干旱脅迫下谷子的轉錄組分析
2018-11-05 02:48:26王慶國李臻管延安劉煒潘教文
山東農業科學 2018年10期
關鍵詞:途徑
王慶國,李臻,管延安,劉煒,潘教文
(1.山東省農業科學院生物技術研究中心/山東省作物遺傳改良與生理生態重點實驗室,山東 濟南 250100;2.山東省農業科學院作物研究所,山東 濟南 250100)
谷子[Setariaitalica(L.)Beauv.]又稱為粟,是起源于我國的傳統優勢作物,也是糧飼兼用作物。谷子去皮后俗稱小米,小米營養豐富且各種成分均衡,具有很高的營養保健價值,被我國列為“五谷”之首。谷子具有抗旱、耐瘠薄、生育期短、C4高光效、適應性廣等特點,且細胞壁較厚、葉面積小、根系發達、水分利用效率較高,對干旱和鹽堿脅迫等都具有較好的耐受性。隨著谷子基因組測序計劃的完成及基因組序列的釋放,谷子逐漸發展成為研究禾本科作物新的模式作物[1,2]。
近年來結合高通量測序研究谷子的抗旱機制,并鑒定干旱響應基因及非編碼RNA已有報道。Qi等[3]研究發現,PEG處理谷子后可導致2 824個基因及215個 miRNA的表達發生變化。Yadav等[4]對兩個抗性不同的谷子品種測序,共鑒定到55個已知miRNA和136個新miRNA,其中18個已知和33個新miRNA均參與干旱脅迫響應。Wang等[5]在干旱敏感品種An04中鑒定到81個已知的miRNA,其中有14個表達受干旱誘導,4個表達下調,同時也鑒定到72個新的miRNA,并結合降解組鑒定到82個miRNA的靶基因。最近,Tang等[6]對兩個抗性不同的谷子品種進行了生理和轉錄組分析,發現干旱脅迫后抗性品種 “豫谷1號”通過抑制能量代謝來保護細胞,從而維持正常的植株生長;而干旱敏感品種An04受脅迫后會消耗更多的能量,光合效率和細胞生長受到嚴重抑制。結合抗旱相關的QTL,作者進一步鑒定到萌發期和苗期干旱抗性相關的20個候選基因。定量蛋白質組學分析發現,谷子在干旱脅迫后會產生大量的參與滲透保護和抗氧化的蛋白,參與能量代謝的蛋白也有較明顯的積累[7]。
隨著谷子基因組序列的釋放,谷子中脅迫相關的基因家族也逐漸被鑒定出來。如NAC、AP2/ERF、C2H2型鋅指轉錄因子、MYB、WRKY等轉錄因子家族,及RNA聚合酶基因家族、PPR蛋白基因家族、DCPK基因家族、HSP基因家族等。谷子中各種脅迫及發育相關基因的功能也逐漸被研究及報道[8-14]。在水稻中異源表達自噬相關基因(autophagy-associated genes, ATG)SiATG8a能有效提高水稻對氮饑餓的耐受性[15]。在擬南芥中過表達SiCDPK24可顯著增強對干旱脅迫的抗性。SiNAC1通過促進ABA的合成加速了葉片的衰老[13]。在K+缺乏和鹽脅迫下,SiHAK1作為K+吸收及轉運蛋白維持胞內K+的穩態。過表達LEA蛋白SiLEA14顯著提高了轉基因擬南芥和谷子的非生物脅迫抗性[16]。SiAGO1(argonaute1)對谷子正常的生長發育和脅迫響應是必需的,該基因缺失導致多重發育缺陷,如植株矮小、葉片狹窄卷曲、小穗以及結實率低[17]。SiLP1編碼一個WRKY轉錄因子,該基因參與調控穗的發育、莖的伸長及種子的大小[11]。
鑒于谷子抗旱機制的研究還不夠深入,本研究利用RNA-seq 技術對干旱脅迫處理不同時間的谷子進行了測序分析,篩選干旱脅迫響應基因,系統分析谷子的抗旱響應機制,為進一步克隆抗旱相關基因提供了重要依據。
1 材料與方法
1.1 谷子種植與干旱處理
