(diào)節(jié)操縱子aguBDAC的轉(zhuǎn)錄[17]。也許是因?yàn)閍guR通常表達(dá)量很低,目前的研究表明它不受胍基丁胺濃度[7]、葡萄糖濃度[12]、培養(yǎng)基pH[18]的影響,這確保位于細(xì)胞表面的AguR可以充當(dāng)胍基丁胺感應(yīng)器。
2.4 AguB基因表達(dá)情況分析
圖5顯示,CK組的aguB基因表達(dá)量是3% NaCl組的292倍,1% NaCl組的aguB基因表達(dá)量是3% NaCl組的118.6倍,2% NaCl組的則是3% NaCl組的19.1倍,說(shuō)明添加NaCl能夠影響aguB的轉(zhuǎn)錄活性。NaCl極顯著(p<0.01)減少了操縱子aguBDAC的轉(zhuǎn)錄,且NaCl濃度越大,影響越大。可能是由于胍基丁胺可以導(dǎo)致AguR的二聚化,從而通過(guò)結(jié)合羧基上的C使啟動(dòng)子aguB促進(jìn)分解代謝基因的轉(zhuǎn)錄繼而產(chǎn)生腐胺[7]。而NaCl影響了AguR對(duì)于胍基丁胺的感應(yīng)和隨后的二聚化,從而影響了的aguB的啟動(dòng)。而且AGDI途徑中產(chǎn)生的能量也會(huì)被用來(lái)抵消NaCl引起的滲透壓失衡,所以添加NaCl不利于AGDI反應(yīng)的發(fā)生。

圖5 不同NaCl濃度下的aguB基因表達(dá)量
有研究表明,pH會(huì)影響胍基丁胺脫亞胺酶基因的表達(dá)[19],Griswold等[20]證明胍基丁胺脫亞胺酶的基因表達(dá)量在酸性環(huán)境比在中性和堿性環(huán)境中更高。Rio等[18]發(fā)現(xiàn)乳酸乳球菌的AGDI基因束受pH影響,酸性pH可能有利于AguR的構(gòu)造[21],使得信號(hào)轉(zhuǎn)導(dǎo)到DNA結(jié)合域更為高效,從而激活aguB的信號(hào)后增加了操縱子aguBDAC的轉(zhuǎn)錄。在其它乳酸菌中也發(fā)現(xiàn)酸性pH有利于其它產(chǎn)生物胺途徑的轉(zhuǎn)錄活性,如Linares等[22]和Perez等[23]證明,酸性pH有益于耐久腸球菌和糞腸球菌中的酪胺合成。由圖3可知,CK組和1% NaCl組供試菌生長(zhǎng)到達(dá)穩(wěn)定期時(shí)pH相差不大,但圖5中CK組的aguB基因表達(dá)量是1% NaCl組的兩倍多,說(shuō)明在不受pH的影響時(shí),NaCl能夠極顯著(p<0.01)抑制德氏乳桿菌AGDI途徑中aguB基因的表達(dá)。而CK組的aguB基因表達(dá)量是3% NaCl組的292倍,且由圖3可知,處于穩(wěn)定期時(shí)兩者pH相差接近1,說(shuō)明CK組和3% NaCl組aguB基因表達(dá)量的差異既有NaCl濃度的影響,又有NaCl濃度不同導(dǎo)致的培養(yǎng)液中pH不同對(duì)aguB基因表達(dá)所造成的干擾。
2.5 腐胺UPLC色譜圖與標(biāo)準(zhǔn)曲線
