豬lncRNA-ENSSSCT00000018610的克隆及其在豬卵泡中的表達
2018-10-15 09:35:48李夢尋馬力鵬公紅斌邱梅玉孫曉梅
畜牧獸醫學報 2018年9期
關鍵詞:梅山
李夢尋,黃 濤,馬力鵬,劉 乙,李 濤,公紅斌,邱梅玉,謝 蘇,孫曉梅
(石河子大學動物科技學院,石河子 832000)
非編碼RNA(non-coding RNA, ncRNA)是指一大類不編碼蛋白質,卻具有生物活性的RNA[1],這類RNA曾一度被認為是轉錄的“暗物質”或“噪音”[2-3],但是隨著研究的不斷深入,越來越多的研究者證實了非編碼RNA在轉錄水平上發揮著廣泛的生物學作用[4]。非編碼RNA根據鏈的長度可劃分為短鏈非編碼RNA和長鏈非編碼RNA(lncRNA)。lncRNA是非編碼RNA中重要的組成部分,其長度一般大于200 nt[5]。目前,根據lncRNA及其相關蛋白編碼基因之間的關系將其分為反義lncRNA(antisense lncRNA)、雙向lncRNA(bidirectional lncRNA)、內含子lncRNA(intronic lncRNA)、正義重疊區lncRNA(sense overlapping lncRNA)和基因間隔區(lincRNA)與加工轉錄本[6-8];根據功能,lncRNA可分為信號因子、引導因子、誘餌因子和支架因子[9]。不同類型的lncRNAs具有不同的作用。目前lncRNA在動物繁殖方面的生物學研究較少。lncRNA在動物早期生殖細胞形成、早期胚胎的著床和發育以及有關激素調節中扮演著必不可少的角色[10]。科學家們通過RNA-Seq對人、小鼠、牛等受精卵與受精前的配子、不同發育階段的早期胚胎間的比較,發現大量與不同胚胎發育時期相關的lncRNAs[11-12],如Meg3與JARID2基因互作從而招募PRC2以反式作用抑制胚胎發育相關基因表達[13]。Chen等[14]在小鼠胚胎發育不同階段的卵巢中發現了24個lncRNA差異表達,在相同胎齡的雌雄生殖器官中發現147個lncRNA差異表達,在卵巢中表達最高的oncRNA3是泛酸激酶1。Roeszler等[15]研究發現,在母雞沉默lnc-MHM會造成卵巢不對稱發育。Brown等[16]在對果蠅和小鼠卵巢lncRNA研究中發現了大量與啟動子相關的反義lncRNAs,這些lncRNAs可能調控其同源基因的轉錄激活。Macaulay等[17]利用共聚焦透射電子顯微鏡和RNA-Seq發現,牛卵母細胞周圍的卵丘細胞為成年母牛的卵母細胞運輸大量的營養和物質,包括mRNA和lncRNA,由于卵母細胞及其周圍細胞細胞質間的信息傳遞和物質運輸方式有限,差異表達的lncRNA可能對卵子的發生具有重要作用。
目前在豬卵泡發育上,對于基因表達調控的研究主要集中在蛋白質編碼基因上,而對參與豬卵泡發育的非編碼RNA的研究相對較少,這遺漏了大量的遺傳信息。本課題組前期通過高通量測序技術獲得了梅山和杜洛克豬中等卵泡lncRNA 3 554個,其中已注釋的為1 997個,未注釋的為1 557個[18]。lncRNA-ENSSSCT00000018610是從梅山豬和杜洛克豬中等卵泡差異lncRNA數據庫中獲得的已注釋長鏈非編碼RNA。
為了對lncRNA-ENSSSCT00000018610進行深入的研究,本試驗采用3′和5′ Rapid Amplification of cDNA Ends (RACE)技術擴增得到其全長序列,利用Real-time PCR對其進行組織表達譜分析,并對其在梅山豬和杜洛克豬的各級卵泡中的相對表達量進行檢測,揭示了其在兩個豬種不同組織和各級卵泡中的表達規律,有助于探究該lncRNA在卵泡發育過程中的作用機理,為深入研究豬種間排卵數差異的分子機理提供了科學依據。
1 材料與方法
1.1 試驗樣品采集
