食源性致病菌生長延滯期建模的研究進(jìn)展
2019-01-28 06:09:54岳思遠(yuǎn)任鵬程劉陽泰董慶利
食品科學(xué) 2019年1期
岳思遠(yuǎn),蘇 亮,任鵬程,劉陽泰,王 翔,劉 箐,董慶利,*
(1.上海理工大學(xué)醫(yī)療器械與食品學(xué)院,上海 200093;2.國家食品安全風(fēng)險評估中心,北京 100022)
食源性致病菌是一類以食品為傳播媒介,可引起食物中毒的致病性微生物,是目前食源性疾病的主要誘因。由食源性致病菌引起的食源性疾病是國際性公共衛(wèi)生問題。世界衛(wèi)生組織2017年的報告表明,全球食源性疾病患者約6億,由食源性疾病引起的死亡人數(shù)約42萬,其中5歲以下兒童死亡人數(shù)約12.5萬[1]。近年來我國建立了食源性疾病監(jiān)測信息數(shù)據(jù)庫,系統(tǒng)分析監(jiān)測網(wǎng)地區(qū)14 年間8 000余起食物中毒案例,發(fā)現(xiàn)微生物性病原仍然是我國食源性疾病的主要病因(占30%~40%)[2]。同時,我國國家衛(wèi)生健康委員會(原國家衛(wèi)生和計(jì)劃生育委員會)辦公廳發(fā)布2017年第4季度的食物中毒公告顯示,食物中毒事件報告數(shù)量和中毒人數(shù)最多的是微生物性食物中毒事件,分別占總報告數(shù)量和總中毒人數(shù)的34.6%和55.5%[3]。因此,需嚴(yán)格控制食品中食源性致病菌的數(shù)量及生長,以保證食品食用的安全性。
近年來,隨著人們對食品安全的重視,預(yù)測微生物學(xué)也得到了良好的發(fā)展。通常情況下,微生物生長一般會經(jīng)歷4 個階段:延滯期、對數(shù)期、穩(wěn)定期及衰亡期。目前微生物延滯期仍未基于微生物生長機(jī)理明確定義,生物學(xué)上被普遍接受的定義是:微生物群體經(jīng)歷環(huán)境突變,自我調(diào)整后開始繁殖的時間[4]。幾何意義上微生物延滯期是指微生物對數(shù)生長期達(dá)到最大比生長速率時,生長曲線切線的反向延長線和初始菌量水平延長線的交點(diǎn)在時間軸上的投影點(diǎn)與零時刻的時間間隔[5]。在傳統(tǒng)預(yù)測微生物生長的模型中,延滯期常通過生長曲線確定。而應(yīng)用數(shù)學(xué)模型預(yù)測微生物生長時,延滯期較最大比生長速率受更多因素影響,因此延滯期較最大比生長速率更難以準(zhǔn)確獲取。