如圖6所示,腐胺出峰時(shí)間在7.45 min左右,圖7中,腐胺標(biāo)準(zhǔn)曲線的回歸方程為y=16850x+115452,R2=0.99957>0.999,說(shuō)明標(biāo)準(zhǔn)曲線的線性關(guān)系良好,符合測(cè)樣要求。
圖6 腐胺標(biāo)準(zhǔn)品UPLC色譜圖

圖7 腐胺標(biāo)準(zhǔn)曲線
2.6 不同NaCl濃度下供試菌產(chǎn)腐胺量分析
如圖8所示,從12 h檢測(cè)到腐胺開(kāi)始,CK組的腐胺積累量始終大于其它組;12~36 h之間CK與1% NaCl組的腐胺積累速率顯著(p<0.05)大于2% NaCl與3% NaCl組;40 h左右,各組的腐胺積累速率開(kāi)始減慢。培養(yǎng)液中添加了足量的胍基丁胺與磷酸吡哆醛,隨著供試菌培養(yǎng)時(shí)間的增加,可在培養(yǎng)液中檢測(cè)到一定量的腐胺,說(shuō)明德氏乳桿菌具有較強(qiáng)的通過(guò)AGDI途徑產(chǎn)腐胺的能力。增加NaCl的濃度可以有效減少生物胺的積累[24]。NaCl可以減少細(xì)菌的生長(zhǎng),破壞位于細(xì)菌細(xì)胞膜上的氨基酸脫羧酶,從而減少生物胺的產(chǎn)生[25]。圖1中,隨著NaCl濃度的增加,細(xì)菌增長(zhǎng)越來(lái)越慢,圖8中,隨著NaCl濃度的增加,培養(yǎng)液中腐胺的含量變小,可見(jiàn)NaCl對(duì)于德氏乳桿菌增長(zhǎng)的抑制作用減少了腐胺的產(chǎn)生。
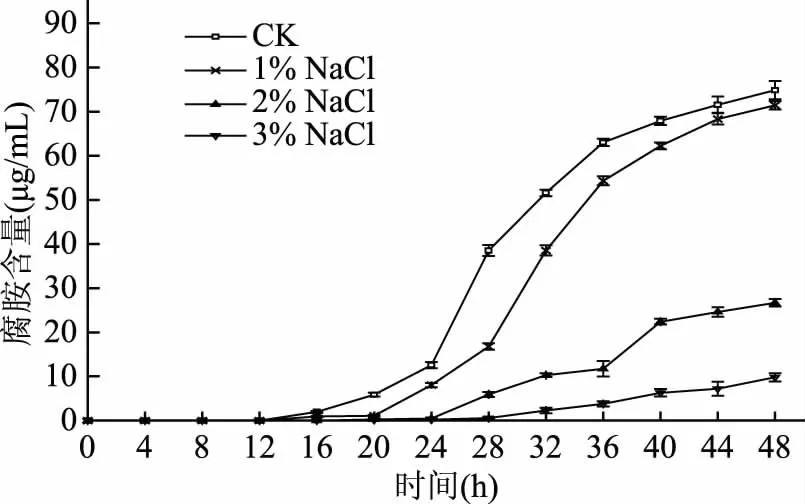
圖8 0、1%、2%、3% NaCl濃度下的腐胺產(chǎn)生量
供試菌生長(zhǎng)代謝過(guò)程中產(chǎn)生的乳酸不斷積累,pH連續(xù)下降,腐胺積累量也慢慢增多。研究表明,乳酸菌生長(zhǎng)過(guò)程中產(chǎn)酸所造成的酸性外部環(huán)境能導(dǎo)致產(chǎn)生更多的生物胺[26-27],Griswold等[20]也證明酸性pH能夠促進(jìn)變形鏈球菌產(chǎn)生腐胺。AGDI途徑中的胍基丁胺代謝除了產(chǎn)生ATP外也會(huì)產(chǎn)生銨離子,ATP的產(chǎn)生是細(xì)菌增長(zhǎng)的主要因素,還可以促進(jìn)質(zhì)子的排出[6],而細(xì)胞外介質(zhì)中銨離子的積累可以導(dǎo)致堿化。Alberto等[28]和Suárez等[29]發(fā)現(xiàn)銨離子導(dǎo)致的堿化有利于微生物的耐酸性。由此我們可以推斷,德氏乳桿菌MSR5-6a在生長(zhǎng)過(guò)程中產(chǎn)酸可以促進(jìn)AGDI反應(yīng)的發(fā)生,這可能就是一種耐酸反應(yīng),可以改善微生物在低pH環(huán)境中的適應(yīng)性,這一點(diǎn)在短乳桿菌[29]、糞腸球菌[30]、變形鏈球菌[20-21]中已經(jīng)得到證明。圖8中CK組在剛開(kāi)始的12 h內(nèi)培養(yǎng)液中基本沒(méi)有檢測(cè)到腐胺,這一段時(shí)間可能微生物在增加數(shù)量的同時(shí)也在對(duì)環(huán)境產(chǎn)生適應(yīng)性。