試驗選取同一胎次、發情正常、繁殖表現符合品種特征的梅山和杜洛克成年母豬各3頭,相同條件下飼喂,每天對其發情狀況進行觀測,以發情出現靜立反應時記做發情周期第0天,發情周期第14天沿耳緣靜脈注射獸用氯前列烯醇注射液1支·頭-1,注射后第4天屠宰,取不同級別卵泡(按直徑分類,S卵泡:1.0~3.0 mm;M1卵泡:3.1~5.0 mm;M2卵泡:5.1~7.0 mm;L卵泡:7.0 mm以上)于液氮中凍存,同時取卵巢、下丘腦、心、肝、脾、肺、腎、小腸、肌肉、子宮、垂體、黃體、輸卵管,用錫紙包裹,液氮速凍后于-80 ℃保存備用。
1.2 主要試劑與儀器
主要試劑:TRIzol試劑(Invitrogen,Carlsbad,CA,USA);反轉錄試劑盒PrimerScriptTM RT regagent Kit(TaKaRa);LightCycler 480 SYBR Green、八連管(Roche);SMARTer?RACE cDNA Amplification Kit、SeqAmp DNA Polymerse、NucleoSpin Gel and PCR Clean-Up Kit、In-Fusion HD Cloning Kit、Stellar Competent Cells(Clontech);DNA Maker、西班牙進口Agarose瓊脂糖、營養瓊脂、胰蛋白胨、酵母提取物(北京天根)等。
主要儀器:超低溫冰箱(-80 ℃)、恒溫水浴鍋、高壓滅菌鍋、移液槍、PCR儀、超凈工作臺、NanoDrop 2000、LightCycler 480實時熒光定量PCR儀、分析天平、高速離心機、電熱恒溫培養箱、水平電泳槽、電泳儀;GelDoc X R型凝膠成像系統等。
1.3 lncRNA-ENSSSCT00000018610保守性分析和編碼潛能預測
根據lncRNA-ENSSSCT00000018610在Ensembl上注釋的序列信息,采用Phast Cons Score工具對lncRNA進行保守性評估,并用Coding-Potential Assessment Tool(CAPT)[19]對lncRNA-ENSSSCT00000018610編碼能力進行預測。
1.4 引物設計與合成
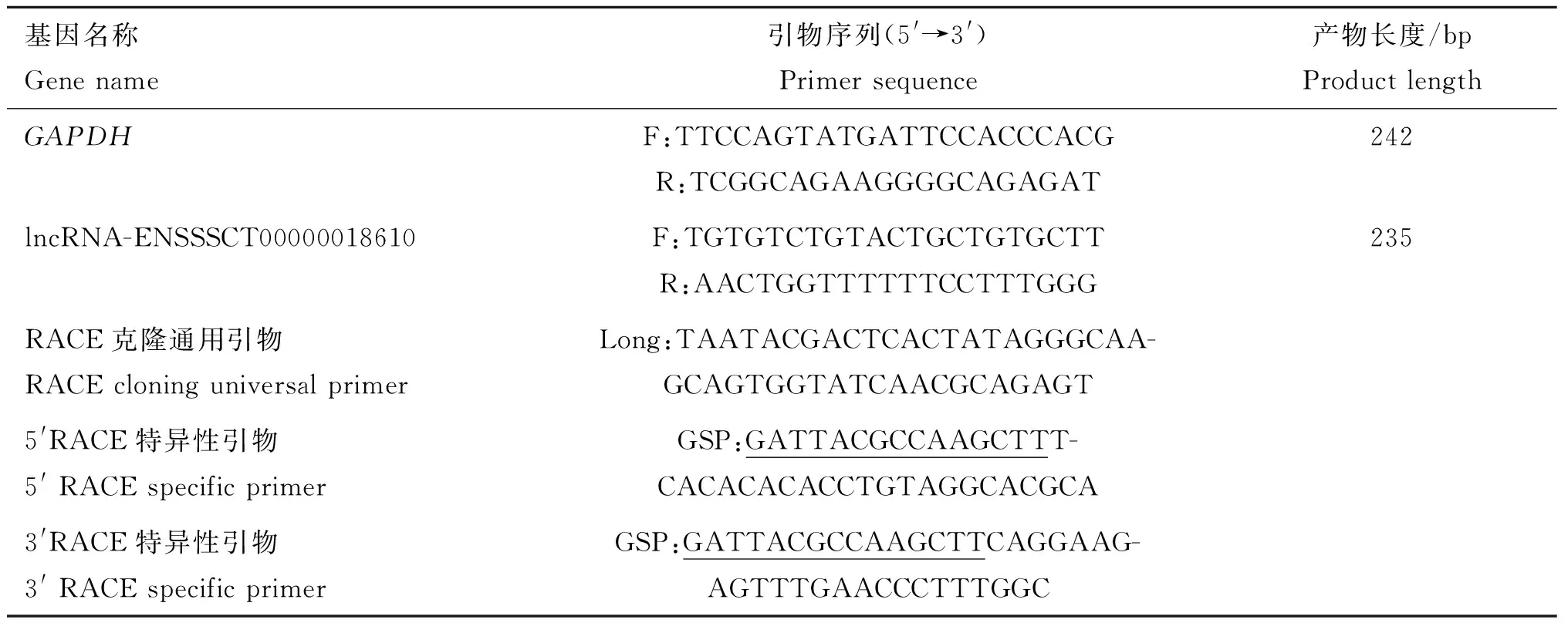
根據lncRNA-ENSSSCT00000018610部分已知核苷酸序列,用Primer Primer 5.0軟件設計引物,分別用于實時熒光定量PCR、5′和3′ RACE PCR擴增,用于RACE克隆的特異性引物在5′末端添加15 bp序列“GATTACGCCAAGCTT”用于后續試驗的連接測序,所有試驗所需引物見表1,引物序列由新疆烏魯木齊市昆泰銳公司合成。
表1lncRNA-ENSSSCT00000018610及內參引物序列
Table1TheprimersforlncRNA-ENSSSCT00000018610andinternalreferencegene
基因名稱Gene name引物序列(5'→3')Primer sequence產物長度/bpProduct lengthGAPDHF:TTCCAGTATGATTCCACCCACGR:TCGGCAGAAGGGGCAGAGAT242lncRNA-ENSSSCT00000018610F:TGTGTCTGTACTGCTGTGCTTR:AACTGGTTTTTTCCTTTGGG235RACE克隆通用引物RACE cloning universal primerLong:TAATACGACTCACTATAGGGCAA-GCAGTGGTATCAACGCAGAGT5'RACE特異性引物5' RACE specific primerGSP:GATTACGCCAAGCTTT-CACACACACCTGTAGGCACGCA3'RACE特異性引物3' RACE specific primerGSP:GATTACGCCAAGCTTCAGGAAG-AGTTTGAACCCTTTGGC
