面向生物合成的代謝工程策略設計
2018-11-22 01:32:46馬紅武陳修來袁倩倩劉立明孫際賓
中國科學院院刊 2018年11期
馬紅武 陳修來 袁倩倩 劉立明,4* 孫際賓*
1 中國科學院天津工業生物技術研究所 天津 300308
2 江南大學 食品科學與技術國家重點實驗室 無錫 214122
3 江南大學 工業生物技術教育部重點實驗室 無錫 214122
4 江南大學 糧食發酵工藝與技術國家工程實驗室 無錫 214122
基因工程技術的出現使得人類不再受限于自然變異和篩選,通過有目的的基因組改造可以大幅度地提升菌株的生產性能。例如,通過對代謝網絡中酶基因的引入、敲除或精細調控可以創造新的細胞,將廉價的原料轉化為有價值的目標產品。這正是目前代謝工程和合成生物學研究的主要內容。從原料到產品的生物轉化途徑常常包括幾十步的酶反應,目前常用的代謝工程改造策略主要就是過表達和優化產物合成途徑中的關鍵酶、敲除副產物生成途徑、解除產物合成抑制等。通過這種傳統的代謝工程改造策略,人們已經獲得大量工程菌株應用于生物化學品(如氨基酸、有機酸、維生素、抗生素)、生物燃料(如乙醇)、生物材料(如聚羥基脂肪酸酯、聚乳酸等)的生產,以可再生的原料和綠色的生產方式,推動人類工業經濟發展進入一個新的階段。
代謝工程研究遇到的主要瓶頸就是細胞代謝網絡的復雜性。自然存在的細胞是長期進化的結果,常包括上千個酶基因并具有復雜的調控機制,僅僅對原料到產品合成途徑上的少數基因進行改造很多時候并不能達到預期的結果。隨著基因組測序技術的快速發展,完成全基因組測序的生物越來越多;通過基因組注釋信息可以快速確定其基因組能夠編碼哪些酶、進行哪些反應,由此構建出該生物的代謝網絡模型[1]。這些基因組規模代謝網絡模型和長期積累形成的跨物種代謝反應數據庫,如 KEGG 和 MetaCyc 等為代謝網絡計算模擬和設計奠定了基礎,為合成生物學和代謝工程提供了新的策略,從而大幅度加快了菌種定向開發和工業化應用進程。
1 合成新產品:從無到有
通過引入外源基因使某一生物合成其原來不能合成的新產品,這是一種常用的代謝工程手段。例如,Galanie 等[2]通過將來源于 6 種不同生物(包括植物、哺乳動物和細菌)的 10 余個基因引入酵母中實現了鴉片類藥物的人工合成。通過微生物細胞工廠合成植物來源的天然產物,其好處是微生物生長更快,單位時空內產量更高,可以通過較小的發酵工廠代替大面積的農業種植,大大降低生產成本。
從無到有的設計首先需要確定在底盤細胞中引入哪些外源基因才能打通該新產品的合成途徑。因此,需要將底盤細胞代謝網絡與代謝反應數據庫(如 KEGG、MetaCyc 等)相結合,通過一定的計算方法找出需要在底盤細胞中引入的新反應,形成新的途徑,以實現新產品的合成。例如,Zhang 等[3]將 KEGG 反應引入到大腸桿菌代謝網絡中,構建了包含 7 316 個反應的復合代謝網絡模型;基于該模型,以葡萄糖為底物進行通量平衡分析發現,1 777 個異源產品可以通過在大腸桿菌中引入適當外源反應合成。