基于公共基因芯片的多農藥代謝相關的擬南芥UGT基因挖掘
2019-01-24 09:01:14賈曉晨趙小明
西北農業學報 2019年1期
關鍵詞:除草劑
吳 凡,賈曉晨,趙小明,尹 恒
(1.中國科學院大連化學物理研究所,遼寧省碳水化合物研究重點實驗室,遼寧大連 116023;2.中國科學院大學,北京 100049)
農藥的使用已成為防治植物病蟲害、去除雜草、調節農作物生長及提高農產品產量和質量的重要措施。然而,化學農藥的大量使用,常常引發農作物藥害并導致高超標的農藥殘留,直接危害人類健康和生存環境。因此,尋找有效、經濟、環保的農藥降解方法對實現農作物高效、安全生產和發展可持續農業具有重要意義。
尿苷二磷酸糖基轉移酶(UGT)是植物4相代謝解毒過程中第2階段的關鍵家族之一,能夠糖基化修飾多種官能團,降低或去除外源物質毒性,對多種外源毒物均有代謝能力,例如農藥、微生物源的植物毒素、環境污染物等[1-2]。研究表明擬南芥中大約40%的UGT能夠參與異源物質的結合反應,主要歸屬于D、E、H和L類群,并且體外試驗證明其中的13個UGT基因能夠同時對羥基、巰基、氨基表現出活性[3-4],UGT基因的廣譜結合能力得到證明。UGT參與農藥代謝的功能已有相關報道,例如:體外試驗證明擬南芥的UGT72B1對2種污染物3,4-二氯苯胺(DCA)和2,4,5-三氯苯酚(TCP)具有高度的結合活性,敲除UGT72B1后對TCP和DCA的結合活性顯著下降[5]。進一步研究證實UGT72B1具有O-糖基化和N-糖基化的雙功能催化活性[6]。擬南芥中的UGT72E1、UGT75B1、UGT75D1、UGT84A1、UGT84A2、UGT84B1經體外試驗證明均能代謝外源TCP[7]。并且,Edwards等[8]通過體外試驗證明糖基轉移酶(Glucosyltransferase,GT)的表達還會受到除草劑安全劑的誘導,在小麥、玉米和擬南芥中,使用多種除草劑安全劑后,GT對TCP和DCA的活性相對于對照組均增強了10倍。盡管植物來源的UGT基因的殺蟲劑代謝功能未見報道,但小菜蛾中的UGT2B17能夠提供對氯蟲苯甲酰胺的抗性,該基因被RNA沉默后,毒性作用顯著增強[9]。因此,UGT不僅參與農藥代謝,并且具備廣譜的底物結合能力,但對具備廣譜農藥代謝能力的UGT基因鮮有研究。
