棉花黃萎病拮抗細菌胞外酶檢測及其酶活測定
2018-12-11 06:59:46李雪艷張濤楊紅梅楚敏高雁曾軍霍向東張濤林青歐提庫爾李玉國婁愷史應武
新疆農業科學 2018年9期
李雪艷,張濤,楊紅梅,楚敏,高雁,曾軍,霍向東,張濤,林青,歐提庫爾,李玉國,婁愷,史應武
(1.新疆大學生命科學與技術學院,烏魯木齊 830052;2.新疆農業科學院微生物應用研究所/新疆特殊環境微生物實驗室,烏魯木齊 830091)
0 引 言
【研究目的】棉花是新疆主要經濟作物,隨著棉花種植區的連年輪作,黃萎病日益嚴重,對棉花的產量和質量損害巨大。大麗輪枝菌(Verticilliumdahliae)是棉花黃萎病的主要致病菌,該真菌類型多、變異快、菌系與寄主關系復雜[1,2]。防治這種土傳病害通常使用化學殺菌劑、高抗育種、生物防治等方法,化學殺菌劑只能治標不治本,對土壤中的致病孢子作用小,而且污染環境;高抗育種年限長且適應新疆棉田的高抗棉株品種少、安全的防控土傳疾病,而且不會影響其他農業系統的有益微生物[3,4],符合綠色可持續發展的理念。研究發現,真菌[5]、細菌[6,7]、放線菌[8]等部分微生物可降低棉花黃萎病發病率,生防機理主要有營養與生態位的競爭,拮抗物質,誘導植物抗病性等[9],抗菌物質中各種酶類起到關鍵作用。真菌細胞壁的主要成分有幾丁質、葡聚糖、纖維素、蛋白等,研究4株拮抗菌產幾丁質酶、葡聚糖酶、纖維素酶、蛋白酶可一定程度解釋真菌細胞腫脹和細胞壁完整性缺失,導致細胞的裂解和死亡的原因[10],為進一步開發應用該4株拮抗菌株提供依據,為4株拮抗細菌的防治機理研究奠定基礎。【前人研究進展】國內外目前已有關于微生物胞外酶防治病害的報道,蘇明慧等[11]發現從芽孢桿菌FM4B分離的幾丁質酶可抑制西瓜枯萎病菌,酶液濃度越高抑制效果越好;田亞琴等[12]發現幾丁質酶、葡聚糖酶與芒果炭疽菌抑制效果相關;Li等[13]發現幾丁質酶、葡聚糖酶能水解植物病原菌細胞壁從而抑制病原真菌;文鳳云等[14]發現葡聚糖酶是多粘類芽孢桿菌CP7拮抗真菌活性組分或其中之一,并克隆葡聚糖酶基因構建高效表達菌。劉艷[5]報道哈茨木霉4種離體蛋白酶對尖孢鐮刀菌、立枯絲核菌、核盤菌、鏈格孢菌和楊樹爛皮病菌有不同程度的抑制作用。【本研究切入點】目前棉花黃萎病防治效果不理想。已通過電鏡掃描驗證4株拮抗細菌均可破壞大麗輪枝菌細胞形態,但其拮抗機理尚不明確。研究棉花黃萎病拮抗細菌胞外酶檢測及其酶活測定。【擬解決的關鍵問題】通過檢測培養基,透明圈法測定胞外酶的種類,經酶類發酵培養基富集,DNS法測定纖維素酶、幾丁質酶、葡聚糖酶活,福林-酚法測定蛋白酶活。