這一研究為選擇合適外源基因構建大腸桿菌的人工細胞工廠提供了指導。該方法還可以計算其他微生物為底盤時的結果,并通過比較分析確定針對某一特定產品的最優底盤生物(需引入外源基因少,相應前體合成能力強等)。Chatsurachai 等[4]分別以大腸桿菌、谷氨酸棒桿菌和釀酒酵母為底盤細胞,通過逐級擴展算法將來源于 KEGG、BRENDA 和 ENZYME 3 個數據庫中的異源代謝物和反應添加到 3 個菌株代謝模型中以合成新的產物。該擴展算法首先從數據庫中找出那些反應物在(但產物不在)該菌株代謝網絡中的反應,通過將這些反應添加到模型中,使該模型可以生成那些新反應中的產物。隨后在擴展的模型基礎上重復該新反應添加過程,直到沒有新反應可以添加。最終,針對每一個底盤細胞得到其可以合成的新產品列表及生成某一新產品需要引入的新反應。作者通過該方法預測出大腸桿菌和釀酒酵母是 1,3- 丙二醇合成的可行宿主,并且只需要在這兩種株菌中引入甘油脫氫酶和 1,3- 丙二醇氧化還原酶便可實現產品合成。
上述方法均需基于特定代謝反應數據庫擴展特定底盤細胞的代謝網絡,要合成的代謝物和相關反應必須存在于數據庫中才能設計出相關途徑,而自然界中還存在許多未知的生化反應并未包含在這些數據庫中。例如,很多酶,如醛縮酶等均具有廣譜催化活性,除了已知反應外還可能催化其他代謝物的轉化。通過酶改造,構建具有新催化活性的酶可以實現新的轉化過程獲得新的生物產品,甚至是合成出自然界不存在的全新生物分子。針對這一問題,人們提出了基于生化反應規則和化合物結構特征來設計新反應的方法。例如,Hatzimanikatis 的研究組基于化學信息學,在 KEGG 數據庫中反應的基礎上擴展酶反應,創建了 ATLAS 數據庫[5],其中包含 13 萬多個在已知反應基礎上通過結構相似性設計得到的新生化反應。利用該數據庫,他們設計了多條可以生成生物燃料丁酮的全新途徑,并對不同途徑的理論得率進行了比較分析,以獲得最優途徑[6]。
2 提高產品得率:從有到優
在賦予了細胞新的合成能力,實現了從無到有的改造之后,代謝工程的下一個目標就是以最小的原料消耗生產最多的產品。消耗單位原料(底物)可以生成的產物的量常用“得率”來表征,其是反映原料成本的一個重要指標,尤其對大宗化學品來說原料成本是影響產品生產成本的最主要因素。對某些通過簡單途徑就可以合成得到的產品,如乙醇、乳酸等,通過直觀觀察就可以得到得率最大化的代謝途徑;但對大部分代謝產物,如核苷酸、芳香族氨基酸等,由于其合成途徑復雜,涉及副產物及多個前體,很難通過簡單觀察或手工計算得到其理論得率。此外,在產品合成過程中還涉及能量(ATP)和還原力的產生和消耗,需要額外消耗底物產生能量和還原力,或通過某種途徑消耗掉多余的能量還原力,這使得產物得率的計算更為復雜。基于代謝網絡模型,人們提出了通量平衡分析(Flux Balance Analysis,FBA)的計算分析方法,以準確計算該網絡中產物的理論得率及能達到該理論得率的最優轉化途徑[7]。