精選“豫谷1號”種子播種于花盆中,每個花盆中裝有3 kg營養土和沙子的混合物(比例為1∶1),在溫室中正常生長2周左右,光周期為14 h,晝夜溫度為30℃/25℃。用20%的PEG模擬干旱進行短期處理,將花盆中的谷子幼苗取出,用清水將根部土壤洗凈,置于Hoagland營養液中適應幾個小時,然后用Hoagland營養液配置20%的PEG溶液處理6 h。長期處理直接在花盆中處理,當花盆中的土壤含水量低于30%后持續處理一周的時間,使含水量控制在20%~30%。處理完的材料經液氮迅速冷凍,保存于-80℃超低溫冰箱備用。
1.2 利用RNA-seq技術篩選參與谷子干旱響應的基因
與北京諾禾致源科技股份有限公司進行合作,進行了RNA-seq相關試驗及數據分析,結合GO注釋(包括細胞組成、分子功能、生物過程)及KEGG代謝通路注釋,對篩選出的差異基因進行分析及歸類。
2 結果與分析
2.1 不同時間處理后差異基因分析
在 RNA-seq 技術中,FPKM (expected number of Fragments Per Kilobase of transcript sequence per Millions base pairs sequenced) 是每百萬測序堿基中來自某一基因每千堿基長度的fragments數目,其同時考慮了測序深度和基因長度對fragments計數的影響,是目前最為常用的基因表達水平估算方法。利用log2FoldChange: log2(樣品1的FPKM值/樣品2的FPKM值)代表差異基因的變化倍數。以log2FoldChange值大于2定義為表達上調的基因,以該值小于-2為表達下調的基因。如圖1A所示,與對照相比,谷子處理6 h后上調的基因有430個,下調的基因有183個。處理6 d后,與對照相比,上調的基因有606個,下調的基因有1 038個(圖1B)。谷子處理6 d后與處理6 h相比,上調的基因有206個,下調的基因有611個(圖1C)。
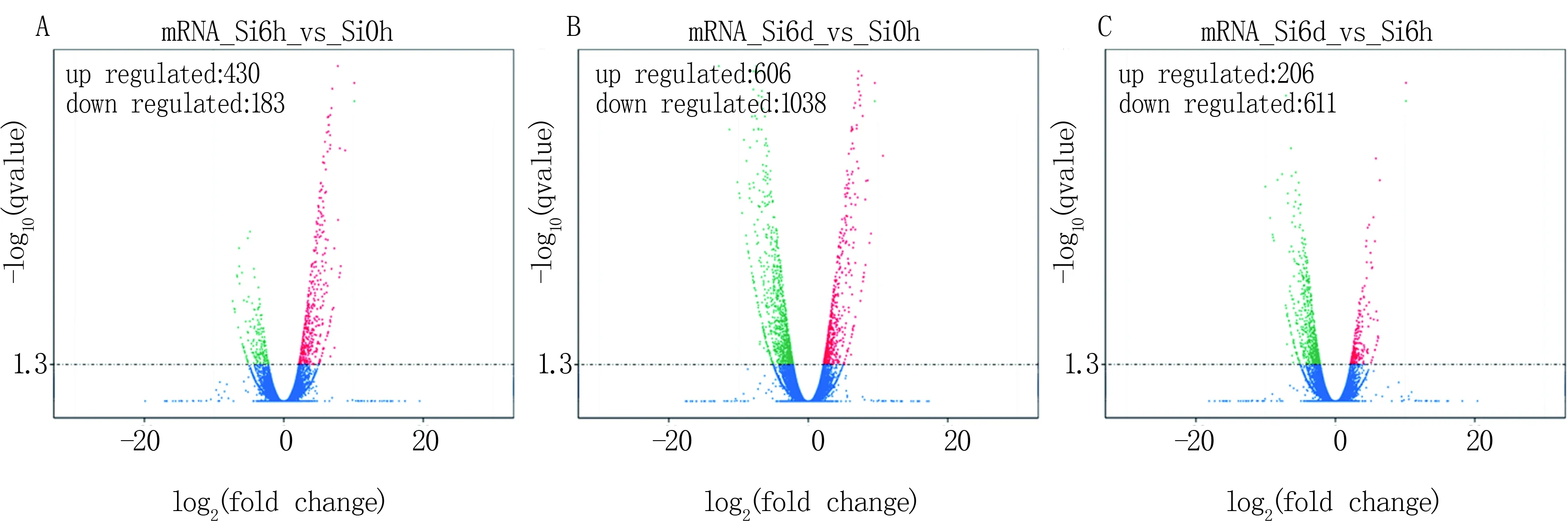
A:谷子處理6 h后的差異基因分布;B:谷子處理6 d后的差異基因分布;C:谷子處理6 d后與處理6 h相比的差異基因分布;篩選閾值默認設置為Q-value < 0.05。
對這三組差異的轉錄本進行共有與特有的差異轉錄本分析發現,有45個基因在這三組數據中共同存在,有177個差異基因在處理6 h后的谷子中被特異調控,有834個差異基因特異存在于處理6 d后的谷子中,有222個差異基因特異存在于處理6 d和6 h谷子的差異基因中(圖2)。
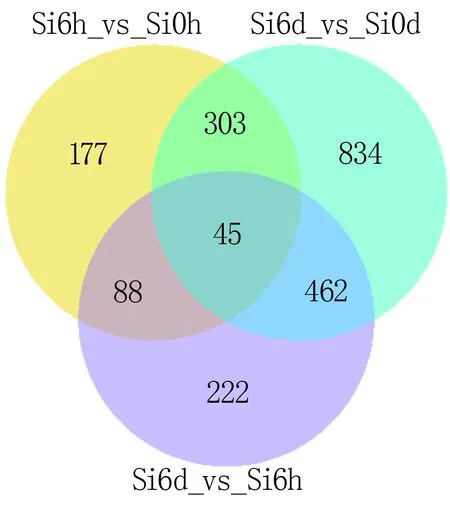
圖2 各差異轉錄本比較組合之間共有與特有數目的維恩圖
2.2 差異基因GO (Gene Ontology)富集分析
根據篩選到的差異基因,研究特異基因在 Gene Ontology 中的分布狀況將有助于闡明試驗中樣本差異在基因功能上的體現。在GO富集分析中,主要分為三大類biological process (BP)、cellular component (CC)和molecular function (MF)(圖3)。