CK和1% NaCl組在24 h后才開(kāi)始大量檢測(cè)到腐胺,2% NaCl和3% NaCl組在28 h左右檢測(cè)到腐胺。這一現(xiàn)象可能與Del等[13]研究發(fā)現(xiàn)的AGDI基因束的調(diào)節(jié)基因AguR和啟動(dòng)子PaguB只有在微生物生長(zhǎng)到對(duì)數(shù)期和穩(wěn)定期的過(guò)渡期時(shí)才會(huì)被激活有關(guān)。而后期腐胺的產(chǎn)生速率越來(lái)越慢是因?yàn)殡S著供試菌對(duì)胍基丁胺的消耗,培養(yǎng)液中胍基丁胺的濃度變低,胍基丁胺脫亞胺酶活性降低,從而腐胺的產(chǎn)生減緩[31]。隨著NaCl濃度的增加,操縱子aguBDAC的轉(zhuǎn)錄受到抑制,所以圖8中CK組比其它組最終的腐胺含量都高,也就是說(shuō)減少培養(yǎng)液中NaCl的濃度有利于胍基丁胺脫亞胺酶基因的表達(dá),從而增加腐胺的積累。
3 結(jié)論
本研究表明,在培養(yǎng)液中添加NaCl,能夠顯著(p<0.05)抑制德氏乳桿菌的生長(zhǎng),且NaCl濃度越高,抑制作用越大。NaCl對(duì)德氏乳桿菌AGDI基因束中的調(diào)節(jié)基因aguR的轉(zhuǎn)錄影響不大,但能抑制分解代謝基因aguBDAC中aguB基因的表達(dá)。其中,CK組的基因aguB的表達(dá)量高達(dá)3% NaCl組的292倍,隨著NaCl濃度的增加,AGDI基因束中的aguB基因表達(dá)量極顯著(p<0.01)減少,從而整個(gè)操縱子的轉(zhuǎn)錄受到抑制。即添加NaCl能夠通過(guò)抑制胍基丁胺脫亞胺酶的基因表達(dá),降低供試菌通過(guò)AGDI途徑產(chǎn)腐胺的能力,而供試菌具有較強(qiáng)的通過(guò)胍基丁胺脫亞胺酶途徑產(chǎn)腐胺的能力,所以增加培養(yǎng)液中NaCl的濃度能夠顯著(p<0.05)減少腐胺的產(chǎn)生。因此,在熏馬腸等發(fā)酵香腸制作過(guò)程中添加適當(dāng)濃度的NaCl,可有效減少后期發(fā)酵過(guò)程中腐胺的積累。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14

- 食品工業(yè)科技的其它文章
- 葉類(lèi)蔬菜貯藏中品質(zhì)變化評(píng)價(jià)指標(biāo)及其分析方法的研究進(jìn)展
- 燕麥生物堿的結(jié)構(gòu)與功能研究進(jìn)展
- 植物乳桿菌發(fā)酵乳對(duì)小鼠降膽固醇作用
- 天然非營(yíng)養(yǎng)型甜味料發(fā)酵乳對(duì)四氧嘧啶誘導(dǎo)糖尿病模型大鼠糖代謝的影響
- Lactobacillus plantarum YS-2對(duì)葡聚糖硫酸鈉誘導(dǎo)C57BL/6J小鼠結(jié)腸炎的抑制作用
- 基于電子鼻技術(shù)研究殼聚糖/ε-聚賴(lài)氨酸復(fù)合膜對(duì)中國(guó)對(duì)蝦的保鮮效果