預(yù)測微生物學(xué)作為食品風(fēng)險評估的重要工具,已被廣泛應(yīng)用于食品安全風(fēng)險控制中,然而因延滯期尚未明確定義,且影響因素較多,相較于最大比生長速率預(yù)測模型更難以準(zhǔn)確估計(jì)微生物生長延滯期,因此延滯期研究仍具有較大的發(fā)展空間,需進(jìn)一步探討。以下分述微生物生長延滯期的測定方法和建模方法的研究進(jìn)展。
1 延滯期的測定方法
現(xiàn)有技術(shù)并不能直接測定微生物生長延滯期,需通過已建立的微生物生長模型擬合生長曲線,間接獲取延滯期。在預(yù)測微生物學(xué)中,微生物生長模型的建立需確定微生物生長曲線,微生物生長曲線的獲取和記錄可采用多種方法。基于微生物群體水平和單細(xì)胞水平下不同的延滯期概念,延滯期的測定分別從群體水平和單細(xì)胞水平進(jìn)行論述。
1.1 群體生長延滯期的測定
目前測定微生物生長曲線的方法主要分為傳統(tǒng)方法和分子生物學(xué)方法。表1中總結(jié)了應(yīng)用傳統(tǒng)方法和分子生物學(xué)方法測定微生物生長曲線進(jìn)而通過模型擬合獲取生長延滯期的相關(guān)研究。傳統(tǒng)測定微生物生長曲線的方法是活菌計(jì)數(shù)法,因其不受菌懸液顏色及濁度的影響而成為測定食品中食源性致病菌的主流手段。但是該方法操作繁瑣、耗時耗力,且應(yīng)用該方法獲取的生長參數(shù)準(zhǔn)確性與數(shù)據(jù)點(diǎn)的數(shù)量及觀測點(diǎn)的位置均相關(guān)[23],因此活菌計(jì)數(shù)法還需進(jìn)一步改進(jìn)。相較于活菌計(jì)數(shù)法,比濁法因其操作簡便、省時省力的優(yōu)點(diǎn)被廣泛應(yīng)用于微生物生長曲線的測定和建模研究中[12,24]。然而,比濁法的檢測限僅能達(dá)到106~107CFU/mL,且只可用于液體培養(yǎng)基的測定,同時因測定的菌懸液濃度并不是活菌濃度而導(dǎo)致獲得的生長參數(shù)有些許誤差。建議今后的研究可基于技術(shù)層面對該方法進(jìn)行優(yōu)化,使其能更好地描述低濃度食源性致病菌的生長,為進(jìn)一步研究微生物生長提供依據(jù)。
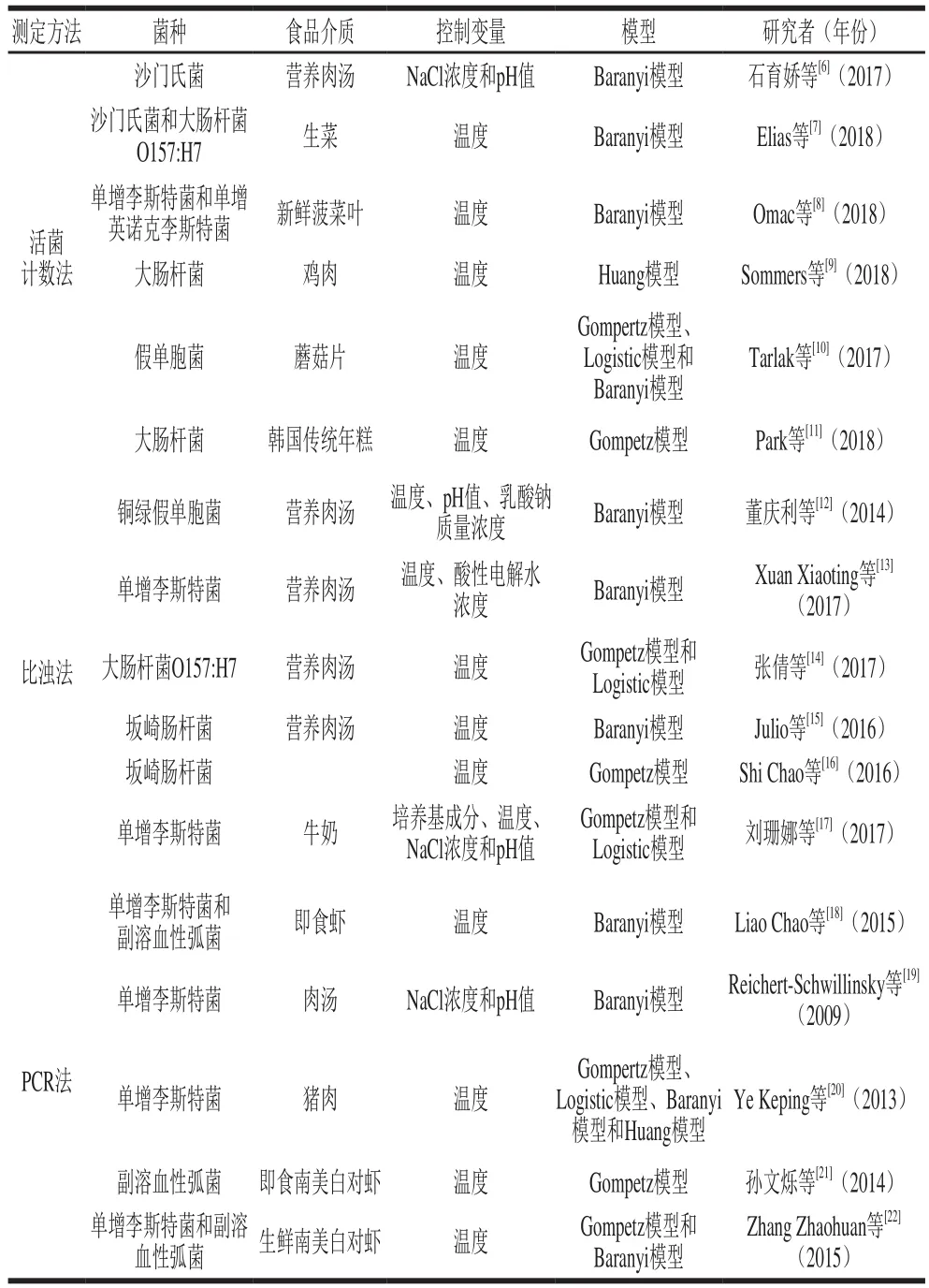
表1 應(yīng)用各種方法測定食源性致病菌生長曲線的相關(guān)研究Table1 A summary of methods for determining the growth curve of foodborne pathogenic bacteria