1 材料與方法
1.1 材 料
1.1.1 菌種來源
BHZ-29、SHZ-24菌株由實驗室分別從博樂、石河子棉株體內篩選;SHT-15、SMT-24均從石河子棉株種植根際土壤中分離所得。
1.1.2 種子液培養基
NB培養基。
1.1.3 鑒別培養基
葡聚糖鑒別培養基:參考文獻[15];纖維素酶檢測培養基:CMC-Na 1 g,(NH4)2SO40.4 g,KH2PO40.1 g,MgSO40.05 g,GaCl20.002 g,蛋白胨0.1 g,酵母膏0.1 g,瓊脂2 g,蒸餾水100 mL,pH中性,0.1%剛果紅備用;蛋白酶檢測培養基:參考文獻[10]海水更改為蒸餾水;膠體幾丁質酶檢測培養基:參考文獻[16]。
1.1.4 發酵培養基
纖維素酶發酵培養基:不加瓊脂的纖維素檢測培養基,pH中性;葡聚糖酶發酵培養基:無瓊脂及染料的葡聚糖檢測培養基,酵母膏2 g,pH中性;膠體幾丁質酶發酵培養基:膠體幾丁質0.2 g,酵母膏0.6 g,K2HPO40.1 g,KH2PO40.15 g,GaCl2·2H2O 0.1 g,MgSO40.02 g,蒸餾水100 mL,pH中性;蛋白酶發酵培養基:可溶性淀粉1 g,蛋白胨0.5 g,Na2HPO4·12H2O 0.4 g,KH2PO40.03 g ,GaCl20.1 g,蒸餾水100 mL,pH中性。
1.1.5 主要試劑
膠體幾丁質參考[17,18]自制,3,5-二硝基水楊酸(DNS)試劑參考[17]配制,0.1 mol/L pH值為4.5的醋酸-醋酸鈉緩沖液、0.02 mol/L pH值為7.5的磷酸緩沖液均參考文獻[18]配制,其余均為分析純。
1.2 方 法
1.2.1 拮抗菌種子液的制備
將NA上劃線分離的拮抗菌單菌落接種到NB液體培養基,30℃、200 r/min振蕩發酵1 d。
1.2.2 粗酶液的制備
將4種拮抗菌的種子液按照分別按照4%的量接種到葡聚糖酶、纖維素酶、幾丁質酶、蛋白酶發酵培養基中,500 mL三角瓶裝液量為50 mL,葡聚糖酶、纖維素酶發酵培養基30℃、200 r/min振蕩培養12、36、60、84和108 h,幾丁質酶、蛋白酶同條件下發酵培養12、36、60、84、108和132 h。10 000 r/min離心15 min,取上清液備用。
1.2.3 拮抗菌產幾丁質酶、蛋白酶、葡聚糖酶、纖維素酶平板檢測
