紅芽芋超低溫療法脫毒苗基因組DNA 甲基化的MSAP分析
2019-01-24 09:01:14尹明華萬(wàn)志庭
西北農(nóng)業(yè)學(xué)報(bào) 2019年1期
關(guān)鍵詞:植物
尹明華,廖 玉,萬(wàn)志庭
(上饒師范學(xué)院 生命科學(xué)學(xué)院,江西上饒 334001)
表觀遺傳學(xué)(Epigenetics)是指在DNA序列沒(méi)有發(fā)生變異的情況下基因表達(dá)的可遺傳改變,且可隨細(xì)胞有絲分裂和(或)減數(shù)分裂遺傳給后代[1]。DNA 甲基化是一種表觀遺傳現(xiàn)象,即在 DNA 甲基轉(zhuǎn)移酶的催化下,以 S-腺苷甲硫氨酸為甲基供體,將甲基轉(zhuǎn)移到 DNA分子中特定堿基上的過(guò)程,主要形成5-甲基胞嘧啶[2]。DNA甲基化通過(guò)改變?nèi)旧|(zhì)結(jié)構(gòu)、DNA 構(gòu)象、DNA 穩(wěn)定性及DNA 與蛋白質(zhì)相互作用方式,關(guān)閉某些基因的活性從而控制基因表達(dá)[3]。DNA甲基化參與植物的生長(zhǎng)發(fā)育和組織分化[4]。DNA甲基化作為細(xì)胞記憶的一種機(jī)制,是表觀遺傳學(xué)研究的“主角”[5]。
紅芽芋(ClocasiaescalentaSchott),單子葉植物,屬天南星科,為多年生宿根性草本植物。江西鉛山素有種植紅芽芋的傳統(tǒng),種植歷史悠久,藥食兼優(yōu)。食用,肉質(zhì)松,品質(zhì)鮮美、細(xì)膩,粉而不粘,口感好;藥用,增強(qiáng)人體的免疫功能,潤(rùn)腸通便,防止便秘[6]。長(zhǎng)期以來(lái),紅芽芋多采用無(wú)性繁殖方法,不僅繁殖系數(shù)低,而且易積累病毒,如芋花葉病毒、黃瓜花葉病毒和芋羽狀斑駁病毒等,導(dǎo)致種性退化,品質(zhì)變差,產(chǎn)量下降[7]。超低溫療法(Cryotherapy)是近年發(fā)展起來(lái)的基于超低溫保存(Cryopreservation)的一種新型高效的脫毒技術(shù),相比傳統(tǒng)脫毒法具有脫毒率高和脫毒率不依賴莖尖大小等兩個(gè)顯著優(yōu)點(diǎn)[8]。現(xiàn)有研究表明,超低溫保存實(shí)質(zhì)是一種逆境脅迫的過(guò)程,在這個(gè)過(guò)程中,存在多種滲透脅迫,如PEG、甘露醇等,均能引發(fā)基因組 DNA 特異區(qū)段的甲基化[9]。但病毒是一種生物脅迫,用超低溫保存的技術(shù)脫除病毒實(shí)際上是一種解除生物脅迫的過(guò)程。
