庫爾勒香梨PsiERF基因的克隆和表達分析
2018-12-11 06:59:46徐航全紹文馬麗周麗牛建新
新疆農(nóng)業(yè)科學(xué) 2018年9期
徐航,全紹文,馬麗,周麗,牛建新
(石河子大學(xué)農(nóng)學(xué)院園藝系/特色果蔬栽培生理與種質(zhì)資源利用兵團重點實驗室,新疆石河子 832003)
0 引 言
【研究意義】庫爾勒香梨(Pyrus×sinkiangensisT.T.Yu),屬薔薇科,是新疆梨的一個古老栽培品種。庫爾勒香梨果實具有果香濃郁,口感細膩,石細胞少且耐貯藏等特點[1]。由于遺傳因素的影響,庫爾勒香梨存在嚴重的萼片宿存現(xiàn)象,極大的影響了香梨的外觀及品質(zhì)。AP2/ERF是一個龐大的轉(zhuǎn)錄因子基因家族,含有由60~70個氨基酸組成的AP2/ERF結(jié)構(gòu)域,存在于所有的植物當中[2]。AP2/ERF超家族中的基因已經(jīng)在119~200種植物中被發(fā)現(xiàn)并進行了研究[3-6]。這個家族中的成員參與多種生物學(xué)過程,包括植物生長、花發(fā)育[7-8]、果實發(fā)育[9]、種子發(fā)育[10]、衰老[11]、病菌防御[12]、非生物逆境脅迫響應(yīng)[13]。另外,AP2/ERF類轉(zhuǎn)錄因子參與水楊酸、茉莉酸[14]、乙烯[15-18]、脫落酸[19]等多種信號轉(zhuǎn)導(dǎo)途徑,而且某些家族成員是逆境信號交叉途徑中的連接因子。根據(jù)其序列相似性和AP2/ERF結(jié)構(gòu)域的數(shù)量,將其分為四類,分別是AP2、ERF、和RAV家族和其他[20]。其中ERF家族包含1個AP2/ERF結(jié)構(gòu)域,可以分為2個大的亞家族,CBF/DREB亞家族和ERF亞家族[21]。研究表明,低溫和脫落酸都對庫爾勒香梨的萼片脫落宿存產(chǎn)生影響,這與ERF轉(zhuǎn)錄因子所調(diào)節(jié)的功能相吻合。因此,研究PsiERF轉(zhuǎn)錄因子的表達量對庫爾勒香梨萼片脫落與宿存的關(guān)系具有重要意義。【前人研究進展】前人的研究主要集中在栽培措施、生長勢、授粉品種[22-23]、生長調(diào)節(jié)劑[24-26]和礦質(zhì)元素[27]等生理層面。近些年來,對于這一問題的分子機制研究逐漸增多,其中有學(xué)者通過差異顯示PCR技術(shù)克隆了庫爾勒香梨中控制開花和激素調(diào)節(jié)相關(guān)的kfpMYB基因[28-30],Qi等[31]通過數(shù)字轉(zhuǎn)錄豐度的方法篩選了一些與光合作用、植物激素信號傳導(dǎo)、細胞壁修飾、轉(zhuǎn)錄調(diào)控和碳水化合物代謝等有關(guān)的可能影響庫爾勒香梨萼片脫落的基因,孫曉霞等[32]采用Illumina高通量測序技術(shù)發(fā)現(xiàn)了3條與萼片發(fā)育相關(guān)的Unigenes,田嘉等[33]分離和克隆了庫爾勒香梨果膠裂解酶PsPL。裴茂松等[34]對庫爾勒香梨中的NAC基因做了克隆及表達分析。【本研究切入點】基于實驗室前人的研究基礎(chǔ),利用庫爾勒香梨花器官轉(zhuǎn)錄組拼接的Unigenes,獲得一個完整的ERF基因cDNA序列,并對該基因進行了序列和結(jié)構(gòu)分析。【擬解決的關(guān)鍵問題】利用實時熒光定量PCR檢測了其在花期不同時期、不同器官,脫萼樣品和宿萼樣品的表達情況,對照前人對ERF轉(zhuǎn)錄因子的研究,研究Psi ERF轉(zhuǎn)錄因子對庫爾勒香梨萼片宿存與脫落的關(guān)系,為研究庫爾勒香梨萼片宿存和脫落分子機理奠定基礎(chǔ)。