分別在葡聚糖酶、幾丁質酶、纖維素酶、蛋白酶檢測培養基上打5孔(直徑6 mm),四周呈正方形對稱打4孔為處理組,每孔加拮抗菌種子液100 μL,培養基中心孔為對照組,加100 μL無菌NB培養基,放置30℃恒溫培養箱培養48 h,將葡聚糖檢測培養基與纖維素檢測培養基用0.1%的剛果紅溶液10~15 mL染色20 min,傾倒染液后加入1 mol/L的NaCl至滿平板,靜置20 min,反復NaCl洗3次,測量記錄透明圈外徑大小。
1.2.4 標準曲線的建立
稱取適量無水葡萄糖、N-乙酰氨基葡萄糖、L-酪氨酸放置于105℃烘箱內干燥至恒重,分別準確稱取 1.000 g,配制成1 mg/mL的葡萄糖標準液,1 mg/mL N-乙酰氨基葡萄糖標準液,0.1 mg/mL的L-酪氨酸標準液。
葡萄糖標準曲線的建立參考文獻[19]稍作修改,葡萄糖標準液加入量改為0、0.1、0.2、0.3、0.4、0.5、0.6 mL,DNS溶液加入量更改為3.0 mL,蒸餾水定容至10 mL。N-乙酰氨基葡萄糖標準曲線的建立同上,將葡萄糖標準液更改為N-乙酰氨基葡萄糖標準液,加入量為0.25、0.30、0.35、0.40、0.45、0.50、0.55 mL,補充蒸餾水至1.5 mL,DNS加入量1.5 mL,最終定容體積為7.5 mL。L-酪氨酸標準曲線建立參考文獻[20]。
1.2.5 拮抗菌幾丁質酶、蛋白酶、葡聚糖酶、纖維素酶活測定
0.5%CMC-Na 、1%葡聚糖、0.5%膠體幾丁質用0.1 mol/L pH為4.5的醋酸-醋酸鈉緩沖液配制,0.5%的酪素用0.02 mol/L pH為7.5的磷酸緩沖液配制。采用DNS法測定葡聚糖、纖維素、幾丁質酶活性,在540 nm波長處記錄樣品OD值;采用福林-酚法測定蛋白酶活性,在680 nm波長處記錄OD值,均以滅活粗酶液為對照組。分別將樣品細菌計數,稀釋105、106、107梯度涂布,每個梯度3個平行試驗。
1.2.5.1 纖維素酶活性測定
1 mL 0.5%CMC-Na與1 mL纖維素粗酶液混合50℃水浴30 min,加3 mL DNS,100℃煮沸5 min,冷卻補充體積至10 mL。
1.2.5.2 葡聚糖酶活性測定
1 mL 1%葡聚糖與1 mL葡聚糖粗酶液,混合50℃水浴30 min,加3 mL DNS,100℃煮沸5 min,冷卻補充體積至10 mL。
1.2.5.3 幾丁質酶活測定
參考文獻[17]稍作修改,1 mL 0.5%膠體幾丁質與1 mL幾丁質粗酶液50℃水浴30 min,10 000 r/min離心15 min,取離心上清液1.5 mL與1.5 mL DNS溶液沸水浴5 min,冷卻后定容7.5 mL。
1.2.5.4 蛋白酶活性測定