有研究[10]表明,植物感染病毒導(dǎo)致的甲基化整體水平上升有利于病毒攻擊時(shí)植物基因組的穩(wěn)定。但超低溫療法脫毒后,再生脫毒苗的基因組DNA甲基化有何變化,尚無(wú)相關(guān)報(bào)道。檢測(cè)DNA甲基化的方法主要有色譜法、酶切法、測(cè)序法、甲基化PCR檢測(cè)法、芯片法等多種方法,MSAP(Methylation sensitive amplification polymorphism,甲基化敏感擴(kuò)增多態(tài)性)技術(shù)是一種以甲基化修飾敏感性不同限制性內(nèi)切酶和PCR為基礎(chǔ)的新技術(shù),靈敏度高,不受被測(cè)DNA序列信息限制,檢測(cè)位點(diǎn)多,成本低,操作簡(jiǎn)單,重復(fù)性高,已成為檢測(cè)植物基因組DNA甲基化水平和模式的重要方法[11]。本試驗(yàn)以紅芽芋超低溫療法脫毒苗為研究對(duì)象,使用MSAP方法并結(jié)合毛細(xì)管自動(dòng)電泳儀對(duì)其基因組DNA甲基化水平和甲基化模式進(jìn)行分析,為超低溫療法脫毒的表觀遺傳研究提供基礎(chǔ)資料。
1 材料與方法
1.1 材 料
江西鉛山紅芽芋帶毒苗(對(duì)照組,編號(hào)為1~10);江西鉛山紅芽芋超低溫療法脫毒苗(處理組,編號(hào)為11~30)。以上30份材料均由上饒師范學(xué)院生命科學(xué)學(xué)院植物組織培養(yǎng)室提供。
1.2 方 法
1.2.1 DNA提取 采取改良的CTAB法進(jìn)行DNA提取,每個(gè)樣本采取0.5 g葉片進(jìn)行DNA提取。
采取改良的CTAB法進(jìn)行DNA提取,每個(gè)樣本采取0.5 g葉片進(jìn)行DNA提取。步驟如下:
①在65 ℃水浴鍋中預(yù)熱CTAB提取液。
②在液氮中迅速研磨樣品,將粉末狀材料轉(zhuǎn)入2 mL離心管中,加入預(yù)熱的CTAB提取液(每克樣品加入3~5 mL的提取液),65 ℃保溫30~60 min,每隔10 min輕輕顛倒混勻。
③11 000 r/min,離心5 min,取上清轉(zhuǎn)入新的離心管中。
④加入等體積酚/氯仿,充分混勻,11 000 r/min,離心10 min,取上清轉(zhuǎn)入新離心管。
⑤加入等體積氯仿,充分混勻,11 000 r/min,離心10 min,取上清轉(zhuǎn)入新離心管。
⑥重復(fù)步驟④和⑤。
⑦加入2/3體積的異丙醇混勻,室溫放置,沉淀15 min。
⑧11 000 r/min,離心6 min,棄上清。
⑨將沉淀用φ=70%的乙醇漂洗1次,室溫條件下11 000 r/min離心2 min,棄上清,重復(fù)洗1次。
⑩往沉淀中加入50 μL 1×TE溶液,先混勻,再靜置30 min,中間過(guò)程顛倒混勻1~2次。