1 材料與方法
1.1 材 料
樣品于4月取自新疆庫爾勒市沙依東園藝場,分別選取強勢樹與弱勢樹各3株作為試驗樹,于初花期、盛花期、末花期采集強勢樹上的第1序位(宿萼)和弱勢樹第4序位(脫萼)的全花各50朵,去除花瓣,分離子房和萼片,分別將兩部分用錫箔紙保存,投入液氮處理,后置于-80℃冰箱保存?zhèn)溆谩?/p>
1.2 方 法
1.2.1 生物信息學(xué)分析
利用NCBI BLAST進行核酸以及氨基酸序列的同源性比對,DNAMAN對核酸的序列進行編輯和分析,OFR Finder查找開放閱讀框,ExPASy-ProtParam toll和SignalP分析氨基酸序列的基本結(jié)構(gòu),氨基酸的親水性、等電點、信號肽等信息,PRABI Lyon Gerland 預(yù)測氨基酸序列的二級結(jié)構(gòu)。DNAMAN和MEGA7.0分析同源性及構(gòu)建系統(tǒng)發(fā)育樹。
1.2.2 RNA的提取和cDNA的合成
利用TIANGEN公司柱式植物RNAout1.0試劑盒提取庫爾勒香梨花樣品的總RNA,用NANODrop2000監(jiān)測RNA的濃度和純度(OD260/280比值),用1.2%的瓊脂糖凝膠電泳檢測RNA的完整性。利用Takara的M-MLV反轉(zhuǎn)錄酶和反轉(zhuǎn)錄引物B26(5'-GACTCGAGTCGACATCGATTTTTTTTTTTTTTTTTT-3')合成單鏈cDNA。
1.2.3 實時熒光定量PCR引物的篩選
對強勢樹第1序位全花(宿萼)和弱勢樹第4序位全花(脫萼)進行轉(zhuǎn)錄組測序共得到103個差異表達基因;對強勢樹第1序位萼片和弱勢樹第4序位萼片進行數(shù)字表達譜測序共得到64個差異表達基因;對強勢樹第1序位子房和弱勢樹第4序位子房進行數(shù)字表達譜測序共得到95個差異表達基因。比較轉(zhuǎn)錄組和表達譜的測序結(jié)果,得到二者共有差異表達基因10個,分別是植物生長素誘導(dǎo)蛋白5NG4(comp50752_c0)、聚半乳糖醛酸酶抑制蛋白PGIG(comp49798_c0)、β-半乳糖甘酶(comp49925_c0)、β-1,3-葡聚糖酶(comp43208_c0)、脫水響應(yīng)蛋白(comp44869_c0)、脫水反應(yīng)結(jié)合元件蛋白(comp49899_c0)、脂轉(zhuǎn)移蛋白前體(comp36582_c0)、NAC結(jié)構(gòu)域蛋白NAC002(comp41728_c0)和乙烯響應(yīng)轉(zhuǎn)錄因子ERF109(comp36863_c0)、ERF027(comp44254_c0)基因。從庫爾勒香梨轉(zhuǎn)錄組測序數(shù)據(jù)中調(diào)出拼接的ERF109即PsiERF的Unigene,提交到NCBI數(shù)據(jù)庫進行比對,該Unigene與白梨的(Pyrus ×bretschneideri)全長cDNA序列(LOC103946790)高度同源,根據(jù)其保守區(qū)用Primer Premier5.0軟件設(shè)計引物,序列為ERFP1:5'-AACTACTTCTCGCCATCGT-3'和ERFP2: 5'-TGTTCTTGCTCTTCCTCGT-3'。