隨著分子生物學(xué)技術(shù)的不斷發(fā)展,變性梯度凝膠電泳(denatured gradient gel electrophoresis,DGGE)技術(shù)和PCR等分子生物學(xué)技術(shù)也被應(yīng)用于微生物生長曲線的測定,相較于傳統(tǒng)方法,這些分子生物學(xué)技術(shù)具有省時省力、高效等優(yōu)點(diǎn),且可同時測定兩種細(xì)菌的生長,極大地改善了傳統(tǒng)方法的不足[25]。Liao Chao等[18]應(yīng)用DGGE結(jié)合PCR技術(shù)測定了單增李斯特菌和副溶血性弧菌的生長情況,進(jìn)而通過Baranyi模型擬合其生長曲線獲取延滯期。傳統(tǒng)PCR技術(shù)需結(jié)合DGGE技術(shù)才可定量測定微生物的生長情況,而實(shí)時熒光定量PCR(real-time quantitative PCR,qPCR)技術(shù)[26]的出現(xiàn)彌補(bǔ)了其缺陷,實(shí)現(xiàn)了從定性到定量的突破。Guan Zhengping等[27]將金黃色葡萄球菌的菌液接種于豬肉中進(jìn)行培養(yǎng),每隔一段時間提取生長后細(xì)菌的總基因組DNA,并對其進(jìn)行qPCR分析,進(jìn)而根據(jù)已建立的微生物濃度與qPCR的循環(huán)閾(cycle threshold,Ct)值間的標(biāo)準(zhǔn)曲線,定量描述豬肉中金黃色葡萄球菌的生長情況,最后應(yīng)用模型擬合其生長曲線,間接獲取生長延滯期。Ye Keping[20]和孫文爍[21]等也同樣應(yīng)用qPCR技術(shù)分別定量描述了豬肉中單增李斯特菌和即食南美白對蝦中副溶血性弧菌的生長情況,并對其進(jìn)行模型擬合,獲取了延滯期。但因qPCR技術(shù)無法區(qū)分活菌和死菌,導(dǎo)致實(shí)驗(yàn)結(jié)果有一定的誤差。基于此,Zhang Zhaohuan等[22]將疊氮溴化丙錠(propidium monoazide,PMA)與多重qPCR技術(shù)相結(jié)合,極好地定量描述了生鮮南美白對蝦樣品中副溶血性弧菌和單增李斯特菌的生長行為。雖然PMA的應(yīng)用可區(qū)分活死菌,但其會對細(xì)菌產(chǎn)生一定的損傷。分子生物學(xué)技術(shù)的發(fā)展為預(yù)測微生物學(xué)注入了新的活力,加速了預(yù)測微生物學(xué)的發(fā)展。然而,任何方法均需辯證地看待,即每一種方法均各有優(yōu)點(diǎn)和局限性,因此分子生物學(xué)技術(shù)并不能完全取代傳統(tǒng)方法,仍需根據(jù)具體的實(shí)驗(yàn)?zāi)康倪x擇合適的實(shí)驗(yàn)方法。
1.2 單細(xì)胞生長延滯期的測定