?提取產(chǎn)物取2~3 μL,用20 g/L瓊脂糖凝膠電泳檢測(cè),其余置于-20 ℃保存,備用。
1.2.2 酶切 用識(shí)別四堿基的HpaⅡ和MspⅠ同裂酶(NEB) 分別與識(shí)別六堿基的核酸內(nèi)切酶EcoRⅠ(NEB) 組合對(duì)樣本DNA進(jìn)行雙酶切,DNA 酶切反應(yīng)體系為20 μL。第1個(gè)反應(yīng)中,400 ng 樣本DNA,2 μL 10×Buffer 4(NEB)[50 mmol/L KAc、20 mmol/L Tris-acetate、10 mmol/L MgAc2、1 mmol/L dTT(pH 7.9,25 ℃)],EcoRⅠ和HpaⅡ內(nèi)切酶各0.8 μL,37 ℃保溫過(guò)夜。第2個(gè)反應(yīng)中,400 ng 樣本DNA,2 μL 10×Buffer1(NEB) [10 mmol/L Tris-HCl、10 mmol/L MgCl2、1 mmol/L dTT(pH 7.0,25 ℃)],EcoRⅠ和MspⅠ內(nèi)切酶各0.8 μL,37 ℃保溫過(guò)夜。
1.2.3 連 接 在酶切片段的兩端加上人工設(shè)計(jì)的與EcoRⅠ和HpaⅡ/MspⅠ酶切位點(diǎn)互補(bǔ)的人工接頭(表1)。接頭連接體系為20 μL:2 μL 10×T4Buffer、0.4 μL 20 μmol/LEcoRⅠ和HpaⅡ/MspⅠ接頭,其余用ddH2O補(bǔ)齊,16 ℃過(guò)夜。
1.2.4 預(yù)擴(kuò)增 預(yù)擴(kuò)增反應(yīng)體系為20 μL,其中含有0.4 μL 10 mmol/L dNTPs、2 μL 10×Buffer、0.2 μL 5 U/ μLTaq酶、0.5 μL 10 μmmol/L E00-primer、0.5 μL 10 μmmol/L M00-primer,其余用ddH2O補(bǔ)齊。反應(yīng)條件為:94 ℃ 30 s,56 ℃ 1 min,72 ℃ 1 min,26 個(gè)循環(huán),72 ℃延伸10 min。預(yù)擴(kuò)增產(chǎn)物稀釋20倍,供選擇擴(kuò)增用。
1.2.5 選擇性擴(kuò)增 選擇擴(kuò)增體系同預(yù)擴(kuò)增體系。條件為:94 ℃預(yù)變性5 min,94 ℃ 30 s,65 ℃至56 ℃ 30 s,72 ℃ 1 min,13個(gè)循環(huán),退火溫度每個(gè)循環(huán)降0.7 ℃進(jìn)行降式PCR擴(kuò)增;94 ℃ 30 s,56 ℃ 30 s,72 ℃ 1 min,23 個(gè)循環(huán),72 ℃延伸10 min。