以cDNA為模版,用PsiERF的特異引物ERFP1/ERFP2進行PCR。反應(yīng)體系為:2×TaqMix 12.5 μL,cDNA(模版)2 μL,引物ERFP1和ERFP2(μmol/L)各1 μL,其余用無菌水補足至25 μL。反應(yīng)條件:94℃預(yù)變性5 min;94℃變性40 s,54℃退火45 s,72℃延伸1 min,30各循環(huán)后72℃再延伸10 min。PCR產(chǎn)物分別進行回收純化后連接到pEASY-T1,轉(zhuǎn)化至DH5α,進行藍白斑篩選和菌液PCR鑒定,將陽性克隆進行測序,對測序結(jié)果進行比對和分析。
1.2.4 實時熒光定量PCR
用天恩澤公司的柱式植物RNAout 1.0分別提取初花期、盛花期、末花期3個時期強勢樹上第1序位的全花、萼片、子房和弱勢樹上第4序位的全花、萼片、子房的總RNA(用Thermo Scientific DNase Ⅰ去除DNA污染),并對每個樣品設(shè)置3個生物學(xué)重復(fù)。利用Takara的M-MLV反轉(zhuǎn)錄酶合成cDNA,用SYBR? Green Realtime PCR Master Mix(東洋坊,上海)進行實時熒光定量PCR,PCR程序為95℃預(yù)變性30s,95℃變性5s,54℃退火10s,72℃延伸15s,40個循環(huán),溶解曲線的溫度是65~95℃。所用的內(nèi)參引物為Actin(上游引物:5'- CCATCCAGGCTGTTCTCTC-3',下游引物:5'-GCAAGGTCCAGACGAAGG-3'),基因的相對表達量分析用2-△△CT法:ΔΔCt= (CT,Target-CT,Actin)Time x-(CT,Target-CT,Actin)Time 0。
2 結(jié)果與分析
2.1 序列分析
基于庫爾勒香梨轉(zhuǎn)錄組測序結(jié)果(庫爾勒香梨萼片脫落組和宿存組花的轉(zhuǎn)錄組測序數(shù)據(jù),已經(jīng)上傳到NCBI SRA 數(shù)據(jù)庫,登錄號:PRJNA303067,(http://www.ncbi.nlm.nih.gov/bioproject/303067))采用TMM(測試成熟度模型)對read count數(shù)據(jù)進行標準化處理,再用DEGseq進行差異分析(篩選閾值為qvalue<0.005且|log2FoldChange|>1),篩選出了一條具有ERF/AP2結(jié)構(gòu)域蛋白的Unigene PsiERF,將核苷酸序列提交到NCBI進行比對,結(jié)果顯示,其與白梨預(yù)測的乙烯轉(zhuǎn)錄響應(yīng)因子(LOC103964489)具有99%的相似度,與蘋果預(yù)測的乙烯轉(zhuǎn)錄響應(yīng)因子(LOC103401371)具有97%的相似度。序列分析結(jié)果顯示,PsiERFcDNA序列的長度是1 195 bp,包含一個編碼264個氨基酸長度為795 bp的開放閱讀框(ORF),一個123 bp的5,端非編碼區(qū)和一個277 bp的3,端非編碼區(qū)。用ProtParam分析可得,該基因編碼蛋白的分子組成為C1 253H1 967N371O407S7,分子量為28.965 06 kDa,等電點為5.78,不穩(wěn)定系數(shù)為47.05,總平均親水性值為-0.726,包含一個保守的AP2/ERF結(jié)構(gòu)域,推測此蛋白質(zhì)為不穩(wěn)定親水性蛋白質(zhì)。SignalP分析,PsiERF沒有跨膜和信號肽顯現(xiàn),為非分泌蛋白質(zhì)。 圖1