參考文獻[20]稍作修改,1 mL蛋白質粗酶液與1 mL 0.5%的酪素混合,37℃水浴20 min,加1 mL三氯乙酸終止反應,10 000 r/min離心15 min后取上清液0.5 mL補充蒸餾水0.5 mL,加0.55 mol/L的碳酸鈉溶液5 mL,最后加福林酚溶液1 mL,37℃水浴顯色20 min。
1.2.6 酶活定義
底物在一定反應條件下(葡聚糖、幾丁質、纖維素50℃下水浴30 min,酪蛋白37℃水浴20 min)與粗酶液反應30 min生成1 μg葡萄糖/酪氨酸為一個酶活單位。
2 結果與分析
2.1 平板透明圈法測定SMT-24、BHZ-29、SHT-15、SHZ-24幾種胞外酶
研究表明,BHZ-29、SHT-15、SMT-24菌株均產葡聚糖酶、纖維素酶、蛋白酶,不產幾丁質酶;SHZ-24產幾丁質酶、葡聚糖酶、蛋白酶,不產纖維素酶。檢測平板上蛋白酶、幾丁質酶、纖維素酶解圈清晰透明,邊緣界限明顯,葡聚糖酶解透明圈菌株SHZ-24、SHT-15清晰,BHZ-29、SMT-24邊緣模糊,界限不清。圖1
研究表明,SHZ-24菌株幾丁質酶解圈與其他處理組間差異顯著,酶解外徑為(14.85±0.15) mm。蛋白酶透明圈外徑大小均與對照組差異極顯著,SHZ-24菌株蛋白酶解圈最大,達(28.85±0.37) mm;葡聚糖酶解圈外徑大小組間差異均顯著,SMT-24菌株葡聚糖酶解圈最大,達(36.50±0.36) mm。BHZ-29、SMT-24、SHT-15菌株之間纖維素酶透明圈外徑大小對照組差異顯著,最大為(17.50±0.42) mm。表1

注:1~4分別代表SMT-24、BHZ-29、SHT-15、SHZ-24,CK為空白對照
Note: 1-4 respectively represent SMT-24, BHZ-29, SHT-15, shz-24,CK as blank control
圖1 平板透明圈法初測結果
Fig.1InitialtestresultsofPlatecultureandclearzonesmethods
表1 酶解透明圈外徑及方差
Table 1 Analysis of size and variance of enzymatic transparent circles
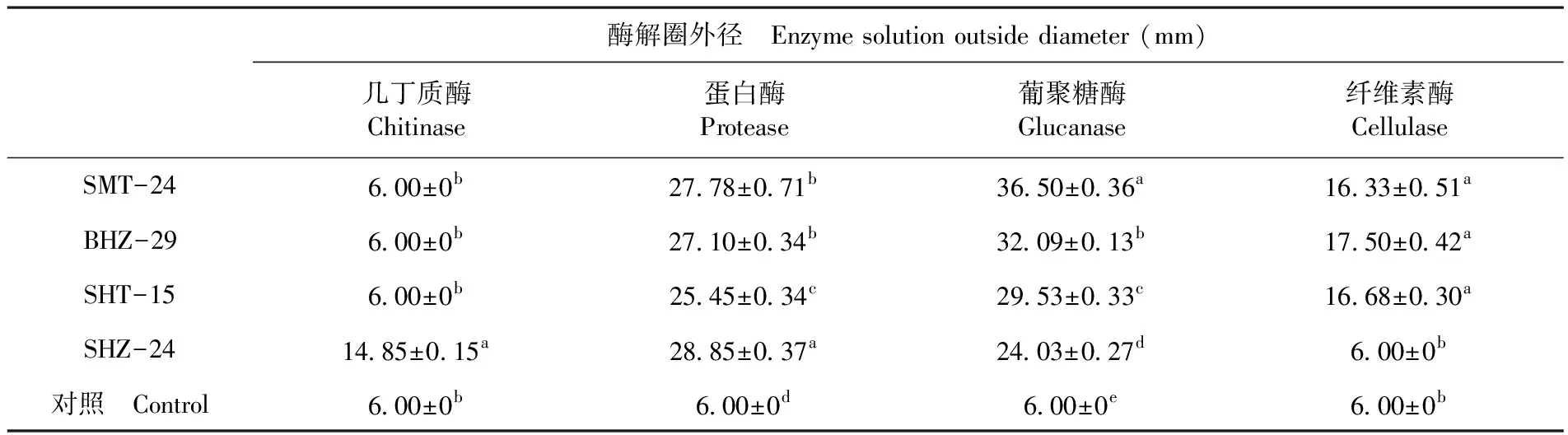