目前,關于UGT家族在農藥代謝中的應用研究依然較少,而且由于農業環境的復雜性,往往會同時存在多種農藥殘留,因此研究具備廣譜糖基化功能的UGT家族具有重要意義。此外,UGT作為自然界中最大的功能酶家族之一,其數量龐大,難以通過生物化學試驗快速有效地鑒定出具有這種特性的UGT基因。近年來發展起來的各種高通量測序手段如Microarray和RNA-sequence,均能快速反映出試驗對象對處理條件的響應,提供了一個強有力的手段來分析預測基因的生物學功能。
因此,本研究利用NCBI中的基因表達綜合數據庫(GEO)公共數據庫進行擬南芥基因芯片的篩選,采用差異表達分析、聚類分析及共表達分析等多種生物信息學的分析手段對擬南芥中的UGT基因進行研究,為探索植物中具備廣譜糖基化農藥的UGT基因提供參考。
1 材料與方法
1.1 試驗材料
在NCBI的GEO數據庫中篩選并收集關于農藥與除草劑安全劑的66個樣本數據,所有樣本都基于GPL198平臺。芯片信息經處理后,其對比試驗注釋如表1所示。
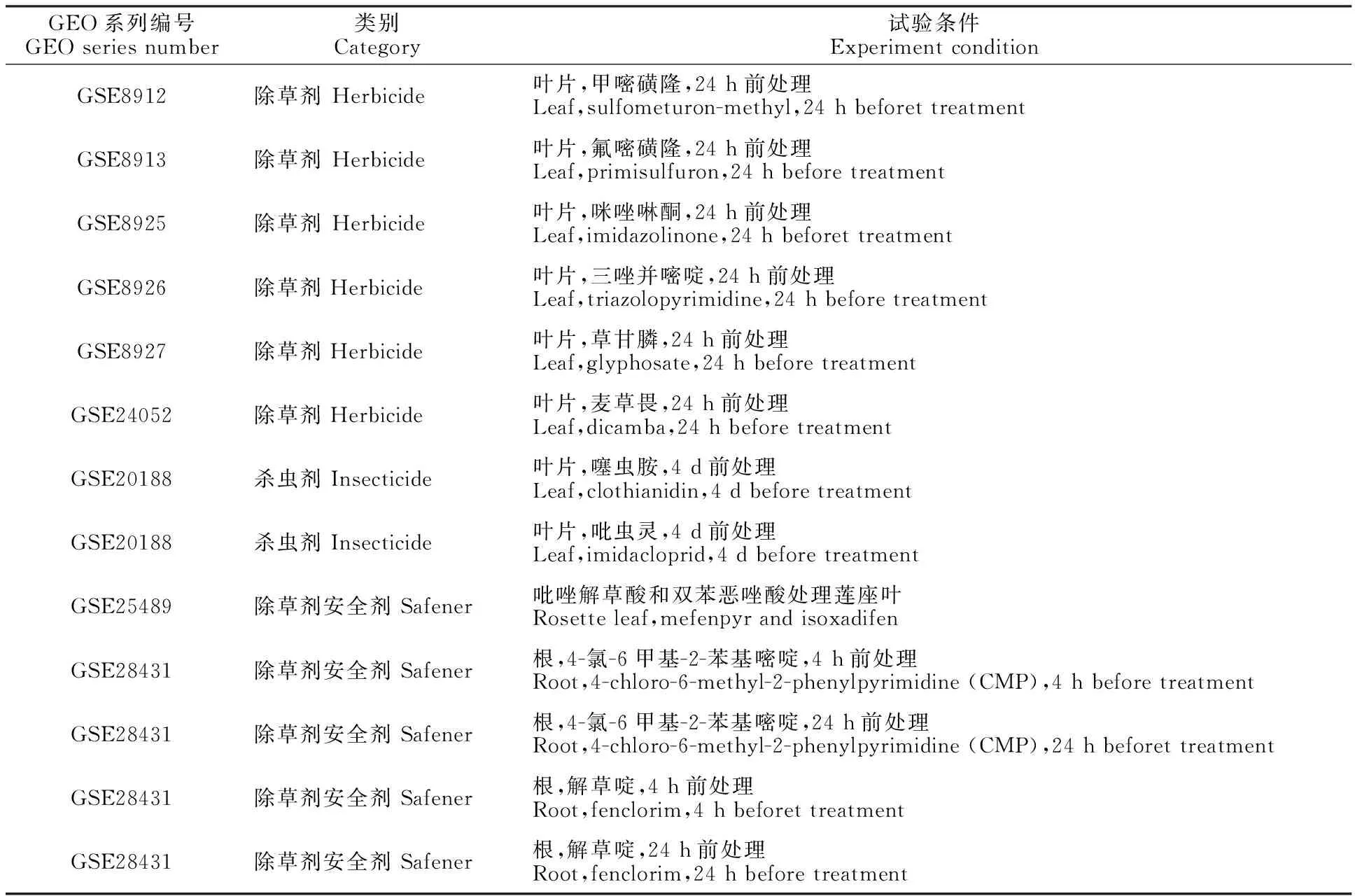
表1 對比試驗注釋表Table 1 Annotation of comparative experiments
1.2 差異表達分析
編寫一份R語言腳本使用Bioconductor limma函數包中的Empirical Bayes方法進行基因的差異表達計算,選擇以| log2(Fold change)(簡稱logFC)|>=1,作為顯著差異閾值。
1.3 熱圖分析
本研究使用R語言中的pheatmap函數包制作熱圖。對UGT基因聚類距離的計算采用歐氏距離。
1.4 相關性
使用皮爾森相關系數(Pearson correlation coefficient,PCC)計算基因間的相關系數,設定顯著性閾值為0.8。為排除不同試驗的背景差異,使用logFC向量計算基因間的相關系數。
1.5 GO分析
