浙江金華螢石礦集區(qū)地質(zhì)特征及成礦條件分析
2021-07-06 13:04:52黃光輝
甘肅科技 2021年9期
關(guān)鍵詞:成礦
黃光輝
(東華理工大學(xué)地球科學(xué)學(xué)院,江西 南昌 330013)
1 概述
該區(qū)位于浙江省中部,地處浙中丘陵盆地區(qū),以中低山、低山、丘陵盆地地貌為主,地勢南北高、中部低,為江紹斷裂帶中段和浙江中生代火山巖匯合區(qū),以斷裂構(gòu)造、火山構(gòu)造、盆地構(gòu)造最為顯著,褶皺不發(fā)育。經(jīng)過野外地質(zhì)調(diào)查和資料對比,對這一地區(qū)的螢石礦的成礦地質(zhì)條件進行系統(tǒng)的分析研究,總結(jié)出螢石的找礦標(biāo)示,為該區(qū)螢石的勘探工作提供參考資料。
2 區(qū)域地質(zhì)概況
該區(qū)坐落于江紹斷裂帶中部和浙東中生代火山巖集中分布區(qū)。大地構(gòu)造位置顯示為兩大構(gòu)造單元,西北區(qū)屬下?lián)P子陸塊東南緣,主體處在常山-諸暨臺隆之航埠-漓渚臺拗內(nèi),東南區(qū)屬華夏陸塊北緣邊部,居于麗水-寧波隆起帶之新昌-鎮(zhèn)海隆斷束與龍泉-遂昌隆斷束內(nèi)[1-3]。西北以新元古代淺變質(zhì)巖系構(gòu)成雙層基底,蓋層為南華-奧陶紀(jì)陸緣海相沉積巖系和中生代陸相火山沉積巖系;東南以元古代八都巖群、陳蔡巖群變質(zhì)雜巖系為結(jié)晶基底,蓋層為中新生代陸相火山沉巖系。區(qū)內(nèi)深大斷裂及燕山期構(gòu)造盆地發(fā)育,燕山晚期巖漿-火山活動頻發(fā),大范圍的中酸性火山噴發(fā)、巖漿侵入和雙峰式火山噴發(fā)、潛火山巖的形成[4],火山期后熱液作用產(chǎn)生眾多的螢石礦床(點)和其他一系列內(nèi)生礦產(chǎn)。
3 區(qū)域成礦特征
3.1 螢石礦床的分布及控制特征
區(qū)內(nèi)螢石礦嚴(yán)格受斷裂構(gòu)造控制,包括北東向、北北東向、東西向及北西向斷裂。
并與火山構(gòu)造、(火山洼地型)盆地構(gòu)造及潛火山巖關(guān)系密切。其中大、中型螢石礦床多產(chǎn)于北東向(或北北東向)斷裂中,如武義楊家、后樹、雞舍灣、潘村-鄭山頭、徐村、溪里、余山頭、永康花溪、瀆溪、西寮、東陽佐村、忠信堂及磐安塘塢、蘭溪冷坑山、浦江溪前等螢石礦;東西向斷裂亦控制了部分中型螢石礦床的分布,如金華楊梅峽、武義麻栗畈、永康下塹、橫山、楓林、東陽上溪和磐安張斯等螢石礦;北西向斷裂控制的螢石礦一般為小型規(guī)模或礦(化)點,但品位較富(如圖1 所示)。
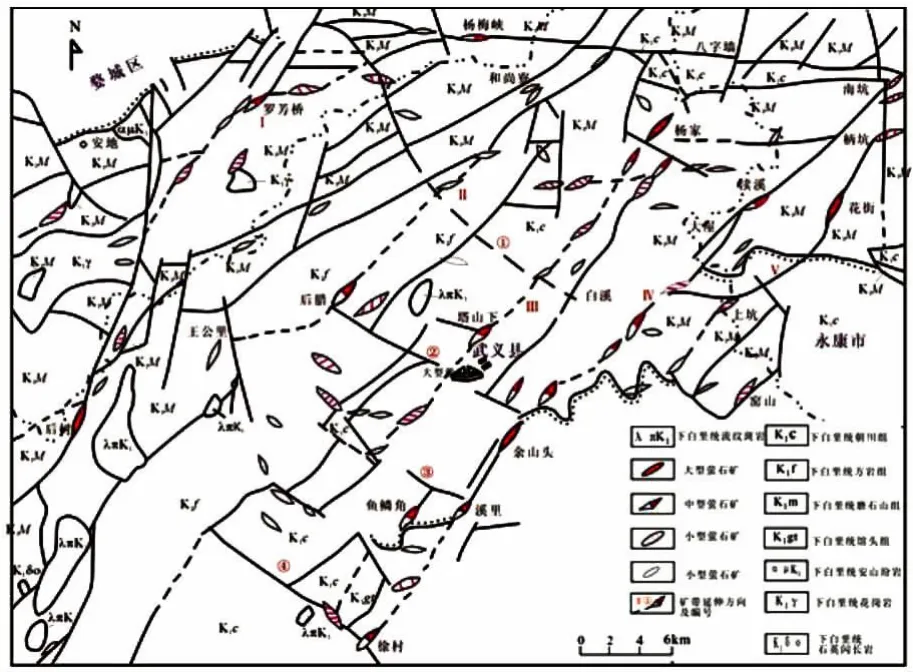
圖1 武義北部礦田地質(zhì)略圖(據(jù)《武義螢石志》(1994),略有修編)
3.2 螢石礦床的地質(zhì)特征