FBA 基于代謝網絡模型中的生化反應計量關系(可以理解為一個反應中各種反應物和產物的摩爾比)和反應的不可逆性約束,通過求解約束最優化問題,得到使產物生成最大化并且滿足物料和能量平衡的產品合成最優途徑。由于求得的途徑是通過對整體網絡計算分析得到的,比由生化知識得出的常規合成途徑更為全面,該方法能考慮能量平衡、輔因子平衡、多前體的平衡以及合成途徑中副產物的再回收利用[8]。該方法可以獲得實現同樣最大得率的多種不同策略,可能超越我們對生物化學知識的理解,得到與已知傳統代謝途徑不同的新途徑,從而提供全新的改造靶點和策略。例如,很多生物產品都是以乙酰輔酶 A 為前體合成得到,但以葡萄糖為底物時,乙酰輔酶 A 一般由糖酵解終產物丙酮酸脫羧生成,這導致葡萄糖中的 2 個碳原子都以 CO2形式損失掉而不能進入產物,1 個葡萄糖(含 6 個碳原子)最多得到 2 個乙酰輔酶 A(共 4 個碳原子)。因此,即使全部乙酰輔酶 A 中的碳都轉化到產品中,產物的最大碳摩爾得率也只能達到 0.67(圖 1a)。而以大腸桿菌代謝網絡模型 iJO1366 通過 FBA 對乙酰輔酶 A 衍生產品聚 3- 羥基丁酸酯(P3HB)最優得率計算結果卻可達到 0.86,這表明在其代謝網絡存在一些新途徑可以減少碳損失。求得的最優途徑中的通量分布如圖 1b 所示,可見在該途徑中除了常規葡萄糖經過 EMP 途徑生成丙酮酸進而生成乙酰輔酶 A 途徑外,還有一條稱之為蘇氨酸循環的途徑回收甲酸和 CO2,由磷酸烯醇式丙酮酸再回到丙酮酸同時生成一個乙酰輔酶 A 用于產品合成,從而提高了產品得率。基于該途徑計算結果,Lin 等[9]在大腸桿菌中通過解調控等激活了該循環途徑,將 P3HB 得率提高了將近一倍。
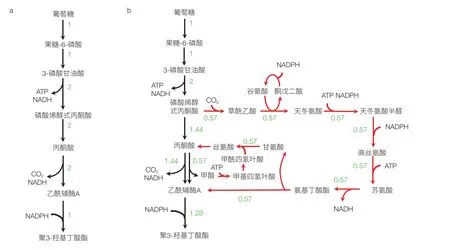
圖1 由代謝網絡分析確定聚3-羥基丁酸酯(P3HB)合成最優途徑
外源基因的引入不但可以使細胞從無到有獲得合成新產品的能力,也可以形成新的途徑提高已有產品的得率。Bogorad 等[10]通過在大腸桿菌中引入來源于 B.adolescentis 的戊糖/己糖磷酸轉酮酶(Fxpk)構建了非氧化酵解途徑(non-oxidative glycolysis,NOG)。通過該途徑可以實現 1 個葡萄糖生成 3 個乙酸,即完全沒有碳損失(圖 2)。我們在大腸桿菌 iJO1366 模型中引入該反應后通過 FBA 分析對上百種生物產品的理論得率進行了計算,發現至少有幾十種生化產品的得率都有了顯著提高。在這一計算結果指導下,構建了含 NOG 途徑的大腸桿菌菌株并分別用于 P3HB、丙酮和異丙醇的生產。改造后的菌株與出發菌株相比得率均有顯著提高[11,12]。最近 Meadows 等[13]將 NOG 途徑引入到釀酒酵母細胞中構建了法尼烯合成細胞工廠,最終菌株法尼烯對葡萄糖的碳摩爾得率由 0.52 提高到 0.65。
3 代謝網絡精細優化:動態調控
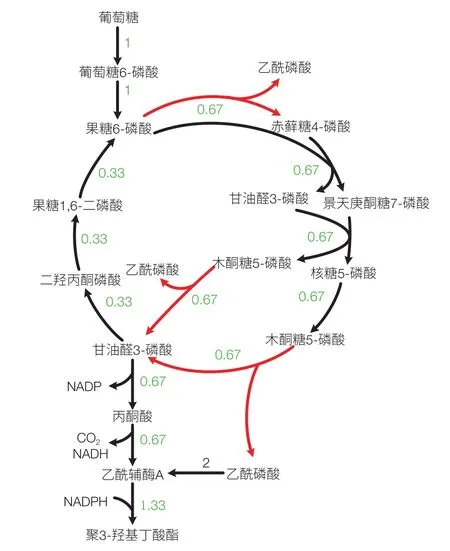
圖2 引入NOG途徑后乙酰輔酶A衍生產品合成最優途徑
