棉花基因組中SABATH甲基轉移酶基因家族的查找與分析
2018-10-11 06:23:20狄佳春陳旭升
江蘇農業科學 2018年17期
趙 亮, 狄佳春, 陳旭升
(江蘇省農業科學院經濟作物研究所/農業部長江下游棉花與油菜重點實驗室,江蘇南京 210014
甲基化作用廣泛存在于生物體內,由甲基轉移酶(methyltransferases,簡稱MTs)依賴S-腺苷-L-甲硫氨酸(S-adenosine-L-methionine,簡稱SAM)作為甲基供體催化形成相對應的甲基化形式產物[1]。在植物次生代謝中甲基化是一個非常普通的酶修飾反應,幾乎所有的植物次生代謝途徑都會發生甲基化,例如氨基酸、生物堿、苯丙素類、糖、嘌呤、甾醇類、硫醇類和黃酮類。甲基轉移酶可根據甲基轉移到受體分子上不同的原子而分為C、N、S和O位甲基轉移酶[2]。
SABATH甲基轉移酶是屬于O位甲基轉移酶(O-methyltransferase,簡稱OMTs),該家族催化一些小分子羧基和生物堿的甲基化,這個家族在最初發現鑒定的3個酶的基礎上共同命名為SABATH甲基轉移酶(SABATH-MTs)。這3個酶分別是水楊酸羧基甲基轉移酶(salicylic acid carboxyl methyltransferases,簡稱SAMT)、苯甲酸羧基甲基轉移酶(benzoic acid carboxyl methyltransferases,簡稱BAMT)和可可堿合酶(theobromine synthase)[3-7]。對SABATHMTs基因序列分析表明,在擬南芥中存在24個該家族基因成員。功能研究清楚的有茉莉酸羧基甲基轉移酶(jasmonic acid carboxyl methyltransferase,簡稱JMT)[8]、苯甲酸/水楊酸羧基甲基轉移酶(benzoic/salicylic acid carboxyl methyltransferases,簡稱BSMT)[9]、吲哚乙酸羧基甲基轉移酶(indole-acetic acid carboxyl methyltransferase,簡稱IAMT)[10]、法尼烯酸羧基甲基轉移酶(farnesoic acid carboxyl methyltransferase,簡稱FAMT)[11]、赤霉素羧基甲基轉移酶(gibberellin acid carboxyl methyltransferase,簡稱GAMT)[12]。棉花基因組數據的公開發表,為SABARH-MTs基因家族的全基因組分析提供了便利的條件。本研究利用生物信息學的方法,從棉花基因組數據庫中查找可能的SABATH-MTs基因家族成員,并對這些成員進行結構、進化及表達上的分析。
1 材料與方法
1.1 數據庫來源
陸地棉遺傳標準系TM-1的基因組數據庫來源于http://mascotton.njau.edu.cn;棉花二倍體D基因組雷蒙德氏棉(G.Raimondii)的基因組數據庫來源于http://www.phytozome.net;棉花二倍體A基因組亞洲棉的基因組數據來源于http://cgp.genomics.org.cn/page/species/index.jsp;擬南芥GAMT基因來源于擬南芥信息資源(the arabidopsis information resource,簡稱TAIR)數據庫(http://www.arabidopsis.org)。