表中5′ RACE和3′ RACE基因特異性引物中的下劃線序列為添加的序列,用于后續產物與pUC 19線性載體連接測序;無下劃線序列為基因引物
The sequences with underline in the 5′ RACE and 3′ RACE gene-specific primers are added sequences that are sequenced with subsequent products and pUC 19 linearized vectors; the sequences without underline are primers for genes
1.5 總RNA的提取與cDNA合成
取兩品種豬的各級卵泡和不同組織100 mg于液氮中研磨,TRIzol法分別提取樣本總RNA,用2%瓊脂糖凝膠電泳和NanoDrop 2000驗證總RNA的完整性、濃度及純度。-80 ℃保存備用。實時熒光定量PCR所用cDNA的合成分別按照TaKaRa反轉錄試劑盒說明書進行操作,5′和3′ RACE PCR擴增參照SMARTer?RACE cDNA第一鏈合成說明書進行操作,獲得的cDNA置于-20 ℃保存備用。
1.6 lncRNA-ENSSSCT00000018610全長克隆
5′ RACE PCR反應:試驗以5′卵泡cDNA為模板,Long和5′ GSP分別為上、下游引物,采用Touch Down PCR對lncRNA-ENSSSCT00000018610進行5′序列擴增。反應體系:5′RACE Ready cDNA 2.5 μL,10×UPM(Long) 5 μL,5′GSP 1 μL,PCR Grade H2O 15.5 μL,2×SeqAmp Buffer 25 μL,SeqAmp DNA Polymerse 1 μL。反應條件:95 ℃ 5 min;(95 ℃ 30 s,72 ℃ 3 min)5個循環;(95 ℃ 30 s,70 ℃ 30 s,72 ℃ 3 min)10個循環;(95 ℃ 30 s,62 ℃ 30 s,72 ℃ 3 min)20個循環。1.2%瓊脂糖凝膠電泳檢測,將出現的明顯條帶膠回收,并連接pUC 19 Linearized Vector轉化并進行菌液測序。
3′ RACE PCR反應:試驗以3′卵泡cDNA為模板,Long和3′GSP分別為上、下游引物,采用Touch Down PCR對lncRNA-ENSSSCT00000018610進行3′序列擴增。反應體系:3′ RACE Ready cDNA 2.5 μL,10×UPM (Long)5 μL,3′GSP 1 μL,PCR Grade H2O 15.5 μL,2×SeqAmp Buffer 25 μL,SeqAmp DNA Polymerse 1 μL。反應條件:95 ℃ 5 min;(95 ℃ 30 s,72 ℃ 3 min)5個循環;(95 ℃ 30 s,70 ℃ 30 s,72 ℃ 3 min)15個循環;(95 ℃ 30 s,62 ℃ 30 s,72 ℃ 3 min)25個循環。1.2%瓊脂糖凝膠電泳檢測,將出現的明顯條帶膠回收,并連接pUC 19 Linearized Vector轉化并進行菌液測序。
1.7 豬lncRNA-ENSSSCT00000018610組織表達譜分析
試驗以各組織與卵泡cDNA為模板,GAPDH為內參,對lncRNA-ENSSSCT00000018610進行實時熒光定量PCR檢測以分析其在豬不同組織中的表達情況。反應體系:SYBR?Green I PCR Mix 10 μL,上、下游引物各1 μL,cDNA模板2 μL,加ddH2O至20 μL。反應條件:95 ℃預變性5 min;95 ℃ 變性20 s,56 ℃退火30 s,72 ℃延伸30 s。