基于 FBA 模型獲得的反應列表及其通量代表了理想狀態下反應速度最佳比例,可以作為代謝工程改造的終極目標——酶的實際表達量與活性要剛好滿足實現最佳代謝通量的要求,不多也不少。實際代謝工程實踐中,容易做到酶活性的“有無”調控或者叫“靜態控制”,比如通過基因敲除徹底失活某個生化反應,或者通過外源基因的引入增加某個反應,但是難以做到按照 FBA 模型預測的通量要求精確地調控每個酶的活性大小。實際上,細胞在長期進化中已經形成多種多樣的動態調控機制,根據不同的環境,精準地調控酶的表達(誘導與阻遏),精準地控制酶的活性(激活與抑制),確保代謝網絡能夠適應環境,實現全局最優化。例如,在發酵培養過程中細胞適應環境變化不斷調整自身的代謝狀態,形成延遲期、指數生長期和產物生成期等不同階段,每個階段對基因蛋白表達水平、代謝物濃度和通量分布都有不同要求。學習自然進化形成的動態調控機制,近年來人們提出了動態代謝工程(Dynamic Metabolic Engineering)的概念和方法,即根據胞內狀態和環境條件的變化對基因表達和活性進行動態調控,避免中間代謝產物的積累或不足,滿足不同階段的生長和產物合成對代謝通量分布的差異化需求。基因回路或稱轉錄調節回路,是最常用的動態調控手段,借助于小分子與轉錄因子(一種蛋白質)的相互作用和轉錄因子與特殊DNA區域(如啟動子、增強子、沉默子)的相互作用,將胞內外環境信號轉化為對酶蛋白表達的精準控制,確保細胞隨時調節代謝活動以適應環境變化[14-16]。通過動態控制酶的水平,可以實現代謝途徑流向的動態轉向調控(圖 3a),或實現代謝活性的精準平衡(圖 3b)。
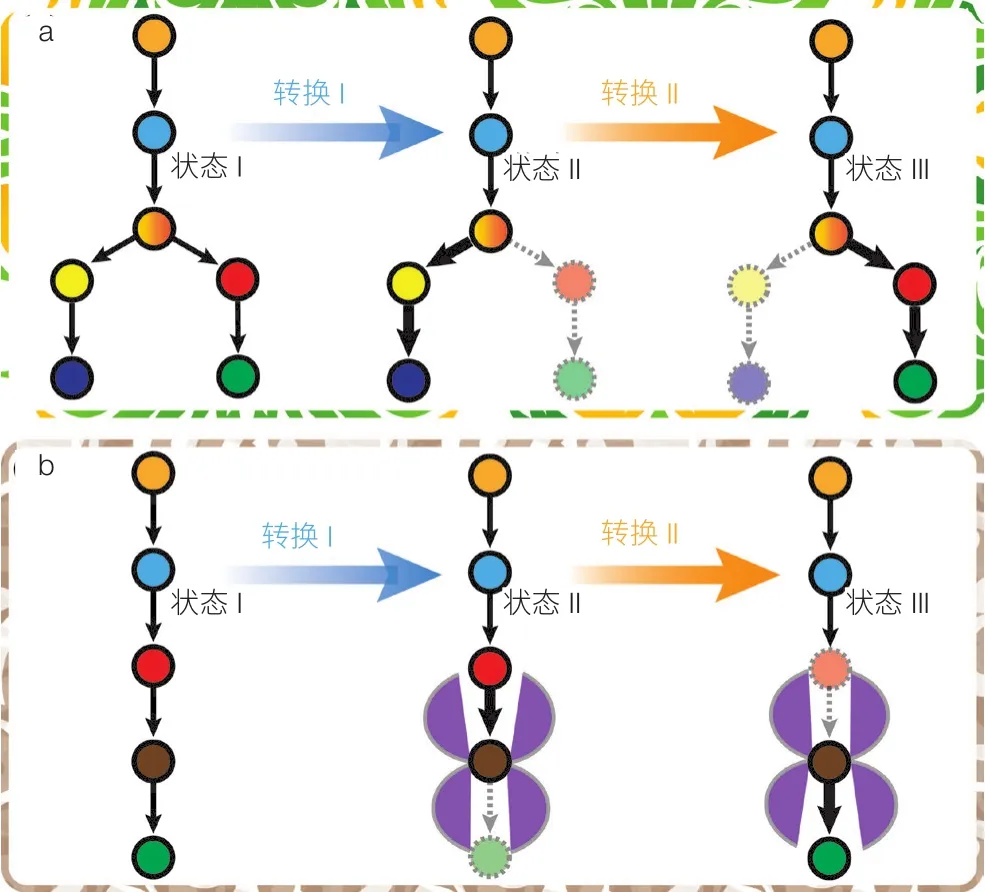
圖3 代謝途徑中代謝流的動態調控
基因回路可以在特定的時間和細胞密度下關閉或開啟基因表達,實現代謝流向在細胞生長的內源代謝途徑和化學品生產的異源途徑之間的切換。切換控制主要集中在生長必需基因上,因為通過它們可以實現生物量形成和產品生物合成之間最直接的控制。基因回路除了可以用于控制酶的合成速度,還可以精細調節酶的降解速度[17]。例如,葡萄糖-6-磷酸(G6P)是一個代謝流控制的重要節點,既可以通過磷酸果糖激酶-1(Pfk-1)進入糖酵解途徑從而進入內源代謝,也可以通過合成途徑進入異源代謝。通過基因回路控制 Pfk-1 的活性就可能將碳代謝流的流向從“生長模式”切換到“生產模式”。為實現這一目的,Brockman 和 Prather[18]設計了一種蛋白質降解回路,將 Pfk-1與 SsrA 降解標簽融合,SsrA 標簽能夠控制適配蛋白 SspB 的結合、ClpXP 蛋白酶招募和蛋白水解降解,從而實現 Pfk-1 降解速度的動態控制。將該回路用于大腸桿菌生產肌醇,與靜態控制相比,肌醇的積累量提高了 2 倍。基因回路通過控制必需基因的表達水平實現代謝途徑的動態調控,成為工業菌株代謝工程設計改造的有效手段:通過對內源代謝網絡進行重新布線,替代傳統靜態控制,實現基因表達的時間依賴性控制以及多個基因或途徑的平衡互作等,擁有廣泛的應用前景[17]。