1.2.6 毛細(xì)管電泳 將甲酰胺與分子質(zhì)量?jī)?nèi)標(biāo)按100∶1的體積比混勻后,取9 μL加入上樣板中,再加入1 μL稀釋10倍的PCR產(chǎn)物。然后使用3730XL測(cè)序儀進(jìn)行毛細(xì)管電泳。電泳結(jié)束后,利用GeneMarker 2.2 軟件對(duì)得到的原始數(shù)據(jù)進(jìn)行分析,將各泳道內(nèi)分子量?jī)?nèi)標(biāo)的位置與各樣品峰值的位置做比較分析,得到片段大小。再根據(jù)無(wú)帶和有帶情況轉(zhuǎn)化為0/1數(shù)據(jù)矩陣。最后進(jìn)行甲基化率和甲基化模式比較分析。
2 結(jié)果與分析
2.1 DNA提取結(jié)果
瓊脂糖凝膠電泳結(jié)果顯示,所提取樣本基因組 DNA主帶清晰,無(wú)降解現(xiàn)象(圖1)。再用紫外分光光度計(jì)測(cè)定,OD260/OD280比值為1.7~1.9,DNA的純度較高,極少含有蛋白質(zhì),完全能滿足試驗(yàn)對(duì)DNA的要求。
2.2 預(yù)擴(kuò)增結(jié)果
預(yù)擴(kuò)增產(chǎn)物通過(guò)20 g/L瓊脂糖凝膠電泳檢測(cè),結(jié)果表明,預(yù)擴(kuò)產(chǎn)物均勻彌散,且連續(xù)成片; 彌散帶只在100~1 000 bp低分子質(zhì)量區(qū)域出現(xiàn),而在高分子質(zhì)量區(qū)域沒(méi)有出現(xiàn),表明DNA消化完全,符合MSAP選擴(kuò)模板的要求。

表1 試驗(yàn)所用接頭和引物序列信息Table 1 Sequence information on adaptors and primers in this test
2.3 基因組DNA甲基化水平分析
依據(jù)HpaⅡ和MspⅠ對(duì)甲基化敏感程度不同,MSAP片段可分為4種類型:第Ⅰ類型EcoRⅠ/HpaⅡ和EcoRⅠ/MspⅠ中均有條帶,甲基化的位點(diǎn)無(wú)改變;第Ⅱ類型EcoRⅠ/HpaⅡ有條帶而EcoRⅠ/MspⅠ中無(wú)條帶(僅有1條鏈甲基化的半甲基化位點(diǎn));第Ⅲ類型EcoRⅠ/HpaⅡ無(wú)條帶(兩條鏈均甲基化)而EcoRⅠ/MspⅠ中有條帶;第Ⅳ類型EcoRⅠ/HpaⅡ和EcoRⅠ/MspⅠ中均無(wú)條帶[12]。本試驗(yàn)采用10對(duì)選擇性擴(kuò)增引物組合(E32MSP40、E32MSP48、E35MSP48、E36MSP43、E37MSP48、E39MSP44、E40MSP44、E50MSP43、E78MSP40、E86MSP48)對(duì)雙酶切后的30個(gè)樣本進(jìn)行擴(kuò)增(圖2)。紅芽芋超低溫療法脫毒苗基因組DNA甲基化水平分析見(jiàn)表2。