1.8 lncRNA-ENSSSCT00000018610靶基因預測
試驗通過cis(co-location)和trans(co-expression)兩種方式對lncRNA-ENSSSCT00000018610靶基因進行預測。搜索lncRNA上游和下游100 kb內的編碼基因作為順式作用;反式作用是指通過表達水平鑒定彼此,并選擇具有Pearson相關系數|r|>0.95的結果[20-21]。
1.9 數據處理
以2-△△ Ct法[22]計算lncRNA-ENSSSCT00000018610在各組織及梅山豬和杜洛克豬各級卵泡中的相對表達量,用SPSS17.0軟件中One-Way ANOVA方法分析比較其在兩豬種內不同級別卵泡及品種間同級別卵泡表達水平的差異。
2 結 果
2.1 lncRNA-ENSSSCT00000018610的保守性分析和編碼潛能預測
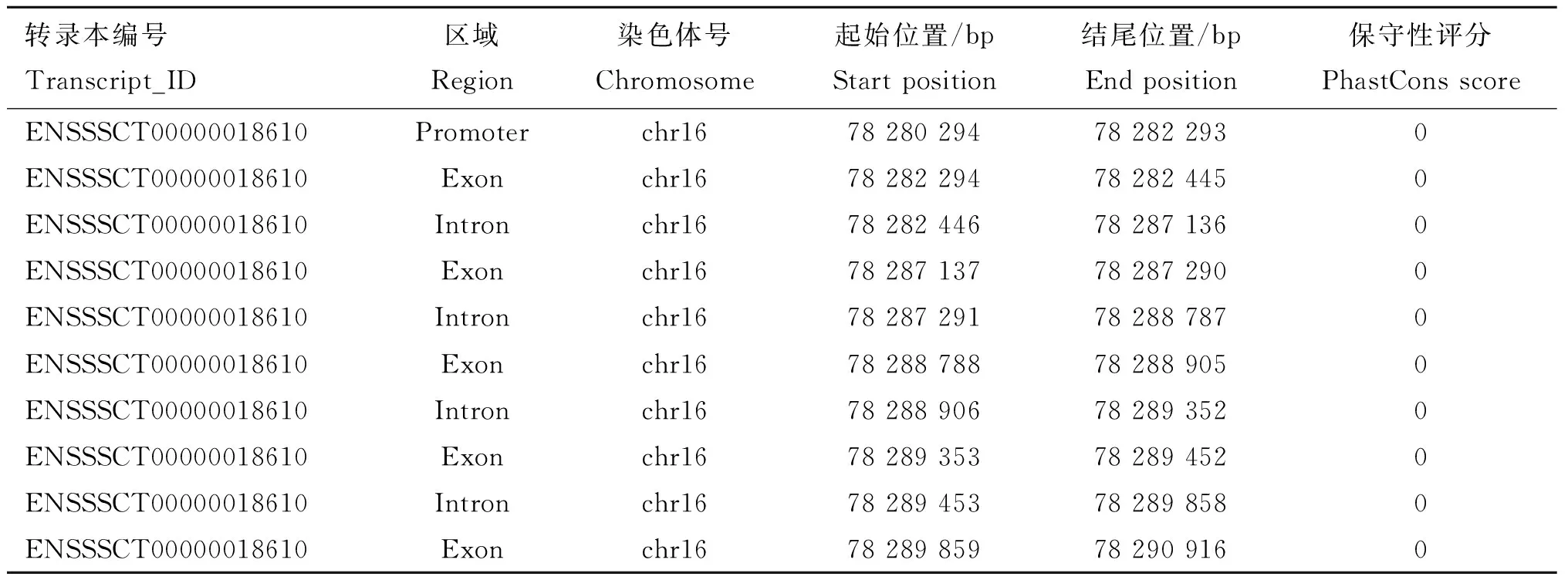
lncRNA-ENSSSCT00000018610位于豬基因組16號染色體,具有5個外顯子,但均不具有編碼潛能,對其保守性進行總體評估,保守性評分=0(表2),符合lncRNA具有不保守性特征。此外,利用Coding-Potential Assessment Tool(CAPT)法對lncRNA-ENSSSCT00000018610編碼能力進行預測,結果表明,lncRNA-ENSSSCT00000018610的編碼能力預測結果為0.159 346 863,該數據比已使用的人lncRNA編碼能力閾值0.364[23]要小,這進一步說明lncRNA-ENSSSCT00000018610不具有編碼蛋白的潛能。
表2lncRNA-ENSSSCT00000018610序列保守性評估
Table2ConservativeevaluationoflncRNA-ENSSSCT00000018610sequence
轉錄本編號Transcript_ID區域Region染色體號Chromosome起始位置/bpStart position結尾位置/bpEnd position保守性評分PhastCons scoreENSSSCT00000018610 Promoterchr1678 280 29478 282 2930ENSSSCT00000018610 Exonchr1678 282 29478 282 4450ENSSSCT00000018610 Intronchr1678 282 44678 287 1360ENSSSCT00000018610 Exonchr1678 287 13778 287 2900ENSSSCT00000018610 Intronchr1678 287 29178 288 7870ENSSSCT00000018610 Exonchr1678 288 78878 288 9050ENSSSCT00000018610 Intronchr1678 288 90678 289 3520ENSSSCT00000018610 Exonchr1678 289 35378 289 4520ENSSSCT00000018610 Intronchr1678 289 45378 289 8580ENSSSCT00000018610 Exonchr1678 289 85978 290 9160
2.2 豬lncRNA-ENSSSCT00000018610全長擴增
5′ RACE獲得一條長度約為1 000 bp的明亮條帶(圖1A),3′ RACE獲得一條長度超過1 000 bp的明亮條帶(圖1B),克隆測序分別獲得長度為1 027、1 343 bp的序列。