使用DAVID (https://david.ncifcrf.gov/home.jsp)進行在線GO分析,富集的顯著性閾值為P<0.01。
1.6 構建共表達網絡
編寫一份R語言腳本獲取UGT基因的相關基因,構成共表達網絡的結點(基因)集合。計算所有UGT基因的相關基因間的相關性,設置PCC閾值為0.8以產生基因關聯(邊),使用Cytoscape[10]構建共表達網絡。
2 結果與分析
2.1 擬南芥UGT的表達譜分析
首先,依照注釋信息對擬南芥106條UGT基因的表達譜進行分析,其結果如圖1所示。結果表明,UGT家族受到農藥誘導后發生強烈的轉錄重編程(圖1-A),暗示UGT家族在農藥代謝中的重要作用。聚類結果顯示,UGT73C1、UGT73B4、UGT73B5、UGT73C6、UGT73B2、UGT76B1、UGT87A2和UGT74E2在多種農藥處理中均高度上調,其平均上調倍數如表2所示,其中UGT74E2的轉錄上調高達26.25倍(表2)。Edwards等[8]對擬南芥UGT家族的底物選擇性研究發現,UGT73C1、UGT73B1、UGT73B2、UGT73B4和UGT73B5能夠糖基化含有羥基或巰基的一種或多種底物,佐證了本研究中候選基因的廣譜結合能力。在收集的芯片數據中,所采用農藥的結構存在明顯差異,包含多種末端基團,其結構更為復雜,暗示這8個UGT基因具有廣泛的底物選擇性,具有更重要的研究價值,是多農藥代謝相關的關鍵候選UGT基因。
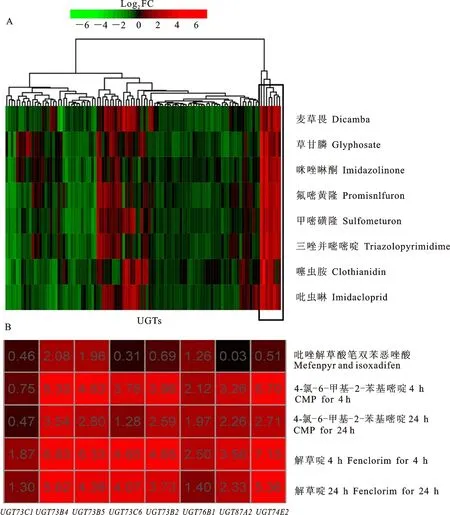
A.擬南芥106條UGT基因在8種農藥處理前后的轉錄水平變化,方框內標記的為8個候選UGT基因 Transcriptional fold-change of 106 UGTs in 8 kinds of pesticides,respectively, UGTs in the box are candidates to metabolize multiple pesticides; B. 8個候選UGT基因在5種不同的安全劑處理條件下的轉錄水平變化,每一個單元格內的數字代表該基因的logFC值 Transcriptional fold-change of 8 key UGTs in 5 kinds of safener treatments,the number in each cell represents logFC value