單細(xì)胞生長延滯期的研究對象為單細(xì)胞,其分布研究需獲取大量的實(shí)驗(yàn)數(shù)據(jù)。因此,傳統(tǒng)的活菌計(jì)數(shù)法并不適用于開展單細(xì)胞水平研究,需采用其他方法測定單細(xì)胞延滯期。
單細(xì)胞生長延滯期可通過直接觀測單細(xì)胞分裂過程進(jìn)行測定。最早可追溯到Kelly等[28]的相關(guān)研究,將少量微生物接種于瓊脂薄膜上,并置于顯微鏡下觀測微生物單細(xì)胞個體的分裂過程,結(jié)果表明,隨著細(xì)胞傳代次數(shù)的增加,分裂時間間隔隨之下降。基于固體培養(yǎng)觀測法,董慶利等[29]設(shè)計(jì)流動槽裝置,顯微觀測銅綠假單胞菌單細(xì)胞的生長分裂情況,流動槽通過液體流動帶走新分裂的子細(xì)胞,進(jìn)而持續(xù)觀測一個目標(biāo)細(xì)胞多次分裂的情況。固體培養(yǎng)顯微觀測法和液體培養(yǎng)顯微觀測法的實(shí)質(zhì)均是通過單細(xì)胞分裂時間計(jì)算單細(xì)胞延滯期。雖然顯微觀測法可直接觀測單細(xì)胞生長分裂的過程,但是該法對儀器設(shè)備要求較高且耗時耗力。同時,采用顯微觀測法開展單細(xì)胞延滯期分布的研究難以獲取大量實(shí)驗(yàn)數(shù)據(jù)。
比濁法通過間接推斷計(jì)算單細(xì)胞生長延滯期。與群體水平的觀測不同,比濁法應(yīng)用Bioscreen FP-1100C型全自動微生物生長曲線分析儀實(shí)時連續(xù)記錄單細(xì)胞生長情況,進(jìn)而通過檢測時間(the time to detection,Tdet)計(jì)算單細(xì)胞延滯期。比濁法中的檢測時間是指初始OD值增加至對應(yīng)于菌懸液菌落總數(shù)為107CFU/mL時OD值的時間。McKellar[30]通過不同稀釋度的菌液與檢測時間線性回歸獲得回歸曲線的斜率(slope),進(jìn)而計(jì)算最大比生長速率(μmax),計(jì)算公式如式(1)所示。

進(jìn)而再采用已得的μmax和模型估計(jì)Tdet,預(yù)測得到的Tdet和實(shí)測Tdet的差值即為單細(xì)胞延滯期。此后,Baranyi等[31]在前人研究的基礎(chǔ)上,改進(jìn)了單細(xì)胞延滯期的計(jì)算方式。假設(shè)Bioscreen微型孔板100 孔中每個孔的細(xì)胞數(shù)為1,且單細(xì)胞生長數(shù)量服從泊松分布[32],同時,細(xì)菌在對數(shù)期以恒定生長速率(μmax)生長,則單細(xì)胞延滯期即可通過公式(2)計(jì)算,生長曲線如圖1所示[5]。
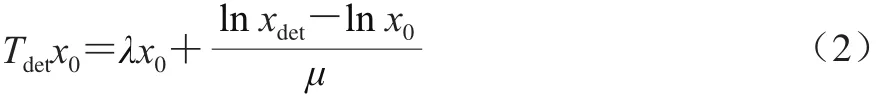