從表2可知,對(duì)照組總擴(kuò)增帶數(shù)522條,甲基化總帶數(shù)297條,總甲基化率57.06%,全甲基化帶數(shù)194條,全甲基化率37.22%,半甲基化帶數(shù)103條,半甲基化率19.84%;處理組總擴(kuò)增帶數(shù)504條,甲基化總帶數(shù)339條,總甲基化率67.33%,全甲基化帶數(shù)253條,全甲基化率50.2%,半甲基化帶數(shù)86條,半甲基化率17.13%。表明超低溫療法脫毒后,總甲基化率提高10.27%,全甲基化率提高12.98%,但半甲基化率下降2.71%。

編號(hào)1~30和M分別表示樣本1~30和Marker No. 1-30 and M represent samples 1-30 and marker,respectively
2.4 基因組DNA甲基化模式分析
紅芽芋經(jīng)過(guò)超低溫療法脫毒后,其基因組DNA甲基化模式有3種:第1種模式為甲基化條帶數(shù)無(wú)變化,可分為A1(非甲基化→非甲基化)、A2(半甲基化→半甲基化)、A3(全甲基化→全甲基化)3種類型,其比例為29.94%。第2種模式為去甲基化模式,可分為B1(半甲基化→非甲基化)、B2(全甲基化→非甲基化)、B3(超甲基化→非甲基化)、B4(超甲基化→半甲基化)、B5(超甲基化→全甲基化)5種類型,其比例為48.53%,表明紅芽芋經(jīng)過(guò)超低溫療法脫毒后在去甲基化模式中主要由甲基化位點(diǎn)變?yōu)榉羌谆稽c(diǎn)。第3種模式為甲基化模式,可分為C1(非甲基化→半甲基化)、C2(非甲基化→全甲基化)、C3(非甲基化→超甲基化)、C4(半甲基化→超甲基化)、C5(全甲基化→超甲基化),其比例為18.20%。表明紅芽芋經(jīng)過(guò)超低溫療法脫毒后在甲基化模式中主要由非甲基化位點(diǎn)變?yōu)榧谆稽c(diǎn)。在第2種去甲基化模式中由甲基化位點(diǎn)變?yōu)榉羌谆稽c(diǎn)的比例為48.53%,在第3種甲基化模式中由非甲基化位點(diǎn)變?yōu)榧谆稽c(diǎn)的比例為18.20%(表3)。可見(jiàn),在紅芽芋經(jīng)過(guò)超低溫療法脫毒后,甲基化和去甲基化并存,但主要是以去甲基化為主,說(shuō)明超低溫療法脫毒后較多的基因表達(dá)被激活。
每個(gè)引物擴(kuò)增圖左為HpaⅡ,右為MspⅠ Left of amplification of each primer wasHpaⅡ,and the right wasMspⅠ
圖2E32MSP40、E32MSP48、E35MSP48和E36MSP43引物組合對(duì)樣本1的MSAP擴(kuò)增產(chǎn)物檢測(cè)