谷子處理6 h后的差異基因聚類分析發現,在biological process中,差異基因主要富集在biological_process和metabolic process中;在cellular component (CC)中,差異基因主要富集在extracellular region中;在molecular function (MF)中,差異基因主要富集在catalytic activity、metal ion binding和cation binding(圖3A)。在干旱處理6 d后的差異基因中,在biological process中,差異基因主要富集在biological_process和metabolic process中;在cellular component (CC)中,差異基因主要富集在membrane、intrinsic to menbrane和integral to membrane;在molecular function (MF)中,差異基因主要富集在catalytic activity和oxidoreductase activity中(圖3B)。在谷子處理6 d和6 h后的差異基因中,在biological process中,差異基因主要富集在metabolic process和biological_process中;在cellular component (CC)中,差異基因主要富集在extracellular region中;在molecular function (MF)中,差異基因主要富集在catalytic activity中(圖3C)。
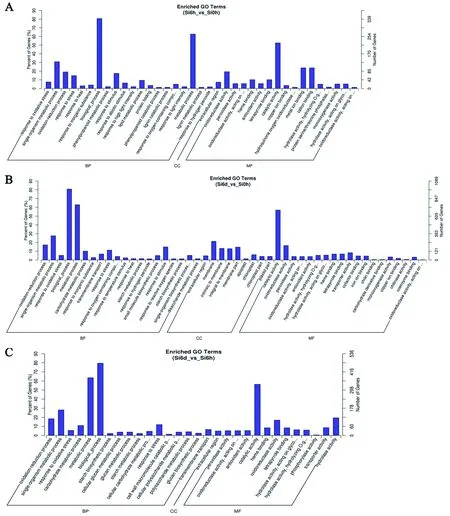
A:處理6 h;B:處理6 d;C:處理6 d和6 h。
2.3 差異基因KEGG代謝途徑分析
在生物體內,不同基因相互協調行使其生物學功能,通過KEGG (kyoto encyclopedia of genes and genomes) Pathway 顯著性富集能確定特異基因參與的最主要生化代謝途徑和信號轉導途徑。通過對差異基因的KEGG途徑分析發現,谷子處理6 h后的差異基因主要富集在Phenylalanine metabolism、Phenylpropanoid biosynthesis、Protein processing in endoplasmic reticulum和Biosynthesis of secondary metabolites途徑中(圖4A)。而處理6 d后、處理6 d和6 h的差異基因主要富集在Phenylalanine metabolism、Phenylpropanoid biosynthesis、Metabolic pathway和Biosynthesis of secondary metabolites途徑中(圖4B和圖4C)。
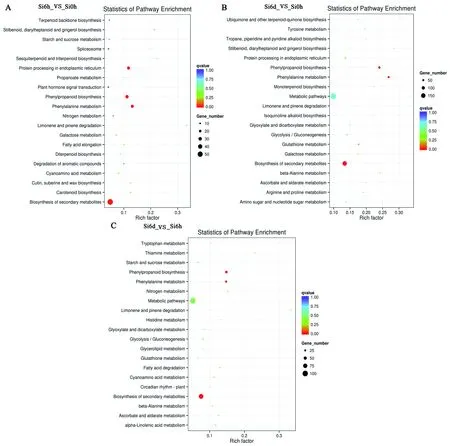
A:處理6 h;B:處理6 d;C:處理6 d和6 h。縱軸表示Pathway名稱,橫軸表示Rich factor (候選基因集所占背景基因的比例),點的大小表示此Pathway中差異表達基因個數多少。
3 討論與結論
干旱會導致植物細胞脫水及滲透脅迫,嚴重影響植物體內各種正常的生理及代謝過程。脅迫信號會被細胞表面的受體所感知,并啟動相關的信號轉導途徑[18-20]。在本研究中,谷子干旱脅迫不同時間后,我們鑒定到各種不同的類受體蛋白激酶(如Si030010m、Si029783m、Si000257m)的表達明顯上調。干旱脅迫會導致胞內Ca2+濃度增加,鈣信號被相關的Ca2+結合蛋白(如Si002992m、Si024895m、Si002992m)感知后啟動鈣信號轉導。脅迫信號通過激活一些蛋白激酶或者磷酸酶進一步向下傳遞[18]。