1.2 基因注釋
GAMT注釋根據Sonnhammer等[13]的方法,HMMER軟件版本為3.0;GAMT種子文件來自于蛋白質家族的集合數據庫(Pfam數據庫,http://pfam.sanger.ac.uk/),種子編號為PF03492。蛋白質的氨基酸序列保守結構域的分析采用在線的SMART(http://smart.embl-heidelberg.de)進行檢測。
1.3 序列比對及進化樹構建
利用MEGA 5.05[14]構建進化樹,構建方法利用最大似然函數,Bootstrap實行1 000次重復。序列比對利用 Clastal 1.83 完成,比對結果及處理利用GENDOC軟件來完成。
1.4 基因表達分析
基因表達分析采用高通量的陸地棉遺傳標準系TM-1營養組織(根、莖、葉)、花瓣、花藥及纖維發育時期[開花當天(0 day post-anthesis,簡稱0 DPA)、開花后10天(10 DPA)、開花后20天(20DPA)]的RNA-seq數據庫[15]。基因表達水平利用Log2(FPKM)的數值進行評估,其結果利用Cufflinks軟件(http://cufflinks.cbcb.umd.edu)進行處理。
2 結果與分析
2.1 基因組數據庫中SABATH-MTs基因的注釋
由于SABATHMTs家族基因的蛋白結構中含有一個Methyltransf_7的蛋白結構域,該結構域在Pfam數據庫中的種子文件編號為PF03492。通過HMMER 3.0中的hmmsearch程序及種子文件,在陸地棉遺傳標準系TM-1基因組中共查找到37個基因,其中At亞組中有16個基因,Dt亞組中有19個,還有2個基因的染色體信息不清楚;二倍體D基因組雷蒙德氏棉中查找到22個基因;二倍體A基因組亞洲棉中查找到24個基因,這些基因在染色體上呈現不均勻的分布。以陸地棉TM-1為例,在查找到的37個基因不均勻的分布在19條染色體上,其中9條染色體屬于At亞組,10條染色體屬于Dt亞組;A10、D10和D13上的分布的基因最多,分別預測到4個基因(圖1)。
2.2 SABATH-MTs基因系統進化分析
根據已經報道具有已知功能的SABATH-MTs基因,從美國國立生物技術信息中心(national center for biotechnology information,簡稱NCBI)上下載其對應的蛋白序列與棉花基因組中預測的基因進行系統進化分析。結果表明,在總計預測到的83個基因(四倍體中有37個;二倍體D組中有22個;二倍體A組中有24個)中,有34個基因能夠與已知功能的SABATH-MTs基因聚到同一分支(圖2)。這34個基因中有8個基因來自于二倍體A組,9個基因來自于二倍體D組;17個基因包括At亞組9個、Dt亞組8個。進一步分析表明,來源二倍體A組和D組的基因在四倍體中都能夠找到其相應的同源基因(表1)。
2.3 SABATH-MTs基因表達分析
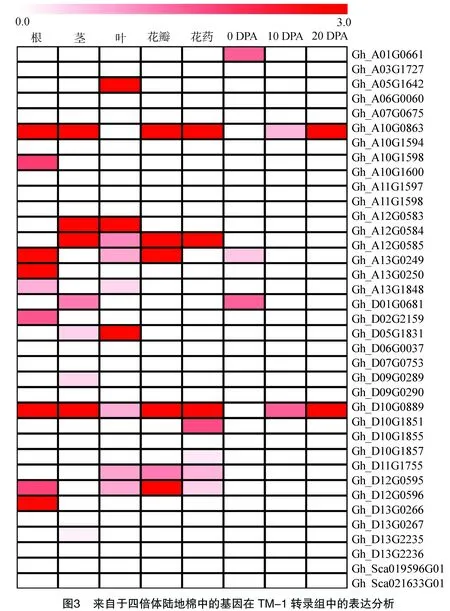