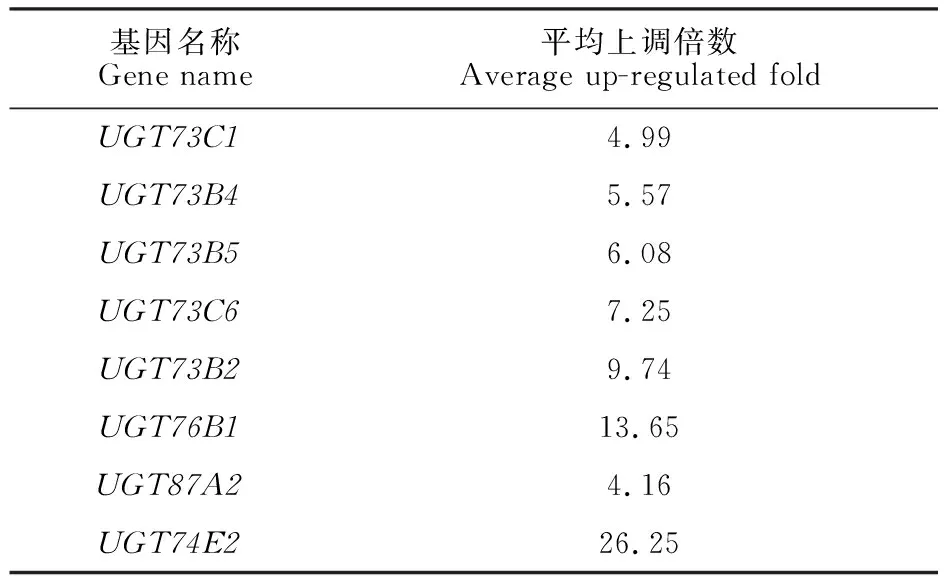
圖1 擬南芥UGT的表達譜分析Fig.1 Analysis of UGT expression profiles in Arabidopsis thaliana
除草劑安全劑可增強作物對除草劑的代謝能力,是目前最受關注和普遍認同的一種機制,并且一般認為僅改變除草劑的代謝速率,而不改變其代謝途徑,其中受到誘導的基因也包括糖基轉移酶[11-12]。因此,筆者收集了NCBI中有關除草劑安全劑的基因芯片數據進行表達譜分析,其結果如圖1-B所示。候選UGT基因在不同的除草劑安全劑中,轉錄水平均受到強烈的誘導上調,這進一步證明了候選UGT基因參與了解毒過程。
2.2 候選UGT與植物解毒過程的相關性
共表達基因是指在多種外界條件下,這些基因的表達均表現出高度的相關性,具有更大的可能性參與相同的生物學途徑或生物學功能[13]。因此,通過UGT基因的共表達基因功能可以對UGT的功能進行探索。通過計算候選UGT基因與全基因組基因的相關性,篩選得到772個高度相關的基因,對這個基因集合進行GO富集分析。結果如圖2所示,在生物學過程分類中,共表達基因主要富集于谷胱甘肽代謝過程、毒性催化過程、響應毒性底物、響應鎘離子等;在細胞組分分類中,共表達基因主要存在于胞漿,蛋白酶體復合物,過氧化物酶體,液泡膜中;按分子功能分類,這些基因主要具有谷胱甘肽轉移酶活性和反向轉運體活性。植物解毒過程通常是經過活化反應(CYP450家族等)、結合反應(UGT家族、GST家族等)和區室化反應(ABC轉運蛋白家族)共同完成,最終異源毒物交聯物會在液泡中等待進一步的代謝反應。共表達基因富集的分子功能、生物學過程以及細胞定位均與植物的解毒過程高度重疊,表明候選UGT與其他解毒酶系構成完整的4相代謝解毒過程,以完成對農藥的解毒作用。
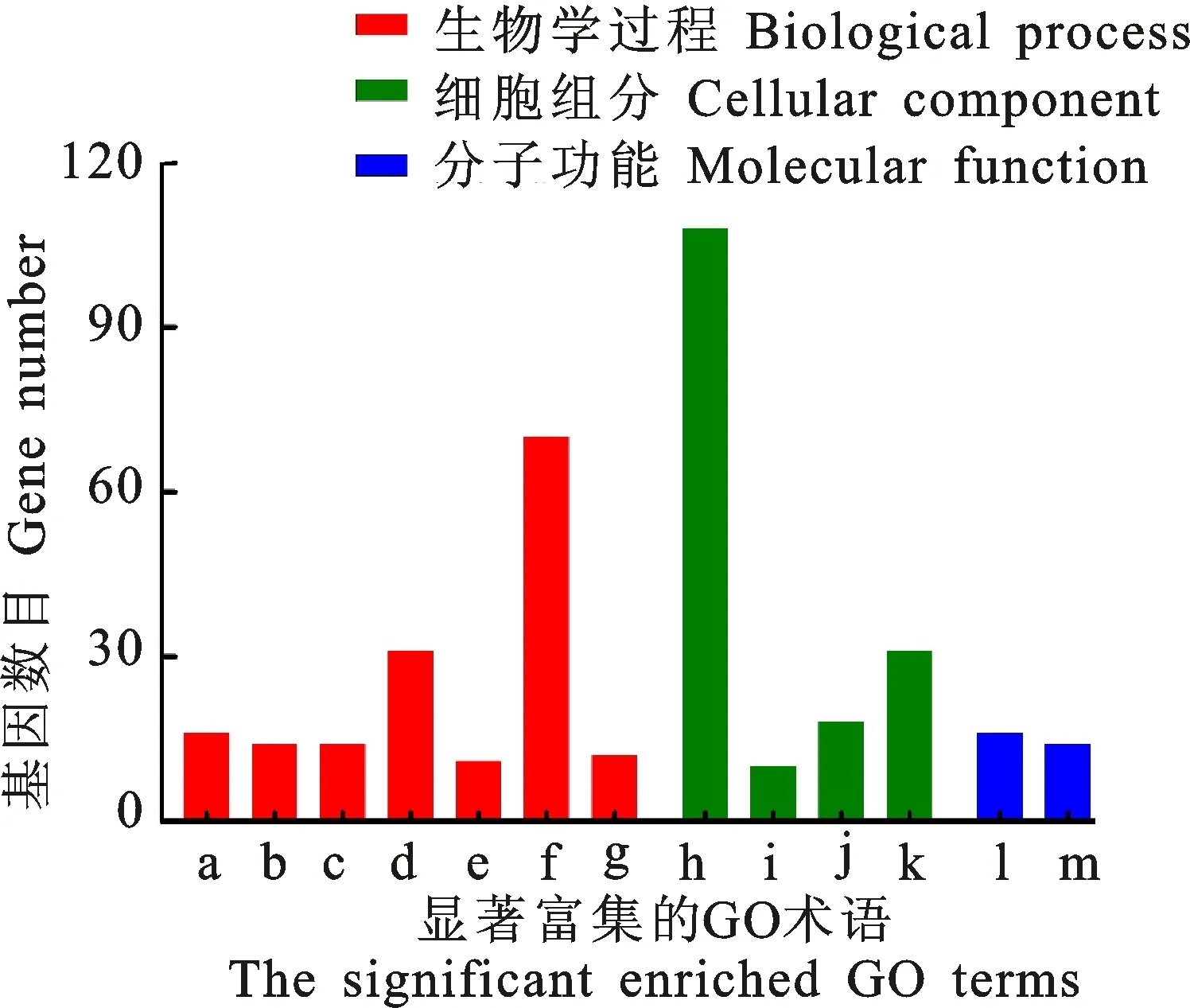