2.2 蛋白質(zhì)的二級結(jié)構(gòu)預(yù)測
通過PRABI Lyon Gerland對PsiERF基因的蛋白質(zhì)二級結(jié)構(gòu)進行預(yù)測。發(fā)現(xiàn)PsiERF的主要的二級結(jié)構(gòu)元件是不規(guī)則的盤繞(63.26%)、α-螺旋(23.86%)和延伸鏈(12.88%)。圖2
2.3 PsiERF基因在白梨基因組中的位置
通過分析PsiERF在白梨基因組中的位置發(fā)現(xiàn),PsiERF基因是白梨scaffold 520.0 基因組上246 130~247 324 nt長為1 194 bp的序列,無內(nèi)含子。
2.4 系統(tǒng)發(fā)育
通過BLASTP分析發(fā)現(xiàn)PsiERF的氨基酸序列與其它薔薇科植物的ERF具有高度同源性,如與白梨(Pyrus×bretschneideri, XP_009375704.1)98%、蘋果(Malus domestica,XP_008338309.1)85%、日本杏(Prunus mume,XP_008241498.1)53%、櫻桃(Prunus avium XP_021824713.1)50%、桃(Prunus persica,XP_007204172.1)51%等都具有一定的同源率,普遍達到了50%以上。通過DNAMAN對這6條氨基酸序列進行比對,發(fā)現(xiàn)它們的N端具有高度保守性。圖3
利用MEGA7.0軟件構(gòu)建PsiERF與其它17條包含AP2/ERF結(jié)構(gòu)域的物種氨基酸序列的系統(tǒng)進化樹,研究表明,PsiERF與白梨(XP009375704.1)蘋果(XP008338309.1)同屬一個亞群,并且于白梨(XP009375704.1)的同源性最高。圖4

加粗字體分別為起始密碼ATG和終止密碼TAG;下劃線所標部分為AP2/ERF結(jié)構(gòu)域保守區(qū);下劃線下斜體標示出的是ERF亞家族特征氨基酸丙氨酸(A)和天冬氨酸(D)
ATG(initiation codon)and TAG(termination codon)are bold;The conserved AP2/ERF domain is underlined; Alanine and asparaginic acid(typical amino acid of ERF subtribe) are italic
圖1 PsiERF基因的核苷酸序列及推定的氨基酸序列
Fig.1 Sequences of nucleotide and putative amino acid of the PsiERF gene

藍線代表α-螺旋,紫線代表不規(guī)則盤繞,紅線代表延伸鏈
The blue line represents the alpha helix,the purple line represents the random coil,the red line represents the extended strand