5′ RACE得到的1 027 bp序列中有940 bp與已知序列相同,獲得5′ 端未知序列74 bp。3′ RACE得到的1 343 bp左右序列中有1 269 bp與已知序列相同,且3′末端有明顯的Poly A尾巴,獲得3′端75 bp的未知序列。通過5′和3′ RACE擴增結果拼接,總共獲得lncRNA-ENSSSCT00000018610的全長序列1 731 bp(圖2)。
2.3 lncRNA-ENSSSCT00000018610在豬不同組織中的表達分析
試驗采用RT-PCR技術對lncRNA-ENSSSCT00000018610在豬不同組織中的表達情況作了分析,結果表明,其在下丘腦、垂體、心、肝、脾、肺、腎、小腸、肌肉、子宮、輸卵管、黃體、卵巢中均有不同程度的表達(圖3)。

M. DNA相對分子質量標準;5′. 5′ RACE PCR電泳條帶; 3′. 3′ RACE PCR電泳條帶M. Marker 10000; 5′. 5′ RACE PCR electrophoresis band; 3′. 3′ RACE PCR electrophoresis band圖1 lncRNA-ENSSSCT00000018610 5′-RACE(A)和3′-RACE(B)擴增結果電泳產物圖Fig.1 The electrophoresis of PCR products of 5′-RACE(A)and 3′-RACE (B) of lncRNA-ENSSSCT00000018610

圖中第一行藍色大寫字母序列為lncRNA-ENSSSCT00000018610的高通量測序結果文件序列,第二行兩端綠色大寫字母序列分別為5′ 和3′ RACE末端克隆獲得序列,中間為5′與3′ RACE拼接序列。第三行小寫字母序列為RACE克隆得到的lncRNA-ENSSSCT00000018610 cDNA序列The first row of blue capital letters in the figure is the lncRNA-ENSSSCT00000018610 high-throughput sequencing result file sequence. The second row of green capital letters are the sequence of the 5′ and 3′ RACE clones, and the middle is 5′ and 3′ RACE splicing sequence. The third row of lowercase letters sequence is the lncRNA-ENSSSCT00000018610 cDNA sequence cloned from RACE圖2 5′與3′ RACE測序序列比對與拼接結果Fig.2 The result of 5′ and 3′ RACE sequence comparison and splicing
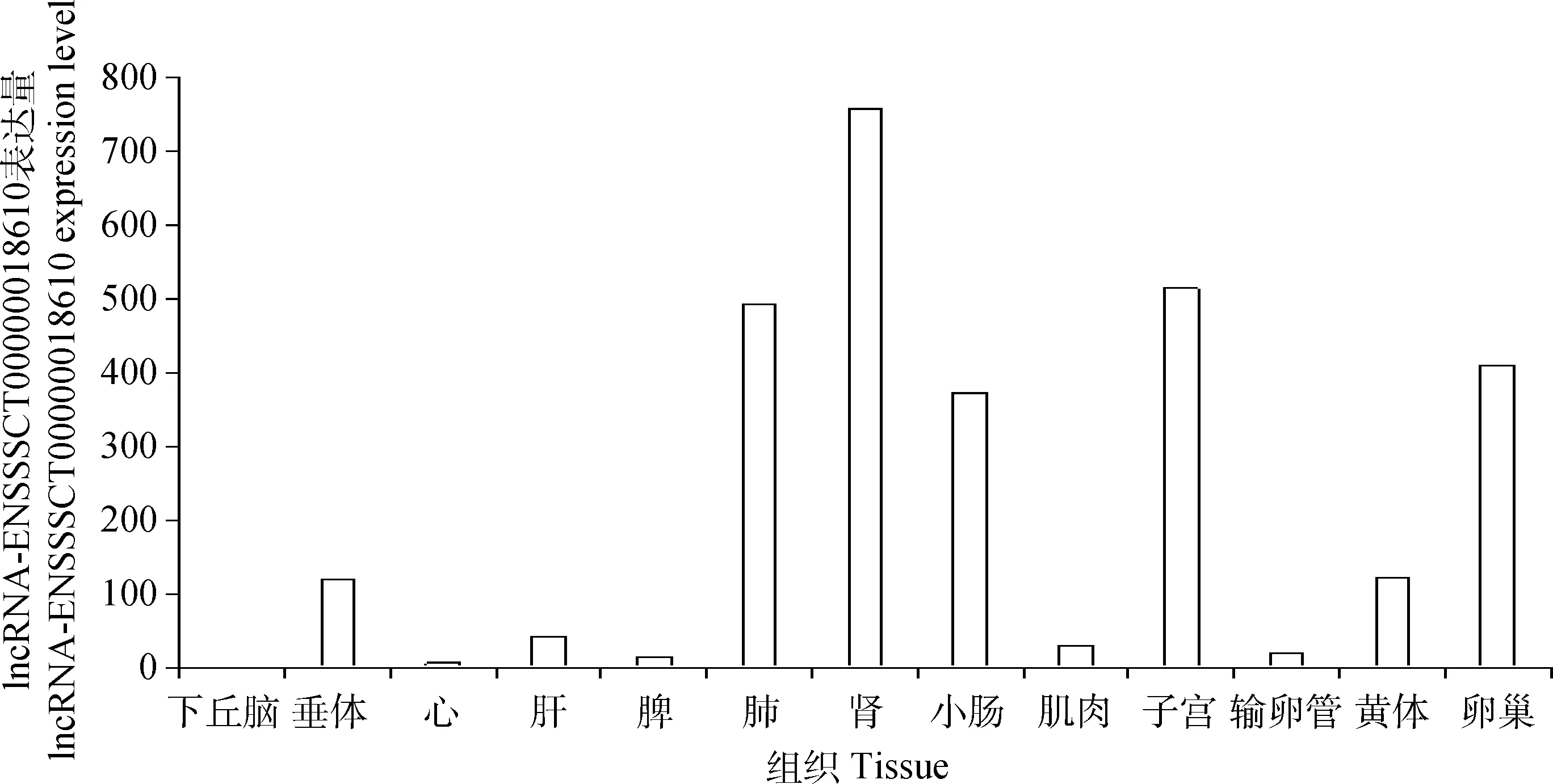
圖3 lncRNA-ENSSSCT00000018610在豬不同組織中的表達情況Fig.3 Tissues expression profile of lncRNA-ENSSSCT00000018610 in pigs
2.4 lncRNA-ENSSSCT00000018610在豬卵泡中的表達分析