a.谷胱甘肽代謝過程 Glutathione metabolic process;b.毒素分解過程 Toxin catabolic process;c.響應毒性底物 Response to toxic substance;d.響應鎘離子 Response to cadmium ion;e.三羧酸循環 Tricarboxylic acid cycle;f.氧化還原過程 Oxidation-reduction process;g.藥物跨膜運輸 Drug transmembrane transport;h.細胞質 Cytosol;i.蛋白酶體復合物 Proteasome complex;j.過氧化物酶體 Peroxisome;k.液泡膜 Vacuolar membrane;l.谷胱甘肽轉移酶活性 Flutathione transferase activity;m.逆向轉運活性 Antiporter activity
圖2擬南芥候選UGT共表達基因的GO富集分析
Fig.2GOenrichmentanalysisofco-expressiongenesof8keyUGT
2.3 共表達網絡揭示解毒過程的關鍵基因
為了進一步探索UGT基因與其共表達基因之間的聯系,筆者將候選UGT基因及共表達基因的相關關系以網絡圖的形式呈現(圖3)。網絡圖中共包含780個結點(基因),PCC大于0.8的結點間會形成一條邊。在網絡圖中,對植物4相代謝過程中重要的功能酶家族進行標記。網絡中包含的解毒第1階段代謝酶系主要有2OG加氧酶超家族、細胞色素P450超家族及alpha/beta水解酶超家族等;第2階段代謝相關酶系包括除候選基因以外的UGT超家族、谷胱甘肽轉移酶超家族及SAM甲基轉移酶超家族等;第3階段相關酶系主要是ABC轉運蛋白超家族。目前,4相代謝前3個階段中的關鍵酶系已經得到大量的研究,但第4個階段涉及到異源物質偶聯物在液泡或胞質的進一步代謝,還不具備代表酶系[14-15],因此未予標記。共表達網絡顯示,8個候選UGT基因之間是直接相連或間接相連,證明了這些UGT基因之間也具備高度的相關性。