螢石礦床賦存于斷裂帶及裂隙帶中,礦床成因以淺成熱液充填型為主,斷裂控礦特征明顯,礦體呈脈狀,少數(shù)為透鏡狀、似層狀,分支復(fù)合、尖滅再現(xiàn)、尖滅側(cè)現(xiàn)和膨縮現(xiàn)象常見,長幾十米到一千米不等,延伸情況多不詳(一般僅控制幾十米至三百米不等)。礦石以塊狀構(gòu)造、角礫狀構(gòu)造(正角礫、負(fù)角礫)為主,其次條帶狀構(gòu)造等,常為半自形-它形粒狀結(jié)構(gòu)。礦石類型以螢石型、石英-螢石型、螢石-石英型為主,品位40%到95%以上不等。圍巖蝕變以硅化、高嶺土化為主,次為絹云母化、綠泥石化,少見方解石化、黃鐵礦化、冰長石化,局部伴生鉬礦化、金銀礦化、鎢礦化等。
3.3 成礦控制條件
1)區(qū)域地質(zhì)條件。工作區(qū)地處下?lián)P子陸塊與華夏陸塊對接部位,主體處于浙東中生代火山巖集中分布區(qū)。區(qū)內(nèi)深大斷裂構(gòu)造發(fā)育,巖漿活動頻繁,早白堊世早期太平洋板塊向歐亞板塊俯沖,處于北西-南東向強烈擠壓,形成泛陸式火山噴發(fā)堆積及巖漿侵入活動;早白堊世晚期區(qū)域應(yīng)力場轉(zhuǎn)入了引張體制,主壓應(yīng)力軸方向變?yōu)楸蔽魑?南東東向,出現(xiàn)了麗水-余姚等北北東向深斷裂及北東東向、北西西向等次級斷裂及東西向斷裂復(fù)活,形成了一系列的斷陷構(gòu)造盆地及斷隆斷塊,并控制了東陽-遂昌高溫暖流帶的形成和分布。沿早期斷裂局部產(chǎn)生繼承性張性斷裂,成為礦液充填的容礦空間。
2)地層巖石條件。區(qū)內(nèi)礦床(點)的主要賦礦地層是下白堊統(tǒng)磨石山群、永康群,含氟量較高,賦礦的酸性火山碎屑巖(凝灰?guī)r、熔結(jié)凝灰?guī)r)含氟量在0.10%左右(有的高達0.46%)。陳蔡群平均含氟量為0.091%(氟背景值的1.5 倍)。以武義盆地為例,礦床圍巖,西山頭組占礦床(點)總數(shù)的41.5%、大爽組占12.5%、朝川組占23.5%。西山頭組是武義、永康、南馬等盆地周邊最重要的賦礦地層,區(qū)內(nèi)大型礦床、62.5%的中型礦床和45.24%的小型礦床均賦存在該地層中[5-6]。基底和火山蓋層的雙重較高氟背景對浙東南,特別是武義-永康-東陽一帶中生代火山巖中脈狀螢石礦的形成最為有利,可能是氟的最主要來源。
主要原因是:西山頭組地層中熔結(jié)凝灰?guī)r、凝灰?guī)r、含(角)礫凝灰?guī)r、流紋巖等脆性巖石所占厚度大,比例高;裂隙發(fā)育,孔隙度大,滲透性好,礦液易于循環(huán);主要分布于盆地邊緣,處于有利構(gòu)造部位。
3)構(gòu)造條件。本區(qū)螢石礦床主要受盆邊斷裂及其套配套的次級斷裂控制,特別是北東向或北北東向斷裂與北西向斷裂交匯處為良好的容礦空間,盆內(nèi)及盆緣斷裂多期次活動,造成多期次含礦熱液運移。礦體的形狀、產(chǎn)狀及規(guī)模嚴(yán)格受斷裂裂隙控制。
4)“紅層”盆地條件。區(qū)內(nèi)武義、永康、南馬、墩頭等火山構(gòu)造盆地發(fā)育了較厚的中生界陸相沉積巖和火山巖,負(fù)向構(gòu)造盆地可造成盆地中的承壓水動力系統(tǒng),火山活動、巖漿期后熱液及大氣降水不僅是成礦流體-含礦地?zé)崴闹饕a給來源,而且滲流的大氣降水與含礦地?zé)崴幕旌鲜窃摰貐^(qū)螢石礦形成的重要條件之一。
3.4 找礦標(biāo)志