Fig.2MSAPamplificationproducttestofsample1basedonE32MSP40,E32MSP48,E35MSP48andE36MSP43
3 討 論
病毒侵染植物是一種生物脅迫[13]。目前有關(guān)生物脅迫對(duì)植物的表觀遺傳影響的研究已有報(bào)道。一般集中于煙草花葉病毒等[14]。對(duì)感染煙草花葉病毒植物的后代進(jìn)一步分析顯示,它們的基因組已大幅度超甲基化[10]。后代的全基因組超甲基化被認(rèn)為是抵抗脅迫的一般保護(hù)機(jī)制的一部分,而重組事件的增加和由病毒攻擊導(dǎo)致的特異性甲基化模式可能是植物適應(yīng)性反應(yīng)的跡象[14]。而這種適應(yīng)性反應(yīng)的跡象將使植物的基因表達(dá)得到抑制,病毒的長(zhǎng)期積累將使植物的某些基因甲基化從而無(wú)法表達(dá),最終導(dǎo)致植物的種性退化、產(chǎn)量和品質(zhì)下降。
研究表明,超低溫處理對(duì)植物材料的甲基化有一定程度的影響。朱文濤等[15]的研究表明,超低溫保存玻璃化法保存成活的五葉草莓甲基化水平降低6.73%,這與木瓜[16]、 蛇麻草[17]、擬南芥[18]等植物超低溫保存后的甲基化變化趨勢(shì)相類似。而超低溫療法脫毒是以超低溫保存的技術(shù)來(lái)快速有效脫除植物病毒,實(shí)質(zhì)上是解除病毒這種生物脅迫的過(guò)程。從理論上講,超低溫療法脫毒是解除病毒的生物脅迫,將會(huì)使植物的某些基因去甲基化,從而在一定程度上恢復(fù)植物的種性,提高其產(chǎn)量和品質(zhì)。
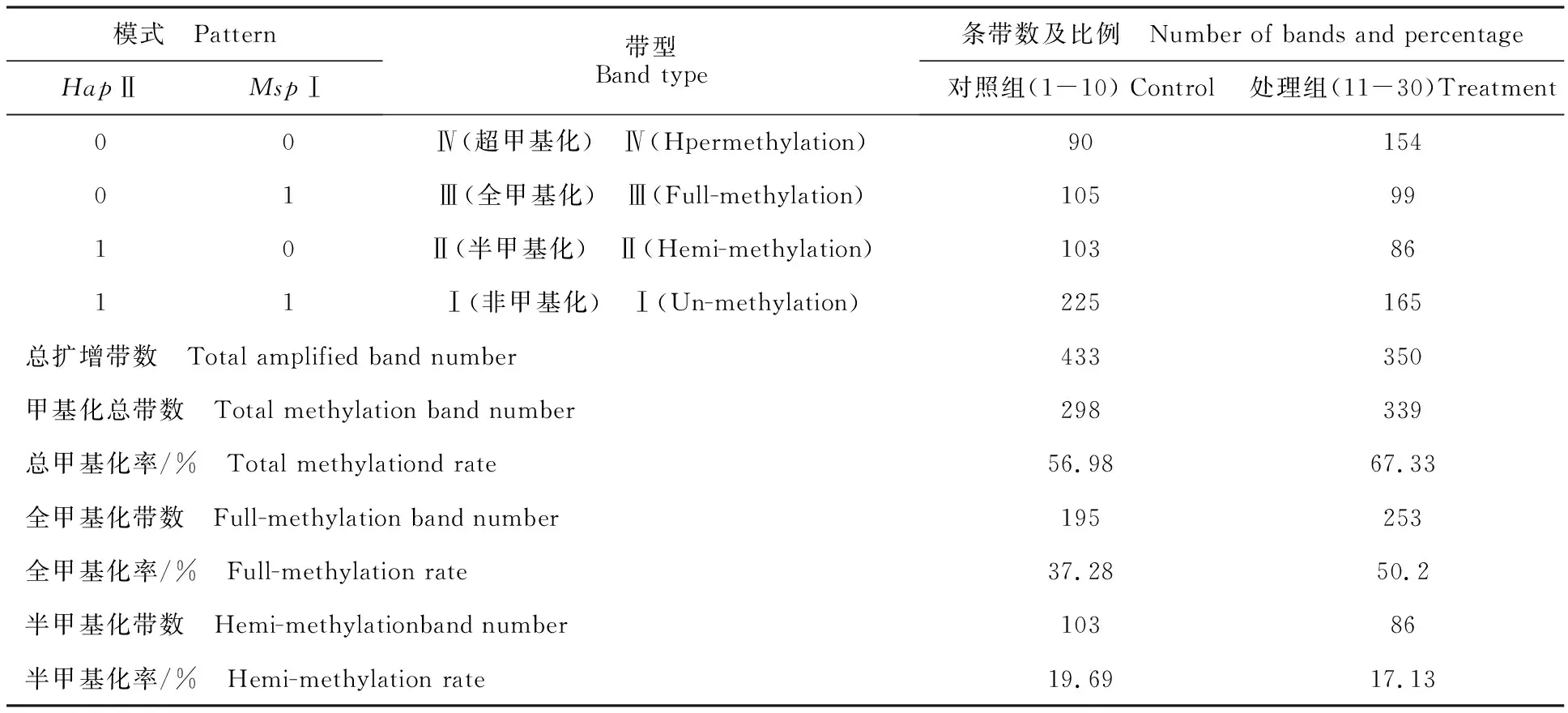
表2 紅芽芋超低溫療法脫毒苗基因組DNA甲基化水平分析Table 2 Analysis of genomic DNA methylation level in virus-free plantlets by cryotherapy of red bud taro
注:總甲基化率=[(Ⅱ+Ⅲ+Ⅳ)]/(Ⅰ+Ⅱ+Ⅲ+Ⅳ)]×100%;全甲基化率=[(Ⅲ+Ⅳ)]/(Ⅰ+Ⅱ+Ⅲ+Ⅳ)]×100%;半甲基化率=[Ⅱ/(Ⅰ+Ⅱ+Ⅲ+Ⅳ)]×100%。
Note:Total methylation rate=[(Ⅱ+Ⅲ+Ⅳ)]/(Ⅰ+Ⅱ+Ⅲ+Ⅳ)]×100%;full-methylation rate=[(Ⅲ+Ⅳ)]/(Ⅰ+Ⅱ+Ⅲ+Ⅳ)]×100%; hemi-methylation rate=[Ⅱ/(Ⅰ+Ⅱ+Ⅲ+Ⅳ)]×100%.
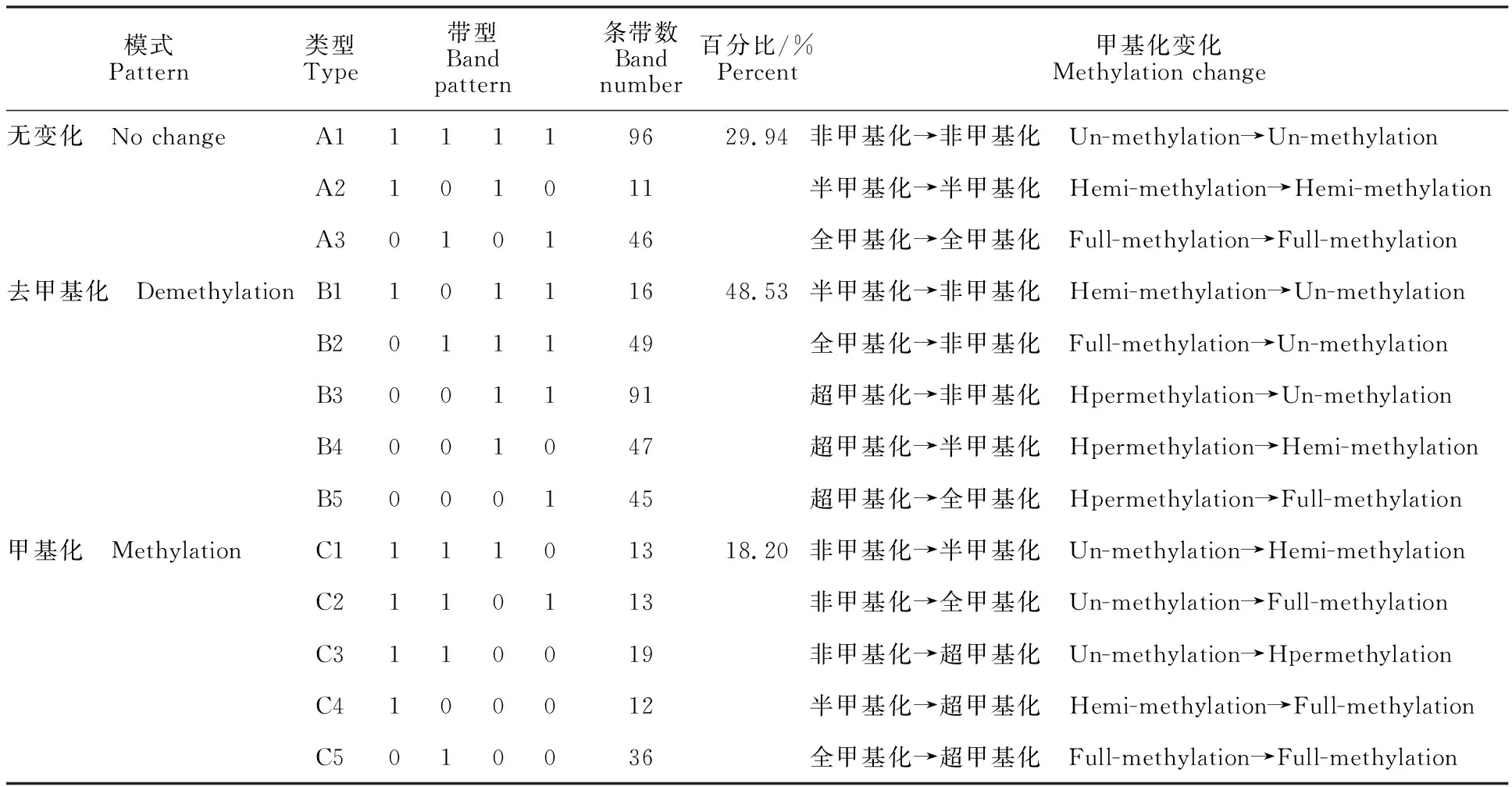
