miR-133調控FGFR1蛋白對喉癌細胞遷移和侵襲的影響
2018-12-26 09:03:32潘慶春喻望博李四軍
解放軍醫藥雜志 2018年12期
潘慶春,喻望博,李四軍
喉癌是全球常見的頭頸部惡性腫瘤,而鱗狀細胞癌是最常見的一種病理類型[1-2]。然而,喉癌被診斷時通常處于中晚期階段,并常伴有局部病灶浸潤擴散和遠處淋巴結的轉移[3]。盡管隨著診斷技術的進步和治療方式的改善,總體的生存率有所提升,但仍然不盡如人意[4]。現在學者們更注重對腫瘤分子特征進行區別,并根據特定的基因組差異進行腫瘤的亞型分類[5]。根據精確的腫瘤分類所開發出的針對精確分子改變的臨床治療稱為靶向治療[6-7]。這種治療方法為癌癥患者提供了更為有效的治療,并且與常規化學療法相比,通常具有更小的毒副作用[8]。miRNAs是一類非編碼RNA,能通過與編碼蛋白RNA的3' -UTR端結合,從而影響基因的功能與表達[9]。miRNA通過結合與之相同或相似的互補序列來調節基因表達,引起翻譯的抑制或切割mRNA靶標[10]。miR-133已被證實在多種癌癥的發生發展過程中起到重要作用[11-15],能抑制腫瘤細胞的增殖、遷移、侵襲和細胞周期進展[16]。但miR-133在喉癌發生發展過程中的作用尚不清楚。本研究擬探討miR-133對喉癌細胞的影響及其相關作用機制。
1 材料與方法
1.1組織標本 選取2017年1月—2018年1月我院收治20例喉癌患者的腫瘤組織及癌旁正常組織。患者年齡37~68歲,平均52歲;病例樣本病理類型均為鱗狀細胞癌,臨床病理分期均為Ⅰ~Ⅱ期。將取下的組織樣品在液氮中快速冷凍。所有患者手術前沒有接受過放療或化療。本研究經醫院倫理委員會批準所有實驗組織標本的使用。
1.2細胞培養 人喉癌細胞株Hep-2和TU212購買于武漢大學;RPMI-1640培養基和0.25胰酶(0.25 Trypsin EDTA)購買于美國Gibco公司;胎牛血清和雙抗(Penicillin-Streptomycin Solution)購買于美國海克隆公司;PBS磷酸鹽緩沖液購買于BOSTER。細胞均培養在含10%胎牛血清和1%雙抗的培養基中,所有細胞均在5%CO2,37℃條件下培養,每隔3 d換液1次,待細胞鋪滿培養瓶底進行傳代,以下所有試驗均取處于對數生長期的細胞。
1.3細胞轉染 miR-133 mimics和miR-133 inhibitor購買于上海吉瑪有限公司。PBS緩沖液清洗Hep-2和TU212細胞,應用胰酶消化2 min,轉移至無菌15 ml離心管,離心并計數細胞,以每孔4×105個細胞接種于6孔板中,使過夜細胞融合率達70%左右,細胞分為Hep-2 mimics組、Hep-2 NC組、Hep-2 inhibitor組、TU212 mimics和TU212 NC組,其中mimics組細胞均轉染miR-133 mimics慢病毒,inhibitor組細胞均轉染miR-133 inhibitor慢病毒,NC組細胞均轉染NC慢病毒。無血清培養基將Lipofectamine 2000按3 μl/L稀釋,37℃孵育20 min,用無血清培養基分別將miR-133 mimics和miR-133 inhibitor按50 μmol/L濃度稀釋,常溫下孵育5 min,與Lipofectamine 2000等體積分別混勻,分別加入細胞中,37℃孵育箱繼續培養;12 h后,觀察轉染細胞狀態,并將無血清培養基更換為完全培養基,繼續培養48 h后,提取細胞RNA,驗證轉染效率。
1.4定量實時PCR分析 氯仿、乙醇、異丙醇購買于重慶川東化工公司;Trizol裂解液、Premix ExTaq Version2.0和SYBR Premix Ex Taq購買于日本Takara公司。PCR引物由上海Sangon Biotech公司合成。miR-133的qRT-PCR檢測以U6為內參,采用2-ΔΔCt方法進行統計,重復3次,利用熒光PCR儀檢測cDNA中miR-133的相對表達量,反應條件為:95℃預變性10 min,95℃ 15 s,60℃ 15 s,45個循環,獲取熒光信號溫度為60℃。普通cDNA的qRT-PCR檢測以GAPDH為內參,采用2-ΔΔCt方法進行統計,反應條件為95℃ 10 min,95℃ 15 s,60℃ 1 min,共40個循環。