橋接多個候選UGT的基因同樣具有重要作用,部分基因的解毒功能已經明確。例如,GSTU24在網絡圖中與4個候選UGT基因相關,能夠催化TNT脫硝并且形成谷胱甘肽交聯物[16]。PMAT1基因在網絡中同時與5個候選UGT基因相關,編碼丙二酰轉移酶,盡管一般認為丙二酰轉移酶系在植物解毒第2個階段發揮作用,但目前有研究表明煙草PMAT1能夠丙二酰化多種酚類糖苷,并且其表達水平與酚類糖苷的體外排出與液泡儲存直接相關[17],暗示PMAT1參與了異源物質的代謝解毒的第4階段。ABCC4與3個候選UGT相關,雖然沒有直接的證據表明擬南芥來源的ABCC4能夠參與農藥代謝,但是斑馬魚來源的ABCC4基因有助于多種農藥的排出[18-19]。這些橋接節點不但本身具備重要的解毒功能,并且還進一步的暗示候選UGT基因參與植物的農藥代謝過程。此外,在共表達網絡中,UGT73B4和UGT73B5的共表達基因集合中含有最豐富的植物解毒相關酶,暗示了這2個UGT基因的底物更為廣泛,是參與廣譜農藥代謝的關鍵UGT基因。
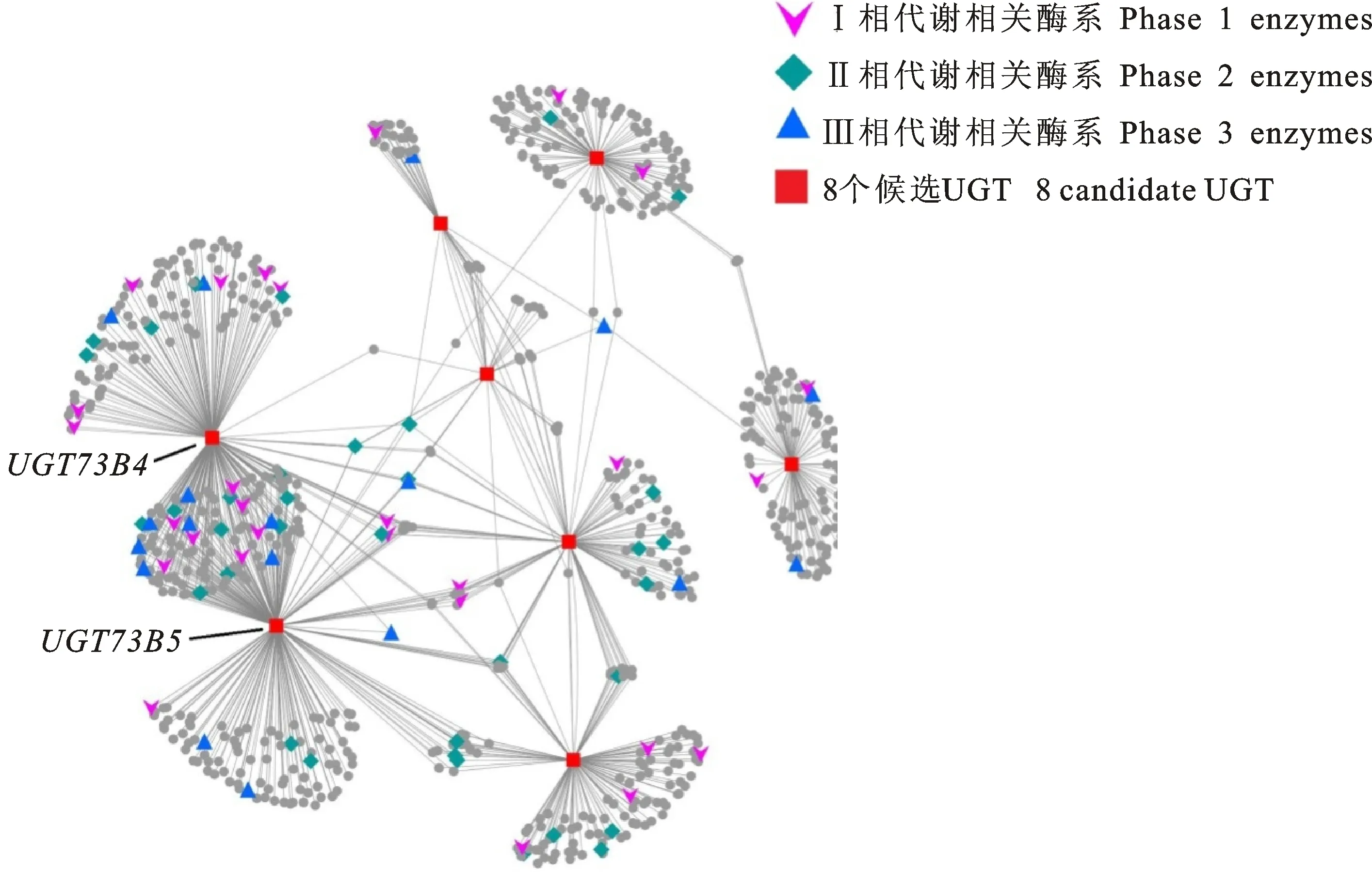
圖3 由8個候選的UGT及其相關基因構建的共表達網絡Fig.3 Co-expression network of 8 key UGTs and their co-expression genes
3 討 論
農藥殘留問題直接威脅到人類健康與生存環境,尋找一個綠色環保的解決方案是目前研究的熱點。植物在對抗異源物質的過程中,進化出了一套復雜的解毒系統,能夠代謝大量的外源物質,包括微生物源的植物毒素、異種化合物、污染物以及農藥等[20]。如何利用植物自身的解毒功能完成農殘降解或者開發新型安全劑等具有廣泛的應用前景。同時,在植物的解毒過程中,最廣泛存在的異源物質結合過程是利用UDP-葡萄糖作為供體形成結合物,該過程的代表是UGT家族。因此筆者以此為出發點,試圖找出具備廣譜農藥代謝能力的UGT基因。