式中:x0為初始細(xì)胞數(shù)/(CFU/mL);xdet為達(dá)到檢測時間的細(xì)胞數(shù)/(CFU/mL);Tdet為初始OD值達(dá)到菌懸液菌落總數(shù)為107CFU/mL所對應(yīng)OD值的時間/h;μ為對數(shù)期細(xì)菌生長的恒定生長速率/h-1;λ為單細(xì)胞生長延滯期/h。
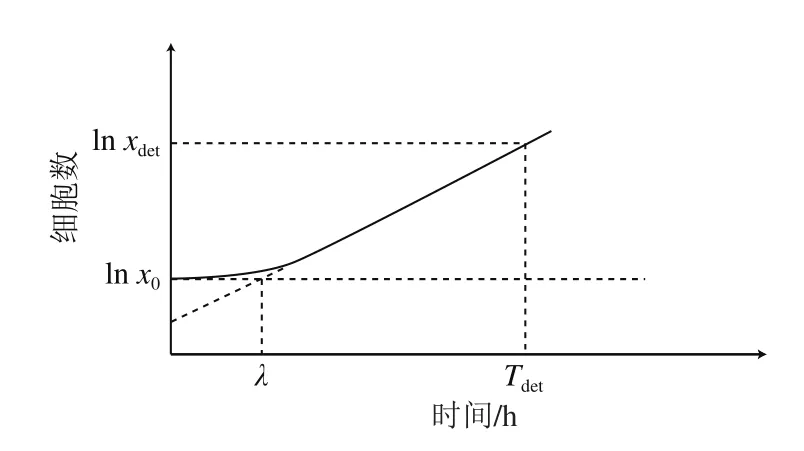
圖1 微生物生長曲線[5]Fig.1 Microbial growth curve[5]
經(jīng)Bioscreen FP-1100C型全自動微生物生長曲線分析儀測定單細(xì)胞延滯期的比濁法可連續(xù)監(jiān)測大量單細(xì)胞的生長情況且省時省力,已廣泛用于單細(xì)胞延滯期分布的研究。
綜上所述,雖然微生物群體延滯期和單細(xì)胞延滯期均可采用上述方法間接獲得,但是微生物群體延滯期會因擬合模型不同而不同,同時微生物單細(xì)胞生長觀測方法依然存在精度低、控制難、耗時長等問題。因此,需在明確延滯期定義的基礎(chǔ)上,進(jìn)一步改進(jìn)已有的微生物觀測手段以提高生長模型預(yù)測延滯期的準(zhǔn)確性。
2 延滯期的建模方法
在預(yù)測微生物學(xué)中,微生物生長模型一般根據(jù)變量類型分為三大類:一級模型是描述恒定環(huán)境條件下微生物數(shù)量與時間的關(guān)系;二級模型是描述微生物生長參數(shù)(延滯期(λ)和最大比生長速率(μmax))與環(huán)境變量(如溫度、pH值和水分活度等)間的關(guān)系;三級模型是基于一級模型和二級模型,應(yīng)用計(jì)算機(jī)軟件獲取微生物生長參數(shù)[33]。
基于微生物生長延滯期的測定方法,微生物群體延滯期需通過生長模型擬合生長曲線間接獲取。表2歸納了可描述微生物生長曲線的一級模型。

表2 預(yù)測微生物學(xué)中常見的一級模型Table2 Primary models in predictive microbiology
由表2可知,與其他模型相比,三段式線性模型因分段描述微生物生長較難實(shí)現(xiàn)擬合。而Gompertz模型和Logistic模型最初并不是基于微生物學(xué)概念建立的模型。同時,Gompertz模型描述的S型曲線中,拐點(diǎn)處的確定曲率會導(dǎo)致高估延滯期[39-40]。與其他模型相比,Huang模型與Baranyi模型有一定的生理學(xué)意義,均為機(jī)械模型。
在預(yù)測微生物學(xué)中,被人普遍接受的一級模型為Baranyi模型[37],該模型引入了細(xì)胞生理狀態(tài)(Q)這一生理學(xué)概念,公式如式(3)、(4)所示。