酶解圈外徑 Enzyme solution outside diameter (mm)幾丁質酶Chitinase蛋白酶Protease葡聚糖酶Glucanase纖維素酶CellulaseSMT-246.00±0b27.78±0.71b36.50±0.36a16.33±0.51aBHZ-296.00±0b27.10±0.34b32.09±0.13b17.50±0.42aSHT-156.00±0b25.45±0.34c29.53±0.33c16.68±0.30aSHZ-2414.85±0.15a28.85±0.37a24.03±0.27d6.00±0b對照 Control6.00±0b6.00±0d6.00±0e6.00±0b
注:不同的字母代表同一種酶不同菌種間的差異顯著(P<0.05)
Note: Different letters represent a significant difference between different strains of the same enzyme (P<0.05)
2.2 標準曲線建立
葡萄糖標準曲線、N-乙酰氨基葡萄糖標準曲線、L-酪氨酸標準曲線對應線性方程分別為y1=13.289x1-0.115 6、y2=11.111x2-0.130 3、y3=0.070 6x3-0.002,相關性系數分別為0.998 6、0.998 9、0.999,線性關系良好。
2.3 SMT-24、BHZ-29、SHT-15、SHZ-24拮抗菌株產幾丁質酶活性測定
研究表明,SMT-24、BHZ-29、SHT-15、SHZ-24菌株數量在0~132 h內出現先快速增加后減小,最后趨于穩定。SMT-24、BHZ-29、SHT-15菌株幾丁質酶活均低于3.00 IU表明該三株菌產幾丁質酶低或不產幾丁質酶,酶活0~132 h內無變化,酶活不受生物量的影響。SHZ-24酶活0~36 h幾丁質酶活呈現指數型增長,36~108 h增長緩慢,108 h達到最大值(12.37±0.15) IU,108~132 h呈現下降趨勢,細菌對數期與酶活呈正相關。圖2

注:A~D分別是菌株SMT-24、BHZ-29、SHT-15、SHZ-24幾丁質酶活變化
Note: A-D are chitinase activity changes of strains SMT-24, BHZ-29, SHT-15 and SHZ-24, respectively
圖2 SMT-24、BHZ-29、SHT-15、SHZ-24拮抗菌株幾丁質酶活變化
Fig.2 Results of chitinase activity change of SMT-24, BHZ-29, SHT-15 and SHZ-24 antagonistic strains
2.4 SMT-24、BHZ-29、SHT-15、SHZ-24拮抗菌株蛋白酶活測定