促分裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)級聯途徑是真核生物中普遍存在的信號轉導模式,滲透脅迫能夠激活MPK3、MPK6和MPK4,干旱脅迫能夠激活AtMEKK1-MKK1-MPK4級聯途徑[21,22]。在谷子中我們同樣鑒定到一個MAPKKK(Si025330m)基因被明顯誘導激活。對于特定刺激的細胞響應主要依賴于MAPK激活的響應幅度和持續時間,因此這個過程受到精細的調控。蛋白磷酸酶是MAPK信號轉導途徑重要的負調控因子。PP2C型的磷酸酶AP2C1能夠滅活MPK4和MPK6[23]。在谷子干旱脅迫后有10個PP2C型蛋白磷酸酶的表達被激活,從而精細調控干旱脅迫信號轉導。
激活后的MAPK通過磷酸化下游的轉錄因子進而調控脅迫基因的表達。脅迫激活的MPK3/MPK6/MPK4能夠磷酸化WRKY和MYB轉錄因子[21]。最近的報道也發現,類受體蛋白激酶GUDK(GROWTH UNDER DROUGHT KINASE)磷酸化并激活轉錄因子OsAP37介導干旱脅迫信號轉導,激活后的OsAP37進一步誘導脅迫響應基因的表達來增強抗旱性和水稻產量[24]。本研究谷子在干旱脅迫后,多種轉錄因子(如DREB、NAC、WRKY、MYB、bHLH等)的表達被明顯誘導,這些結果與近兩年來谷子中轉錄因子家族鑒定的結果基本一致。激活后的轉錄因子通過結合脅迫響應基因啟動子中的順式調控元件,如DRE/CRT、ABRE、YCRS/MYBRS,進而調控基因的表達[18]。下游脅迫響應的功能蛋白,如LEA(late embryogenesis abundant protein)、不同分子量的HSPs(heat shock protein)、脫水素蛋白dehydrin、分子伴侶蛋白ClpB1(chaperone protein)等在干旱處理6 h和6 d后被明顯誘導。進一步分析發現,水孔蛋白(aquaporin)在干旱脅迫6 d后明顯下調,水孔蛋白的下調可能有助于保持植物體內的水分,防止水分的散失。泛素蛋白酶體途徑是真核生物體內重要的蛋白質降解途徑,E3泛素連接酶在該過程中能特異性識別并結合底物蛋白,從而調節靶蛋白的降解代謝。研究表明E3泛素連接酶在植物脅迫響應過程中起著重要的調控作用。擬南芥中的E3泛素連接酶DRIP1和DRIP2通過結合并泛素化DREB2A抑制其調控的干旱脅迫響應基因的表達[25]。在本研究谷子脅迫響應的后期(干旱處理6 d后),發現10個E3泛素連接酶基因的表達受到明顯調控,說明E3泛素連接酶介導的泛素化途徑在谷子脅迫響應過程中起著重要的調控作用。
本研究對干旱脅迫下的谷子進行了轉錄組分析,比較了不同時間處理后的差異表達基因,為進一步篩選并克隆谷子關鍵抗旱基因提供了依據,也為揭示谷子的抗旱機制奠定了基礎。
猜你喜歡
語數外學習·高中版中旬(2023年2期)2023-05-10 13:26:53
語數外學習·高中版中旬(2022年5期)2022-07-13 20:47:51
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中學生百科·大語文(2017年10期)2017-11-04 06:56:38
中國衛生(2016年3期)2016-11-12 13:23:26
公民與法治(2016年22期)2016-05-17 04:20:13
中國衛生(2014年12期)2014-11-12 13:12:52
癌變·畸變·突變(2014年6期)2014-02-27 06:15:03