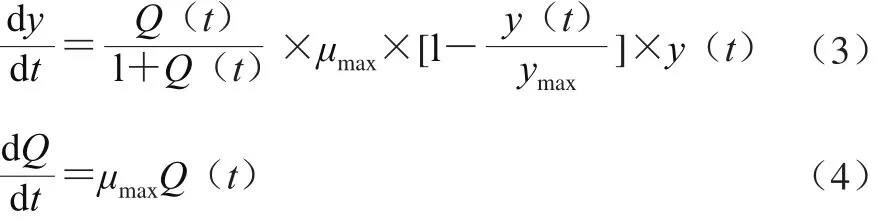
將Baranyi模型式(3)中的Q(t)/(1+Q(t))為調(diào)整函數(shù)α(t)。α(t)描述微生物延滯期向?qū)?shù)生長期的轉(zhuǎn)換。假設(shè)微生物生長由某一物質(zhì)P(t)控制,且該物質(zhì)的累積遵循Michaelis-Menten動力學(xué),則α(t)的計(jì)算如式(5)所示。
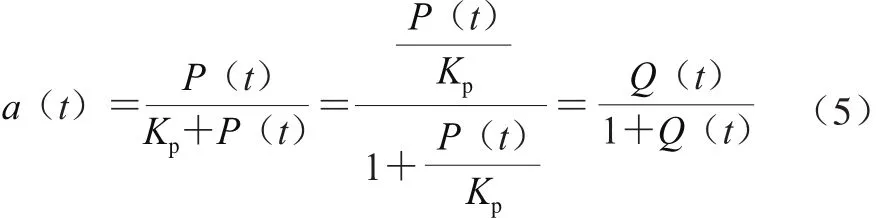
式中:Kp為米氏常數(shù),當(dāng)微生物生長過程中環(huán)境條件改變時,調(diào)整函數(shù)α(t)取決于P(t)/Kp速率。P(t)/Kp速率用于表征細(xì)胞的生理狀態(tài)Q(t)。Baranyi模型中延滯期可用細(xì)胞生理狀態(tài)表示,公式如式(6)所示。
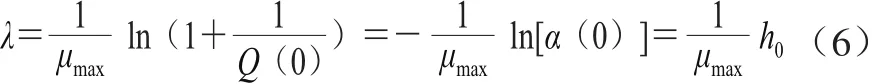
式中:Q(0)為細(xì)胞的初始生理狀態(tài);h0為α(t)與Q(t)在數(shù)學(xué)意義上的轉(zhuǎn)換參數(shù)。
除延滯期一級建模外,還存在延滯期的二級建模。目前最常用的延滯期二級模型是Ratkowsky等[41]于1982年提出的平方根模型,用于描述微生物延滯期隨溫度的變化情況,具體表達(dá)式如式(7)所示。

式中:λ為微生物生長延滯期/h;b為平方根模型的系數(shù);T為微生物培養(yǎng)溫度/℃;Tmin為微生物理論最低生長溫度/℃。
延滯期的二級模型僅適用于描述微生物延滯期隨溫度的變化,而微生物所處生長環(huán)境較為復(fù)雜,因此,延滯期二級模型存在一定的局限性,建議在建模過程中增加其他影響因素。
延滯期的獲取一般需將實(shí)驗(yàn)獲取的微生物生長數(shù)據(jù)輸入到三級模型即計(jì)算機(jī)軟件中,進(jìn)而再選擇軟件中的一級模型進(jìn)行擬合,獲得相應(yīng)的生長參數(shù),其中包括延滯期。表3總結(jié)了目前國內(nèi)外可獲取延滯期的相關(guān)軟件,其中以英國食品研究所開發(fā)的ComBase和美國農(nóng)業(yè)部開發(fā)的病原菌模型程序(pathogen modeling program,PMP)最為著名[45]。由表3可知,每個軟件中所包含的一級模型并不是很全面,因此,建議在日后的軟件開發(fā)方面,盡可能將所有一級模型編入程序中,供使用者自由選擇合適的生長模型進(jìn)行延滯期的獲取。
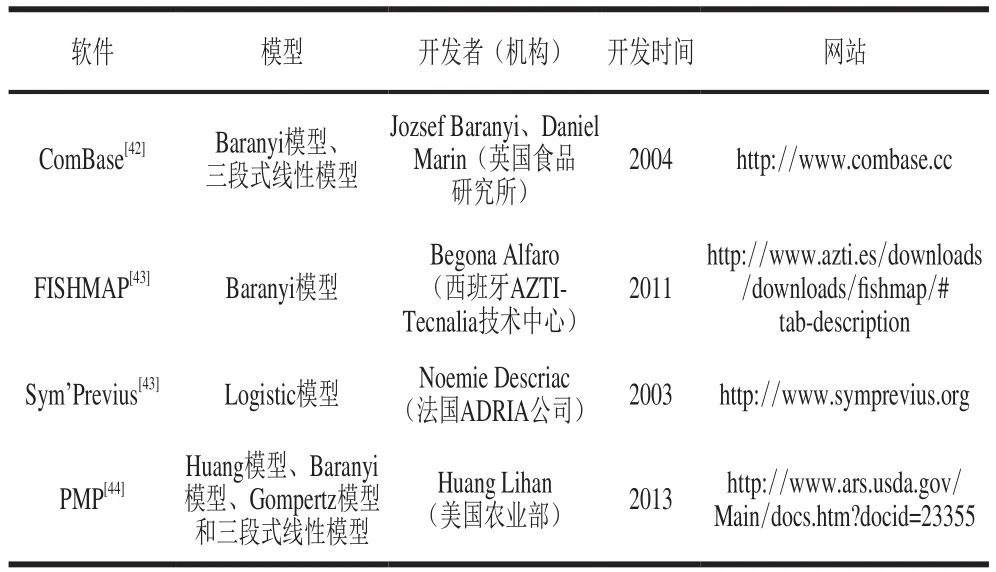