研究表明,SMT-24、BHZ-29、ShT-15、SHZ-24菌株細菌數量在0~36 h逐漸增加,36 h后細菌數量先下降最終維持穩定。SMT-24、BHZ-29、SHT-15、SHZ-24蛋白酶活在0~12 h迅速增加,隨后緩慢增加,SMT-24、BHZ-29、SHZ-24菌株在84 h蛋白酶活達到最大值分別為(21.21±0.54) IU、(8.65±0.35) IU、(24.22±0.45) IU,SHT-15菌株在108 h蛋白酶活達到最大值(17.78±0.21) IU,84 h后SMT-24、BHZ-29、SHZ-24蛋白酶活緩慢下降,SHT-15酶活達到穩定狀態。蛋白酶活受對數期生長的細菌的影響最大,呈正相關。圖3
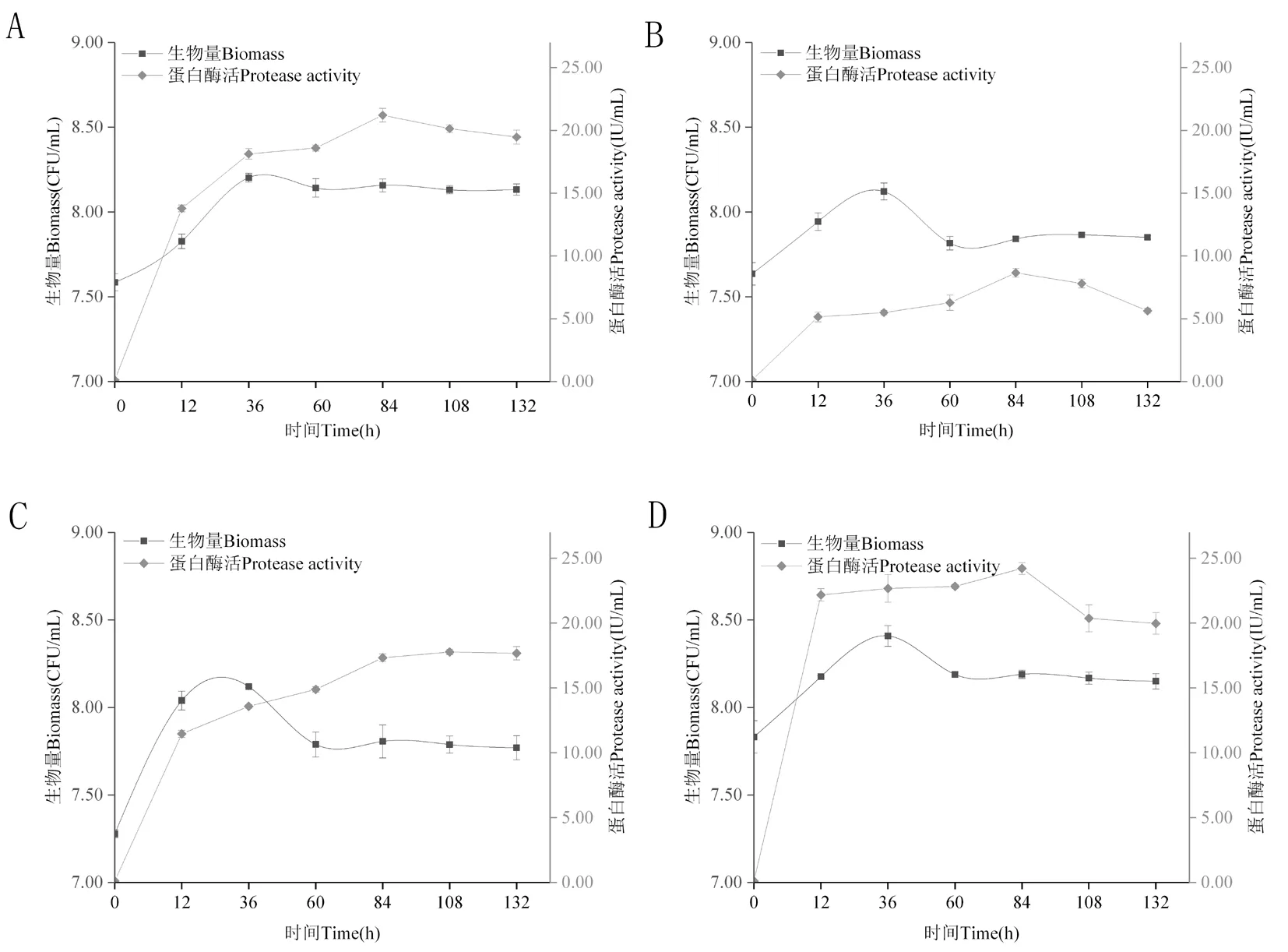
注:A~D分別是菌株SMT-24、BHZ-29、SHT-15、SHZ-24蛋白酶活變化
Note: A-D are protease activity changes of strains SMT-24, BHZ-29, SHT-15 and SHZ-24, respectively
圖3 SMT-24、BHZ-29、SHT-15、SHZ-24拮抗菌株蛋白質酶活變化
Fig.3 Results of protease activity changes of SMT-24, BHZ-29, SHT-15 and SHZ-24 antagonistic strains
2.5 SMT-24、BHZ-29、SMT-15、SHZ-24拮抗菌株葡聚糖酶活測定
研究表明,SMT-24、BHZ-29菌株細菌數量出現先迅速上升后下降,最終維持細菌數穩定,SHT-15、SHZ-24菌株細菌數量先上升后維持穩定。SMT-24、BHZ-29、SHT-15、SHZ-24菌株葡聚糖酶活整體呈現先增加后下降的趨勢,SMT-24、SHT-15、SHZ-24菌株葡聚糖酶活在上升階段增長較快,BHZ-29菌株酶活在上升階段0~12 h增長快速隨后緩慢增加,SMT-24、BHZ-29菌株葡聚糖酶在84 h達到最大值,分別為(12.29±0.23) IU、(9.16±0.46) IU,SHT-15菌株葡聚糖酶活在60 h達到最大值(12.07±0.59) IU,SHZ-24菌株在36 h葡聚糖酶活達到最大值(8.12±0.25) IU。SMT-24、BHZ-29、SHT-15、SHZ-24菌株在0~12 h菌株生物量與酶活呈正相關發展趨勢。圖4

注:A~D分別是菌株SMT-24、BHZ-29、SHT-15、SHZ-24葡聚糖酶活變化