代謝中間產物的過量積累可能引發副反應,占據重要細胞資源(如磷酸鹽、輔因子),以及產生直接或間接的細胞毒性,對細胞生長或產物合成不利。基因回路可以借助對中間產物濃度響應的生物傳感器,激活代謝途徑下游酶,阻遏代謝途徑上游酶,實現中間代謝物合成和利用速度適配,提高整個代謝途徑的合成效率。例如,莽草酸是芳香族氨基酸代謝途徑的中間體,莽草酸的積累會與芳香族氨基酸的代謝產生競爭。可以通過響應莽草酸的基因回路控制莽草酸代謝途徑關鍵酶——莽草酸激酶(AroK)實現代謝途徑各分支的平衡,實現碳代謝流在莽草酸積累與芳香族氨基酸代謝之間的平衡。Mahr 等[19]通過篩選和組裝適當的生物傳感器元件,包括半乳糖響應啟動子和苯丙氨酸響應啟動子,構建了代謝產物響應回路網絡。該回路的應用使得工程菌株 E. coli K-12 MG1655/pJC1-mtr sensor-type1 的苯丙氨酸產量與出發菌株 E. coli K-12 MG1655 相比提高了 4.3 倍。除了直接響應中間代謝物濃度以外,細胞生長狀態、輔因子失衡程度等都可以用于構建生物傳感器。
生物合成速度的最大化取決于單位菌體生產能力和菌體總量。通常情況下,生長和生產競爭使用同樣的細胞資源,需要進行合理的安排和均衡,才能實現生物合成速度整體的最大化。能夠對細胞密度進行響應的基因回路——群體感應系統(quorum sensing,QS)為調控生長和生產的關系提供了新的手段。例如,科研人員借助特征性啟動子和不同強度核糖體結合位點,組裝源自 Pantoea stewartii 的部分量子感應元件,實現了在特定時間和細胞密度下 pfk-1 等基因表達的開與關,工程菌株E. coli L19S 的肌醇產量與靜態控制相比提高了 5.5 倍[20]。借助QS回路控制aroK表達的工程菌株E. coli L19SA,其莽草酸的產量由 0 提高到 105 mg/L[20]。
4 結語
代謝網絡模型及基于代謝網絡模型的計算設計方法為代謝工程菌株改造提供了新的方法和策略。通過這些計算設計方法,人們可以選擇合適的外源基因構建新途徑實現從無到有的新產品合成,也可以確定改造靶點優化已有產品的得率,或者通過基因回路將細胞發酵培養過程中的狀態環境變化與特定基因的表達相關聯,使細胞適應不同的目標而始終保持最優狀態,最大化目標產物的生成。隨著人們對細胞機理和相關調控機制的了解不斷深入,有可能建立全細胞模型對細胞內代謝物、蛋白和基因表達水平的變化進行精確描述和準確預測,實現完全模型指導下的代謝工程策略設計。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
航空世界(2020年10期)2020-01-19 14:36:20
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
Coco薇(2015年1期)2015-08-13 02:23:50
玩具(2009年10期)2009-11-04 02:33:14
個人電腦(2009年9期)2009-09-14 03:18:46