1)區(qū)域構(gòu)造標(biāo)志。燕山晚期,中生代陸相盆地進入衰亡階段落,在區(qū)域北西西-南東東擠壓應(yīng)力作用下,全區(qū),特別在武義-永康一帶,形成平行展布的北東向斷裂及其配套的北西向斷裂形成網(wǎng)格狀構(gòu)造格局,螢石礦床(點)呈等間距分布特點。螢石礦帶與早期斷裂重合處是大、中型礦床的主要賦存部位;次級火山構(gòu)造、潛火山巖與區(qū)域性斷裂的復(fù)合部位、多組斷裂(或礦帶)交匯處是成礦有利部位。
2)多階段成礦作用標(biāo)志。多階段成礦導(dǎo)致礦體富集或增厚,有利形成大、中型礦床,尤其是塊狀、負(fù)角礫狀礦石形成階段是形成有工業(yè)價值螢石礦床的重要階段[8-9]。多階段成礦特點反映在礦石特征上,是各種構(gòu)造類型礦石的出現(xiàn)和不同結(jié)構(gòu)構(gòu)造的螢石脈、石英螢石脈的穿插、膠結(jié),區(qū)內(nèi)大中型螢石礦床通常具有復(fù)雜多樣的礦石構(gòu)造。
3)圍巖蝕變標(biāo)志。螢石礦床近礦蝕變?yōu)榈蜏匚g變組合,蝕變?nèi)酰謳Р幻黠@,硅化帶不發(fā)育,硅化的強弱與礦體富集無必然聯(lián)系,但螢石礦化與硅化關(guān)系密切,可作為找礦的直接證據(jù)。
4)地球化學(xué)標(biāo)志。區(qū)內(nèi)大中型螢石礦床在水系沉積物測量F、CaO 地球化學(xué)圖上均顯示極為明顯的高含量區(qū),且F、CaO 異常套合程度高,異常強度大。因此,F(xiàn)、CaO 異常是很好的找礦標(biāo)志。對一些低值異常,若民采規(guī)模小,則也不應(yīng)忽視。
5)其他標(biāo)志。地表螢石脈、石英螢石脈露頭、轉(zhuǎn)石、機械分散暈、采礦遺址是直接的找礦標(biāo)志。
4 結(jié)論
1)區(qū)內(nèi)螢石礦嚴(yán)格受斷裂構(gòu)造控制,包括北東向、北北東向、東西向及北西向斷裂。并與火山構(gòu)造、(火山洼地型)盆地構(gòu)造及潛火山巖關(guān)系密切,螢石礦床賦存于斷裂帶及裂隙帶中,礦床成因以淺成熱液充填型為主,
2)通過分析區(qū)內(nèi)的成礦控制條件,大致厘清找礦的重點標(biāo)志:圍巖蝕變標(biāo)志、地球化學(xué)標(biāo)志、地表螢石脈、石英螢石脈露頭、轉(zhuǎn)石、機械分散暈、采礦遺址等直接標(biāo)志。
猜你喜歡
大地構(gòu)造與成礦學(xué)(2021年5期)2021-10-27 11:15:36
大地構(gòu)造與成礦學(xué)(2021年4期)2021-08-24 05:34:48
河北地質(zhì)(2021年1期)2021-07-21 08:16:04
礦產(chǎn)勘查(2021年3期)2021-07-20 08:01:52
大地構(gòu)造與成礦學(xué)(2021年3期)2021-06-29 11:16:24
大地構(gòu)造與成礦學(xué)(2021年2期)2021-05-07 13:57:12
礦產(chǎn)勘查(2020年2期)2020-12-28 00:23:50
礦產(chǎn)勘查(2020年1期)2020-12-28 00:21:12
礦產(chǎn)勘查(2020年6期)2020-12-25 02:42:08
礦產(chǎn)勘查(2020年6期)2020-12-25 02:41:08