Note: A-D are glucanase activity changes of strains SMT-24, BHZ-29, SHT-15 and SHZ-24, respectively
圖4 SMT-24、BHZ-29、SHT-15、SHZ-24菌株葡聚糖酶活變化
Fig.4 Results of glucanease activity changes of SMT-24, BHZ-29, SHT-15 and SHZ-24 antagonistic strains
2.6 SMT-24、BHZ-29、SHT-15、SHZ-24菌株纖維素酶活性測定
研究表明,BHZ-29、SHT-15、SHZ-24菌株在纖維酶發酵培養基中培養60 h達到最大,生長期較長。SMT-24菌株細菌數量60 h后下降,SMT-24、SHZ-24菌株數量60 h后均維持穩定。SMT-24菌株生長緩慢,0~36 h增長幅度較小,30~108 h先下降后達到穩定,穩定后的細菌數量低于接種量。SMT-24、BHZ-29、SHT-15菌株纖維素酶活在0~84 h為上升階段,84 h達到最大值,分別為(6.43±0.24) IU、(5.80±0.12) IU、(4.81±0.04) IU,84~108 h纖維素酶活下降,SHZ-24菌株纖維素酶活低于3.00 IU,且0~108 h酶活性無變化,SHZ-24菌株不產纖維素酶或者產纖維素酶較低。SMT-24、BHZ-29、SHT-15菌株纖維素酶活受細菌對數期影響較大,呈正相關。SHZ-24菌株纖維素酶活性不受生物量影響。圖5
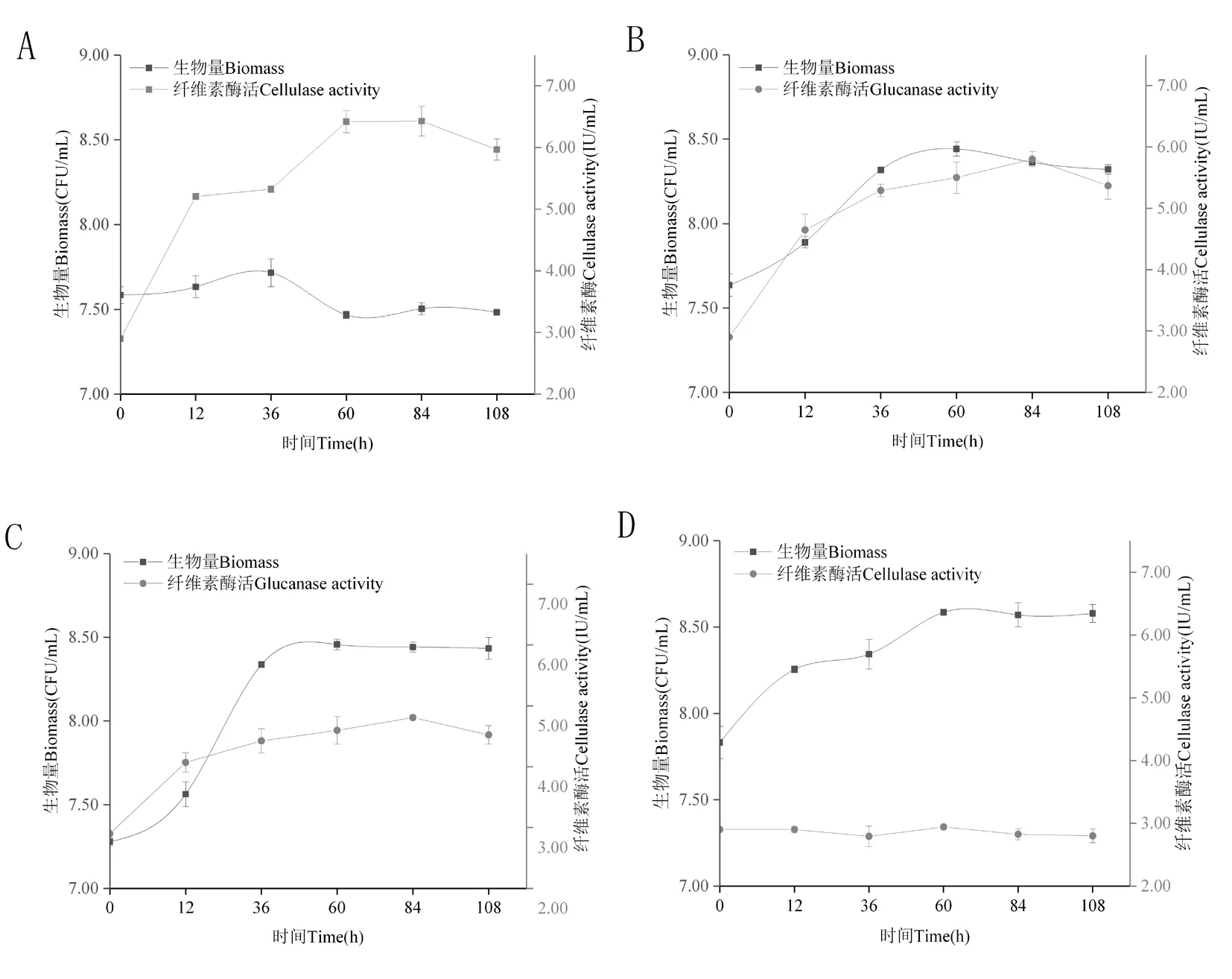
注:A~D分別是菌株SMT-24、BHZ-29、SHT-15、SHZ-24纖維素酶活與生物量變化
Note: A-D are activity cellulase changes of strains SMT-24, BHZ-29, SHT-15 and SHZ-24, respectively
圖5 SMT-24、BHZ-29、SHT-15、SHZ-24菌株纖維素酶活變化
Fig.5 Results of cellulase activity changes of SMT-24, BHZ-29, SHT-15 and SHZ-24 antagonistic strains
2.7 SMT-24、BHZ-29、SHT-15、SHZ-24菌株酶活大小比較