在陸地棉TM-1中預測到的37個SABATH-MTs基因在其根、莖、葉、花瓣、花藥及3個纖維發育時期(0、10、20 DPA)的轉錄組中的表達分析表明,共有20個基因在至少1個轉錄組數據庫中能夠檢測到表達[Log2(FPKM)>1]。其中有10個基因只在1個轉錄組數據庫中能夠檢測到表達。Gh_D02G2159、Gh_A10G1598、Gh_A13G0249和Gh_D13G0266只在根的轉錄組中檢測到有表達;只在莖的轉錄組中檢測有表達的為Gh_D13G2235和Gh_D09G0289;Gh_A05G1642在葉的轉錄組中專化表達;Gh_D10G1851和Gh_D11G1755在花藥的轉錄組中專化表達;Gh_A01G0661在0 DPA的胚珠中專化表達。花瓣、10 DPA和20 DPA的纖維中沒有找到專化表達的基因(圖3)。
3 結論與討論
甲基化在許多生物進程中發揮著關鍵的修飾作用,它包括基因和代謝的調控。在植物中,甲基化也有著廣泛的作用。與其他已知的甲基轉移酶比較發現,SABATH家族在幾個方面都是獨特的。首先,除了保守的SAM結合序列,SABATH-MTs與其他MTs沒有序列的相似性[2];再者,SABATH基因家族的蛋白能夠催化O位和N位2個位置的甲基化,而其他的MTs只能執行1種類型的甲基化[3,5,7]。擬南芥基因組中包含24個SABATH家族蛋白成員,其中一些蛋白具有明顯的特征。例如,茉莉酸羧基甲基轉移酶(JMT)、苯甲酸嘌呤/水楊酸羧基甲基轉移酶(BSMT)、吲哚乙酸甲基轉移酶(IAMT)、法尼烯酸羧基甲基轉移酶(FAMT)及赤霉素羧基甲基轉移酶(GAMT)等[8-9,11-12,16-18]。而根據生物信息學的分析,在四倍體陸地棉中預測了37個SABATH-MTs家族成員;在二倍體亞洲棉中預測了24個;在二倍體雷蒙德氏棉中預測了22個。這些家族成員通過進化分析,有部分成員能夠與已經報道的具有特定功能的SABATH-MTs基因聚集到同一分支。四倍體陸地棉TM-1中預測到的37個基因中,有20個基因能夠在轉錄組數據庫中檢測到表達,另外的17個基因未能檢測到表達。這些未能檢測到表達的基因有可能是以假基因的形式存在于基因組中, 或者在其他組織部位及纖維發育時期專化表達而未能檢測到。通過進化及表達分析的結果,為今后對于棉花中SABATH-MTs家族成員的功能研究起到了一定的指導作用。
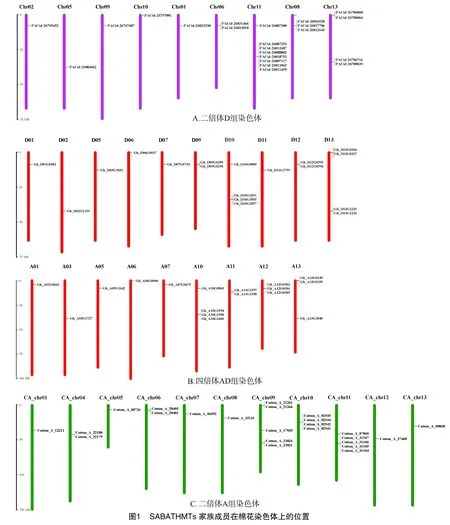

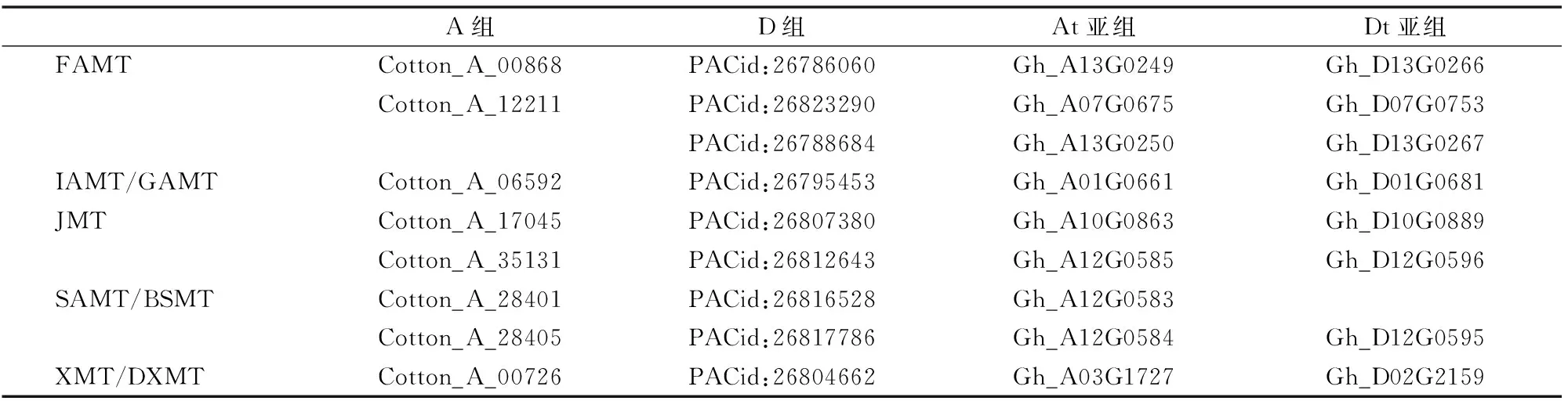
表1 通過進化分析具有特定功能的SABATH-MTs基因
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
財經(2017年15期)2017-07-03 22:40:49
財經(2017年2期)2017-03-10 14:35:35
山東工業技術(2016年15期)2016-12-01 05:31:22
財經(2016年15期)2016-06-03 07:38:02
財經(2016年3期)2016-03-07 07:44:46
財經(2016年6期)2016-02-24 07:41:51
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