圖2 PsiERF 基因的蛋白質(zhì)二級結(jié)構(gòu)預(yù)測
Fig.2 Prediction of the protein secondary structure of PsiERF

黑色背景的白色字母表示一致的氨基酸,紅色背景字母表示一致性高于75%的氨基酸,藍色背景字母表示一致性高于50%的氨基酸
Identical amino acids are in white letters with black background. Red background indicates amino acid with ≥ 75% identity and the blue background indicates amino acid with ≥ 50% identity
圖3 PsiERF與其它含有AP2/ERF結(jié)構(gòu)域蛋白的多重比對
Fig.3 Amino acid sequence alignment of PsiERF and other proteins containing the AP2/ERF domain
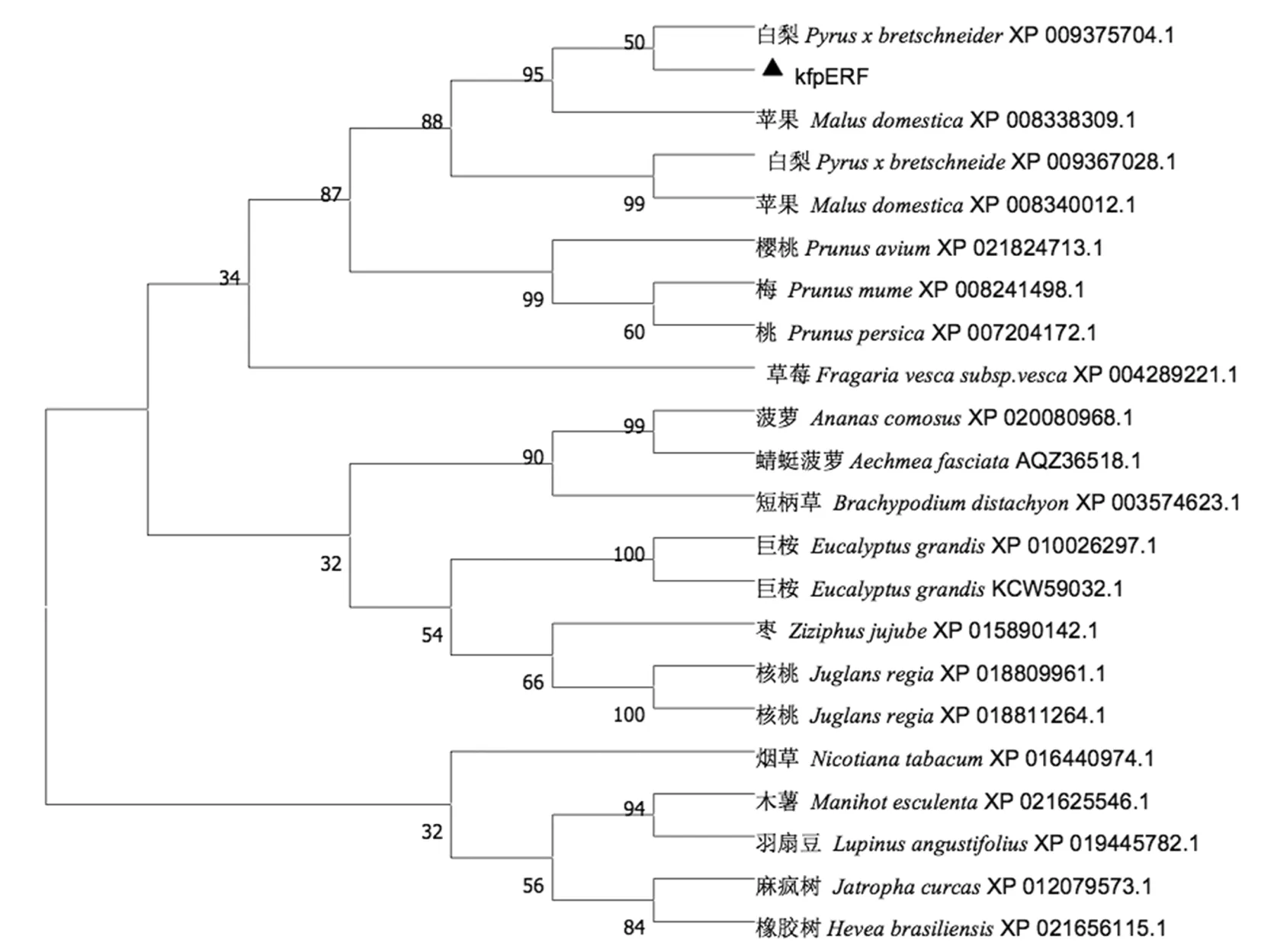
圖4 不同來源ERF氨基酸序列的系統(tǒng)進化樹
Fig.4 Phylogenetic tree of ERF amino acid sequence in different sources
2.5 實時熒光定量PCR
研究表明,PsiERF(ERF109)基因在宿萼組全花樣品中的表達量較脫萼組全花樣品高,而且在始花期和末花期表現(xiàn)出極顯著差異;該基因在兩組樣品中均是在始花期表達最高,且均極顯著高于盛花期和末花期,在末花期,宿萼組全花樣品中的表達量極顯著高于脫萼組全花樣品中的表達量;PsiERF基因在末花期萼片中的表達量極顯著高于始花期,在始花期和盛花期,子房中的表達量均高于萼片中的表達量,但是在末花期,萼片中的表達量極顯著的高于子房中的表達量。PsiERF基因在末花期脫萼組中子房和萼片中的表達無明顯差異,且顯著低于始花期和盛花期。圖5
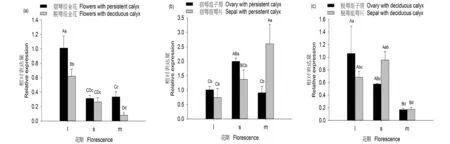
不同小寫字母表示在P<0.05水平具有顯著性差異,不同大寫字母表示在P<0.01水平具有極顯著性差異;l、s、m分別表示初花期、盛花期、末花期
Different small letters within a panel indicate significant differences atP<0.05, different capital letters within a panel indicate significant differences atP<0.01; l, s, m indicate early bloom, full bloom and late bloom respectively
圖5 PsiERF在脫萼組和宿萼組不同花器官全花、子房、萼片不同花期表達
Fig.5 Level of the relative expression of PsiERF in whole flowers,ovaries,and sepals with or without the calyx group
3 討 論