試驗采用RT-PCR技術檢測了梅山與杜洛克豬不同級別卵泡中lncRNA-ENSSSCT0000-0018610的表達情況(圖4)。結果表明,lncRNA-ENSSSCT00000018610在梅山豬M2卵泡中的表達量高于S、M1、L卵泡(P<0.05),在杜洛克豬M1、M2卵泡中的表達量極顯著高于S和L卵泡(P<0.01),提示該lncRNA可能與卵泡發育有關。在杜洛克豬S、L卵泡中的表達量分別顯著高于在梅山豬S、L卵泡中的表達量(P<0.05),在杜洛克豬的M1、M2卵泡中的表達量分別極顯著高于在梅山豬的M1、M2卵泡中的表達量(P<0.01)。
2.5 豬lncRNA-ENSSSCT00000018610的靶基因預測
本研究通過構建蛋白編碼基因的位置關系(co-location),以及與蛋白編碼基因表達相關性(co-expression)預測得到lncRNA-ENSSSCT00000018610有多個潛在靶基因(表3)。
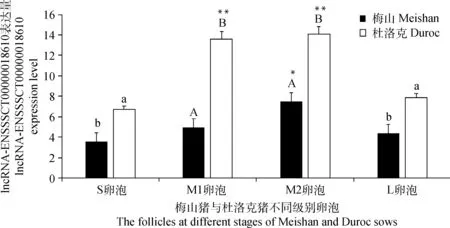
同一品種不同級別卵泡間,**表示差異極顯著(P<0.01);*表示差異顯著(P<0.05)。不同品種同級卵泡間,肩標大寫字母不同表示差異極顯著(P<0.01);肩標小寫字母不同表示差異顯著(P<0.05)Among the different follicles in the same breed, ** mean extremely significant difference(P<0.01);* mean significant difference(P<0.05). In the same follicles of different breeds, the different capital letters mean extremely significant difference(P<0.01); the different small letters mean significant difference(P<0.05)圖4 lncRNA-ENSSSCT00000018610在梅山和杜洛克母豬各級卵泡中的相對表達量Fig.4 The relative expression of lncRNA-ENSSSCT00000018610 in follicles at different stages of Meishan and Duroc sows
表3lncRNA-ENSSSCT00000018610靶基因預測
Table3lncRNA-ENSSSCT00000018610targetgenesprediction
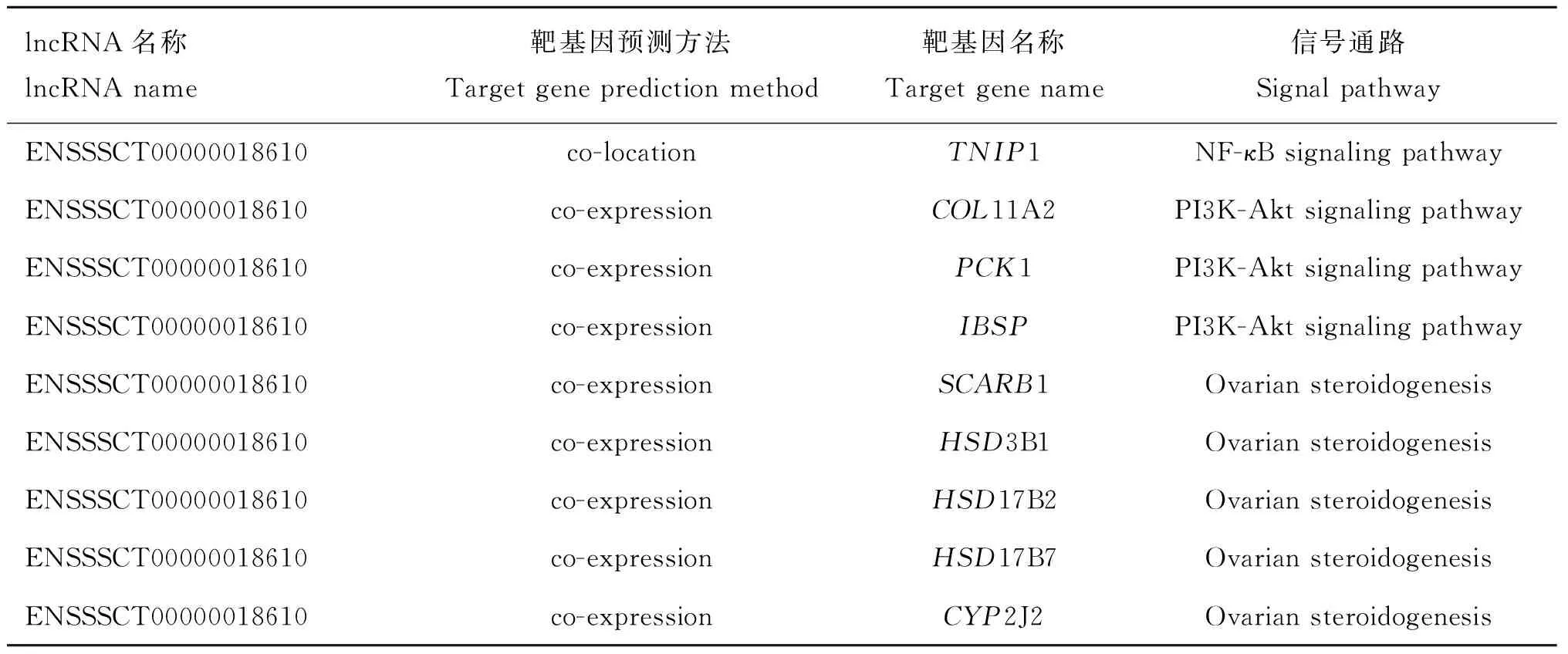
lncRNA名稱lncRNA name靶基因預測方法Target gene prediction method靶基因名稱Target gene name信號通路Signal pathwayENSSSCT00000018610co-locationTNIP1NF-κB signaling pathwayENSSSCT00000018610co-expressionCOL11A2PI3K-Akt signaling pathwayENSSSCT00000018610co-expressionPCK1PI3K-Akt signaling pathwayENSSSCT00000018610co-expressionIBSPPI3K-Akt signaling pathwayENSSSCT00000018610co-expressionSCARB1Ovarian steroidogenesisENSSSCT00000018610co-expressionHSD3B1Ovarian steroidogenesisENSSSCT00000018610co-expressionHSD17B2Ovarian steroidogenesisENSSSCT00000018610co-expressionHSD17B7Ovarian steroidogenesisENSSSCT00000018610co-expressionCYP2J2Ovarian steroidogenesis