1.5劃痕愈合實驗 將喉癌細胞以每孔2×104個細胞密度接種于6孔板,融合度達到后,用200 μl消毒槍頭垂直劃線,并于顯微鏡下測量劃痕的初始距離,在恒溫孵箱中孵育72 h后,再次測量劃痕的距離,并計算細胞的遷移率。實驗重復3次。
1.6Transwell侵襲試驗 Transwell基質膠購買于美國BD公司;Transwell小室購買于美國康寧公司。將5×104個細胞置于上室,并加入Matrigel基質膠。將含10%胎牛血清和HGF(20 μg/ml)的培養基置于下室。37℃孵育24 h,去除上膜表面的細胞。95%乙醇固定20 min,0.5%結晶紫溶液染色10 min,自來水沖洗后,在倒置顯微鏡下計數。
1.7熒光素酶報告基因測定 構建含成纖維細胞生長因子受體1(FGFR1)基因野生型3'-UTR(含miR-133識別位點/wt)和突變型3'-UTR(miR-133識別位點點突變/mut)片段,兩端均設計XbaⅠ酶切位點,插入熒光素酶報告基因質粒構建pGL3-3'-UTR wt、pGL3-3'-UTR mut。取生長狀態良好的喉癌Hep-2細胞,質粒轉染后48 h據操作說明,裂解細胞,加入熒光素酶底物,檢測熒光素酶活性,以證明miR-133直接調控PRDX1。
1.8Western blotting實驗 GAPDH小鼠抗人一抗、β-actin小鼠抗人一抗購買于武漢三鷹生物技術有限公司;Western blotting凝膠試劑盒和全蛋白提取試劑盒購買于碧云天公司;ECL發光液購買于索萊寶生物科技有限公司。根據蛋白分子量大小,配制凝膠,根據所測蛋白濃度加入蛋白樣品以及每孔5 μl蛋白marker;跑膠,首先加壓80 V,保證所有樣品在同一水平線,待樣品至分離膠處,將電壓加到120 V;切膠,根據所需條帶大小,對照蛋白marker進行切膠,并準備與所切膠條相同大小濾紙2片和PVDF膜(浸泡甲醇15 s);按照濾紙、膠、PVDF膜、濾紙的順序按序排好,并趕出氣泡,壓緊,在250 mA恒流下,按1 kD蛋白1 min進行轉膜;轉膜結束后,PBST洗膜1次,脫脂奶粉封閉1 h;PBST洗膜1次,將膜浸入稀釋好的一抗中,4℃過夜;PBST洗膜3次,將膜浸入稀釋好的二抗中,常溫下孵育1 h;顯影,PBST洗膜3次后,在暗室中曝光,并分析。
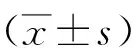
2 結果
2.1miR-133的表達情況 miR-133在喉癌組織和癌旁正常組織的表達分別為0.57±0.12和1.09±0.02。與癌旁正常組織比較,喉癌組織中miR-133呈顯著低表達狀態(P<0.05)。
2.2miR-133對Hep-2和TU212細胞遷移的影響 Hep-2 mimics組和Hep-2 NC組的細胞的遷移率分別為(40.1±10.4)%和(98.4±9.5)%,TU212 mimics組和TU212 NC組的細胞的遷移率分別為(58.6±8.0)%和(99.2±10.1)%。與Hep-2 NC組和TU212 NC組比較,Hep-2 mimics組和TU212 mimics組的細胞遷移率均顯著降低(P<0.05)。見圖1。

圖1 miR-133對人喉癌Hep-2和TU212細胞遷移的影響
2.3miR-133對Hep-2和TU212細胞侵襲的影響 Hep-2 NC組和Hep-2 mimics組細胞侵襲率分別為(98.4±7.5)%和(48.1±10.4)%,TU212 NC組和TU212 mimics組細胞侵襲率分別為(97.2±10.1)%和(38.2±5.3)%。與Hep-2 NC組和TU212 NC組比較,Hep-2 mimics組和TU212 mimics組的細胞侵襲率顯著降低(P<0.05)。見圖2。

圖2 miR-133對人喉癌Hep-2和TU212細胞侵襲的影響