表3 紅芽芋超低溫療法脫毒苗基因組DNA甲基化模式分析Table 3 Analysis of genomic DNA methylation patterns in virus-free plantlets by cryotherapy of red bud taro
但超低溫療法脫毒不同于莖尖常規(guī)培養(yǎng)脫毒,在超低溫療法脫毒過(guò)程中牽涉到超低溫脅迫和滲透脅迫等非生物脅迫的因素,在解除病毒生物脅迫的同時(shí),也遭受到超低溫脅迫和滲透脅迫等非生物脅迫。但目前的研究偏重于對(duì)超低溫保存后再生植株的甲基化研究,沒(méi)有考慮到脫毒方面的效應(yīng),如王芳等[19]對(duì)馬鈴薯莖尖玻璃化法超低溫保存后的 DNA 甲基化遺傳變異進(jìn)行研究,發(fā)現(xiàn)經(jīng)超低溫保存后去甲基化和甲基化都有發(fā)生,但以去甲基化變化為主。何艷霞等[20]在對(duì)擬南芥幼苗的超低溫保存研究中,也發(fā)現(xiàn)有去甲基化現(xiàn)象。這些研究表明,超低溫保存后植物材料存在去甲基化和甲基化,但去甲基化變化是主要趨勢(shì)。在本試驗(yàn)中,以超低溫療法脫毒的紅芽芋為研究對(duì)象,考察超低溫保存和脫毒對(duì)紅芽芋甲基化水平和甲基化模式的影響,發(fā)現(xiàn)超低溫療法脫毒苗總甲基化率提高10.27%,全甲基化率提高12.98%,但半甲基化率下降2.71%。紅芽芋超低溫療法脫毒苗去甲基化模式和甲基化模式并存,但去甲基化模式高于甲基化模式,說(shuō)明超低溫療法脫毒后有較多的基因表達(dá)被激活。究其原因,可能是脫毒和超低溫保存兩者平衡的結(jié)果,一是解除病毒生物脅迫,二是給予超低溫脅迫和滲透脅迫等非生物脅迫,最終去甲基化模式高于甲基化模式可能也是植物適應(yīng)性反應(yīng)的跡象,導(dǎo)致更多的基因得到表達(dá),低溫療法脫毒可恢復(fù)種性的機(jī)理可以從甲基化水平和甲基化模式去解釋。這種變化趨勢(shì)可能是植物對(duì)超低溫保存這種逆境處理和病毒脅迫解除的一種綜合適應(yīng),其內(nèi)在機(jī)制目前未明,需要深入探討。另外,超低溫療法脫毒中的甲基化能否引起基因的差異表達(dá),也需要更深入地探討。本研究結(jié)果可為紅芽芋脫毒苗的種質(zhì)保存和規(guī)模化生產(chǎn)提供理論依據(jù)。
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級(jí)版(2021年3期)2021-03-19 06:12:40
小天使·二年級(jí)語(yǔ)數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫(huà)刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