TNIP1是通過lncRNA-ENSSSCT00000018610位置關系(co-location)預測得到的潛在靶基因,KEGG富集分析發現,TNIP1被富集在NF-κB信號通路上,GO分析發現其參與19種生物學過程;CYP2J2、SCARB1、IBSP等基因是通過lncRNA-ENSSSCT00000018610表達相關性(co-expression)預測得到的潛在靶基因,它們分別被富集在卵巢類固醇激素通路和PI3K-Akt信號通路,參與豬卵巢類固醇激素合成、細胞生長分化以及卵泡發育等生物學過程。這提示,lncRNA-ENSSSCT00000018610可能通過對其靶基因的調控間接參與豬卵泡發育。
3 討 論
lncRNA在各類生命活動中發揮著重要的調節作用,受到了更多科學家的關注。目前lncRNA的研究多停留在芯片檢測和高通量測序層面上,對其機理的探究更是局限于人類的疾病、免疫及模式生物上,而在重要的經濟動物生長發育及繁殖上,lncRNA的研究相對滯后,仍然處于摸索階段。
隨著基因芯片和測序技術的不斷發展,對lncRNA的獲取和鑒定方法也在不斷更新,不同的研究者提出了不同的研究方法和鑒定流程,主要是高通量測序技術和基于lncRNA特征的計算機預測鑒定。高通量測序技術被廣泛應用于非編碼RNA研究,lncRNA測序能夠通過rRNA去除法對含有和不含有PolyA尾巴的RNA一同富集建庫測序,使得數據不浪費,從而能夠鑒定到更多的lncRNA[23-24],這也使得lncRNA測序在更多的研究中應用。基于lncRNA特征的計算機預測包括CPAT、CPC、PhyloCSF等軟件[25]。CPAT能從大量的候選轉錄物中迅速確認出編碼和非編碼記錄,它用了一種全新的序列匹配的方法。CPAT預測方法采用邏輯回歸模型,使用4種序列特征:ORF長度、覆蓋率特征、Fickett Score和六聚體偏好性特征[11]。就軟件的預測性能而言,CPAT (敏感性 0.96,異性:0.97)精度更優于CPC (敏感性 0.99,異性:0.74)、PhyloCSF (敏感性 0.90,異性:0.63)[26]等方法,此外CPAT運算速度比其他軟件快大約4個數量級[27]。因此這個運算速度快、方便且精度高的編碼能力預測軟件被研究者所認可。本試驗所用的lncRNA是通過高通量測序技術結合計算機軟件分析獲得的,高通量測序結果顯示,lncRNA-ENSSSCT00000018610在梅山和杜洛克豬中等卵泡中的表達差異極顯著,加之使用CPAT對其進行編碼能力預測,結果顯示其編碼能力分值為0.159,比已使用的人長鏈非編碼RNA篩選閾值0.364[28]小。lncRNA-ENSSSCT00000018610含有一個長度為360 nt的ORF,編碼120個氨基酸,而許多研究比較深入的lncRNA,如HOTAIR家族和H19都存在長度大于100 aa的ORF[29],這些ORF極有可能不翻譯或翻譯效率很低,也可能翻譯出來的蛋白質會被快速的降解。所以盡管lncRNA-ENSSSCT00000018610的ORF >100 aa,其也不具有編碼能力或者編碼能力很弱。當然,也有一些RNA在最初的研究中被鑒定為lncRNA,但隨著研究的不斷深入,發現其也有編碼能力[30],這些都需要更進一步驗證。