2.4miR-133靶向調控FGFR1 通過Targetscan網站發現miR-133與FGFR1具有相似的結合位點。見圖3。通過雙熒光素酶報告基因驗證二者的關系,結果顯示野生型和突變型FGFR1基因的熒光素酶活性分別為0.26±0.02和0.96±0.06,比較差異有統計學意義(P<0.05)。表明miR-133可以明顯抑制野生型FGFR1基因的熒光素酶活性,而對突變型FGFR1基因的熒光素酶活性無影響。Western blotting結果也表明,Hep-2 mimics組、Hep-2 inhibitor組和Hep-2 NC組FGFR1蛋白表達分別為(31.8±8.0)%、(96.1±9.6)%和(73.2±6.8)%,3組比較差異有統計學意義(P<0.05)。miR-133的過表達和沉默能使Hep-2細胞中的FGFR1蛋白表達量下調和上調。見圖4。

圖3 FGFR1基因與miR-133的3'-UTR基因序列比對FGFR1為纖維細胞生長因子受體1

圖4 miR-133對Hep-2細胞FGFR1蛋白表達的影響FGFR1為纖維細胞生長因子受體1
3 結論
喉癌患者存在miRNAs的失調,有研究顯示,miRNAs在喉癌的增殖、轉移、侵襲和凋亡過程中發揮重要作用[17]。miRNAs在喉癌中可以發揮致癌作用或抑癌作用[18]。Cao等[19]通過研究發現,共有26個miRNAs在喉鱗狀細胞癌組織中高表達,有3個miRNAs顯著低表達,而且還發現,miR-21、miR-93、miR-205和miR-708是顯著上調的,而miR-125b和miR-145顯著下調。本研究結果發現,喉癌組織中miR-133呈顯著低表達狀態。表明miR-133在喉癌中可能是扮演抑癌基因的角色。
不同miRNAs對喉癌細胞的影響可能通過不同的行為方式及相應的機制。如miR-1通過直接影響纖維連接蛋白1的途徑抑制Hep-2細胞的生長、遷移和侵襲[20]。Guo等[21]研究發現,miR-24通過下調S100A8誘導Hep-2細胞形態改變并抑制細胞增殖和侵襲。另外,miR-30b參與調節重要的腫瘤抑制基因p53,在miR-30b過表達的Hep-2細胞中p53表達顯著增強,同時miR-30b過表達增加p53介導的腫瘤細胞凋亡[22]。本研究結果發現,與Hep-2 NC組和TU212 NC組比較,Hep-2 mimics組和TU212 mimics組細胞的侵襲率和遷移率顯著降低。表明miR-133的過表達能顯著抑制Hep-2和TU212細胞的增殖和遷移能力。
成纖維細胞生長因子受體(FGFRs)是一種酪氨酸激酶,位于染色體8p12上,與多種腫瘤侵襲行為密切相關,而且能一定程度上預測患者的生存和預后[23-24]。FGFRs家族包括酪氨酸激酶的亞組受體,由5個結構保守的反式膜糖蛋白組成,含有3個免疫球蛋白樣結構域在其胞外區[25]。被認為是真正的致癌基因,可能在腫瘤發展過程中發揮關鍵作用[26]。FGFRs及其配體,即成纖維細胞生長因子(FGF),參與多種正常生物學進程,包括細胞分化、增殖、遷移和血管生成等[27]。大量證據還表明,FGF-FGFR途徑在腫瘤發展過程中起關鍵作用,并可能促進上皮間質轉化,并激活關鍵的致癌信號通路,包括PI3K/AKT,RAS/MAPK和TGFβ途徑[28]。
FGFR1的高表達也在頭頸部的惡性腫瘤中被發現,而且其上調多見于人乳頭狀瘤病毒陰性鱗癌[29]。與肺癌類似,FGFR1的升高預示著喉癌患者生存率的降低[30]。目前,對喉癌尚無針對FGFR1的靶向治療。本研究通過Targetscan網站發現,miR-133與FGFR1具有相似的結合位點,兩者可能有內在調控關聯。本研究還發現,miR-133可以明顯抑制野生型FGFR1基因的熒光素酶活性,而對突變型FGFR1基因的熒光素酶活性無影響。表明miR-133可以與FGFR1的3'-UTR特異性結合。同時miR-133的過表達和沉默能使Hep-2細胞中的FGFR1蛋白表達量下調和上調。以上結果均證實miR-133能靶向調控FGFR1的表達,從而進一步影響腫瘤細胞惡性生物學進程。
綜上所述,miR-133能通過靶向調控FGFR1蛋白的表達從而影響喉癌細胞遷移和侵襲能力。miR-133可能會成為治療喉癌的新的靶點。