研究表明,SHZ-24菌株幾丁質酶活最大,SMT-24、BHZ-29、SHT-15酶活曲線重合,表明該三株菌幾丁質酶活大小相同。SMT-24、BHZ-29、SHT-15、SHZ-24菌株蛋白酶活大小為:SHZ-24> SMT-24> SHT-15> BHZ-29。葡聚糖酶活整體SMT-24、SHT-15菌株較大,BHZ-29菌株次之,SHZ-24菌株葡聚糖酶活最小。纖維素酶活大小比較為SMT-24> BHZ-29> SHT-15> SHZ-24菌株。圖6
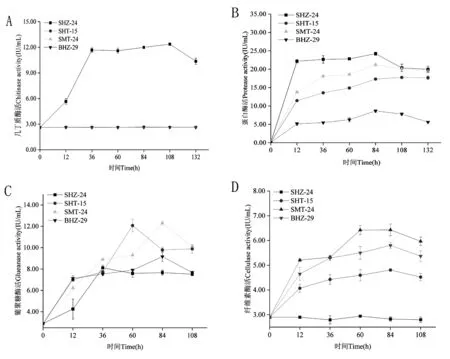
注:A~D分別代表幾丁質酶、蛋白酶、葡聚糖酶、纖維素酶
Note: A-D represent chitinase, protease, glucanase and cellulase, respectively
圖6 SMT-24、BHZ-29、SHT-15、SHZ-24菌株酶活大小比較
Fig.6 Comparing the Enzyme Activity of SMT-24, BHZ-29, SHT-15 and SHZ-24
3 討 論
根據測定與DNS法酶活測定結果,蛋白酶、纖維素酶活、葡聚糖酶活與平板的透明圈外徑大小不一致,表明透明圈外徑大小與液體樣品的酶活之間的正相關關系并不是絕對的,僅以透明圈徑大小作為菌株產酶能力的唯一定量指標并不可靠,這在蔣明星、包衎等[21,22]的研究中也出現類似情況。可能原因是酶活受培養基狀態的影響,細菌在固體平板生長分泌的酶活與在發酵液中生長產酶活性存在差異,另外蛋白酶固體平板中營養成分與液體發酵培養基的成分不同也是造成酶活差異的原因之一。細菌在衰退期時隨著發酵時間延長酶活有不同程度的降低,這可能是隨著發酵時間的延長,代謝物積累,細胞凋亡,發酵液pH降低,從而降低酶活。4株拮抗菌產酶類豐富,蛋白酶在飼料、皮革、食品方面應用廣泛[23],4株拮抗菌均可作為產蛋白酶來源菌株及其他產酶來源菌株。
4 結 論
SMT-24、BHZ-29、SHT-15、SHZ-24菌株均分泌幾丁質酶、蛋白酶、葡聚糖酶、纖維素酶;在發酵期間酶活總體呈現先增加后減小的趨勢,酶活性受細菌對數期影響最大,酶活性與拮抗細菌對數期呈正相關關系(酶活低于3.00 IU除外);4株拮抗菌酶活比較中,SHZ-24菌株產幾丁質酶活最高,酶活最高為(12.37±0.15) IU,SHZ-24菌株產蛋白酶活最高,最高值達(24.22±0.45) IU,SMT-24、SHT-15菌株產葡聚糖酶活較高,最高酶活分別(12.29±0.23) IU、(12.07±0.59) IU,SMT-24菌株產纖維素酶活最高,酶活最高達(6.43±0.24) IU。