RACE(rapid amplification of cDNA ends)是一種快速克隆cDNA末端的技術,此方法被廣泛應用于未知堿基序列基因的克隆。該方法基于PCR技術,步驟少、耗時短,只需知道RNA內部很短的一小段cDNA序列就能快速克隆出基因的未知序列。RACE技術包括3′和5′末端序列擴增,完整且純度高的總RNA和高質量3′和5′RACE ready cDNA是RACE成功的基礎,而根據已知核酸序列設計出的優良特異性引物是RACE成功的決定性因素。本試驗通過RACE技術成功克隆出了lncRNA-ENSSSCT00000018610全長為1 731 bp的cDNA。這為進一步研究lncRNA-ENSSSCT00000018610的生物學功能奠定了基礎。
本研究通過在杜洛克豬不同組織中對lncRNA-ENSSSCT00000018610進行的熒光定量PCR發現,這個lncRNA在杜洛克豬各組織中廣泛表達且范圍波動較大,說明它具有組織表達特異性,其在子宮和卵巢中的較高表達可能提示,lncRNA-ENSSSCT00000018610可能參與母豬生殖活動過程。在梅山豬和杜洛克豬不同級別卵泡中的定量PCR顯示,lncRNA-ENSSSCT00000018610在梅山豬中等卵泡M1、M2中的表達量均極顯著低于杜洛克豬的M1和M2卵泡,與高通量測序結果一致,這說明了高通量測序具有一定的準確性。
此外,本研究通過構建lncRNA與蛋白編碼基因的位置關系(co-location)預測得到其潛在靶基因TNIP1(又名ABIN-1),可通過結合A20調節對NF-κB信號的抑制作用,與細胞的生長分化以及動物胚胎發育相關[31],并且是一個影響長白豬產仔性狀的候選基因[32];lncRNA與蛋白編碼基因表達相關性(co-expression)預測得到其潛在靶基因CYP2J2、SCARB1、HSD3B1、HSD17B2、HSD17B7參與卵巢類固醇激素合成信號傳導,參與卵泡發育及排卵過程,COL11A2、FGFR1、IBSP參與PI3K-Akt信號通路,而PI3K-Akt通路在哺乳動物卵泡發生和排卵的過程中起著關鍵的調控作用[33-34]。因此,lncRNA-ENSSSCT00000018610可能通過對其靶基因的調控作用間接參與卵泡發育過程,但lncRNA-ENSSSCT00000018610與預測得到的蛋白質編碼基因之間的靶向關系仍需進一步驗證。
4 結 論
lncRNA-ENSSSCT00000018610是一個不具編碼潛能的非編碼RNA,RACE克隆獲得其全長序列1 731 bp。lncRNA-ENSSSCT00000018610在豬腎、子宮、卵巢中表達水平相對較高。lncRNA-ENSSSCT00000018610在梅山豬M2卵泡中的表達量顯著高于S、M1、L卵泡,在杜洛克豬M1、M2卵泡中的表達量極顯著高于S和L卵泡,在杜洛克豬S、L卵泡中的表達量分別顯著高于梅山豬,在杜洛克豬M1、M2卵泡中的表達量分別極顯著高于梅山豬。靶基因預測結果說明,lncRNA-ENSSSCT00000018610可能通過對其靶基因的調控作用間接參與卵泡發育過程。該試驗對lncRNA-ENSSSCT00000018610的研究為進一步揭示其在豬卵泡發育中的作用機制提供了重要參考依據。
猜你喜歡
黃河之聲(2019年1期)2019-03-30 03:38:06
少兒美術(快樂歷史地理)(2018年4期)2018-09-25 02:48:54
黃河之聲(2018年6期)2018-05-18 00:47:57
作文新天地(2016年18期)2016-02-13 01:15:01
中國資源綜合利用(2016年5期)2016-02-03 02:56:17
邵陽學院學報(社會科學版)(2015年3期)2015-02-28 16:18:23
水道港口(2014年1期)2014-04-27 14:14:35
電力需求側管理(2014年3期)2014-03-20 13:35:43
水道港口(2013年3期)2013-04-27 14:14:30
湖南人文科技學院學報(2012年6期)2012-04-16 03:32:32