表3 預(yù)測微生物學(xué)中的相關(guān)軟件Table3 Overview of predictive microbiology software tools
綜上所述,雖然已有諸多一級模型可描述微生物生長,但是現(xiàn)有一級模型均是基于群體建立的,未考慮單細(xì)胞生長變異性。同時,因已有一級模型中的數(shù)學(xué)概念并不能通過實(shí)驗(yàn)驗(yàn)證,還需進(jìn)一步改進(jìn)已有模型或建立新模型以提高生長模型預(yù)測延滯期的準(zhǔn)確性。除此之外,基于現(xiàn)有一級模型,還需在已有的計(jì)算機(jī)軟件中添加部分一級模型,或開發(fā)新的覆蓋所有一級模型的計(jì)算機(jī)軟件,從而在應(yīng)用計(jì)算機(jī)軟件進(jìn)行模型擬合時實(shí)現(xiàn)一級模型的最優(yōu)選擇。
3 結(jié) 語
雖然目前的已有研究可觀測食源性致病菌的生長,也可應(yīng)用模型擬合生長曲線獲得生長延滯期,但仍需改進(jìn)測定食源性致病菌生長的方法和擬合食源性致病菌生長的模型,進(jìn)一步完善食源性致病菌生長延滯期建模工作。建議食源性致病菌生長延滯期建模及預(yù)測的研究方向如下:首先,應(yīng)從機(jī)理出發(fā),進(jìn)一步明確食源性致病菌生長延滯期定義,為延滯期建模提供理論依據(jù);其次,基于目前已有的延滯期觀測手段,改進(jìn)傳統(tǒng)計(jì)數(shù)法耗時費(fèi)力等問題,或優(yōu)化分子生物學(xué)技術(shù)的精度及特異性,為延滯期的準(zhǔn)確預(yù)測提供幫助;最后,延滯期建模中的模型仍存在一定問題,建議改進(jìn)現(xiàn)有一級模型的部分參數(shù)或基于延滯期機(jī)理建立新的延滯期模型,同時優(yōu)化已有三級模型或開發(fā)新的三級模型,使其覆蓋所有延滯期模型,為延滯期建模的發(fā)展提供強(qiáng)有力的支持。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
小讀者(2021年2期)2021-03-29 05:03:48
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
數(shù)學(xué)物理學(xué)報(2020年2期)2020-06-02 11:29:24
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
Coco薇(2016年2期)2016-03-22 02:42:52
Coco薇(2015年1期)2015-08-13 02:47:34