本研究收集公共基因芯片數據,通過表達譜分析在擬南芥的UGT家族中篩選出8個候選UGT基因,可能具備廣譜農藥代謝能力。對這些候選UGT基因的共表達基因進行GO富集分析,結果表明這些基因參與的生物學途徑最富集于谷胱甘肽代謝過程、催化和響應毒性物質中,分子功能則主要具有谷胱甘肽轉移酶(GST)活性。谷胱甘肽中半胱氨酸上的巰基為其活性基團,能夠在谷胱甘肽轉移酶的催化下與毒性物質交聯,從而轉化成無毒物質,在許多異源物質的脫毒中起到至關重要的作用[6]。GST在農作物針對除草劑代謝中有廣泛的報道,Milligan等[21]將玉米GST27導入小麥中,表現出對甲草胺、二甲酚草胺和撲草滅的抗性。水稻的GSTU3對酰胺類除草劑具有顯著的脫毒作用[22]。GO富集分析的結果顯示,候選UGT基因與GST等重要的解毒酶系共同參與解毒過程,暗示了這些UGT基因的農藥解毒活性。進而,共表達網絡不但顯示UGT73B4和UGT73B5是廣譜農藥代謝的關鍵基因,還揭示了一批與植物解毒相關的重要基因。在網絡中,與多個候選UGT相關的基因通常也具備農藥代謝能力,例如,有研究推測沿煙草PMAT1可能涉及到異源物質的代謝調控[17],共表達網絡為PMAT1基因參與農藥代謝提供了新的依據,其與5個關鍵的UGT基因存在高度相關性。
本研究利用公共數據庫的基因芯片數據,對擬南芥糖基轉移酶1家族進行了農藥代謝相關的整體生物信息學分析與驗證,為解決農業上的農藥殘留問題提供了參考,為新的農藥解毒劑或安全劑研發提供依據,為擬南芥UGT基因下一步的生理學和毒理學研究奠定基礎。
猜你喜歡
世界農藥(2019年3期)2019-09-10 07:04:10
今日農業(2019年15期)2019-01-03 12:11:33
現代園藝(2017年19期)2018-01-19 02:50:21
長江蔬菜(2016年10期)2016-12-01 03:05:27
獸醫導刊(2016年12期)2016-05-17 03:51:29
現代農業(2016年5期)2016-02-28 18:42:36
雜草學報(2015年2期)2016-01-04 14:58:05
種業導刊(2016年9期)2016-01-03 01:27:14
營銷界(2015年23期)2015-02-28 22:06:18
營銷界(2015年22期)2015-02-28 22:05:11