研究通過分析PsiERF(ERF109)基因的序列、結(jié)構(gòu)、蛋白質(zhì)結(jié)構(gòu)以及PsiERF基因在脫萼和宿萼樣本中全花、子房,萼片各時期的表達情況,以及前人關(guān)于AP2/ERF家族性質(zhì)以及庫爾勒香梨脫萼宿萼影響因素的相關(guān)研究,來分析PsiERF基因與庫爾勒香梨脫萼、宿萼這一生理現(xiàn)象有什么樣的關(guān)系。
ERF家族包含1個AP2/ERF結(jié)構(gòu)域,可以分為2個大的亞家族,CBF/DREB亞家族和ERF亞家族。DREB亞家族和ERF亞家族的主要區(qū)別在AP2/ERF結(jié)構(gòu)域的第14位和第19位氨基酸殘基,DREB亞家族的第14位和第19位氨基酸分別是纈氨酸和谷氨酸,而ERF亞家族是丙氨酸和天冬氨酸[20]。根據(jù)分析可知,PsiERF是屬于ERF亞家族中的成員。ERF亞家族成員可識別GCC盒(AGCCGCC),應(yīng)答生物脅迫和非生物脅迫,功能廣泛[35]。
調(diào)節(jié)植物組織衰老、脫落、死亡的激素有很多,如脫落酸、赤霉素、生長素、乙烯等[36]。研究表明,果萼中較高水平生長素和赤霉素以及較低水平的脫落酸有利于果萼宿存[37]。除此之外,影響植物組織的程序性死亡的因素還包括低溫這一非生物脅迫。有研究表明,低溫會促進香梨的脫萼[38]。ERF調(diào)控赤霉素的表達以及對低溫產(chǎn)生應(yīng)答。在水稻(Oryzasativa)中,ERF109的高表達會降低乙烯的合成,間接影響赤霉素的合成[39]。在佛手(Citrus me~ca L. vat. Sarcodactylis Swingle)中,ERF的高表達會增加植物對低溫的抗逆性[40]。實驗實時熒光定量顯示,PsiERF在宿萼組中全花樣品在始花期、盛花期、末花期的表達量都高于脫萼組全花樣本,并且在始花期和末花期表現(xiàn)出極顯著差異,并且在宿萼組末花期,萼片中PsiERF表達量極顯著高于子房而在脫萼組末花期,萼片與子房中PsiERF表達量低并且無顯著差異。由此推測,PsiERF基因決定香梨脫宿萼的關(guān)鍵時期是在末花期,且PsiERF低表達能夠促進萼片脫落。由宿萼組子房萼片樣本末花期數(shù)據(jù)可以看出,PsiERF在萼片中的表達量極顯著高于子房,PsiERF在萼片中的表達量高低是影響香梨萼片脫落和宿存的關(guān)鍵因素。綜上所述,庫爾勒香梨中,PsiERF基因能夠通過響應(yīng)調(diào)控影響植物程序性死亡的激素,由此來影響香梨的萼片宿存與脫落。
4 結(jié) 論
克隆鑒定一個新的庫爾勒香梨ERF109基因,命名為PsiERF。該基因cDNA序列長度為1 195 bp,編碼264個氨基酸序列,在第14位和第19位分別丙氨酸和天冬氨酸,是屬于AP2/ERF家族中的ERF亞家族。同源性分析結(jié)果表明,與白梨和蘋果的ERF109親緣較近,分別達到了95%與85%。實時熒光PCR結(jié)果顯示,PsiERF在宿萼組樣品中表達量均高于脫萼組,特別是在始花期與末花期呈現(xiàn)極顯著(P<0.01)差異。宿萼組末花期,萼片中PsiERF的表達量極顯著(P<0.01)高于子房中的表達量。脫萼組末花期,萼片中PsiERF的表達量和子房中的表達量無明顯差異。故推測該基因在萼片中表達的高低與庫爾勒香梨萼片宿存和脫落密切相關(guān)。
