SPAC1006.01基因敲除對裂殖酵母生長及產孢的影響
2019-02-15 08:08:10袁榮美丁祥譚秀梅侯怡鈴
科技與創(chuàng)新 2019年1期
關鍵詞:生長
袁榮美,丁祥,譚秀梅,侯怡鈴
?
SPAC1006.01基因敲除對裂殖酵母生長及產孢的影響
袁榮美1,丁祥2,譚秀梅1,侯怡鈴1
(1.西華師范大學生命科學學院,西南野生動植物資源保護教育部重點實驗室,四川 南充 637009; 2.西華師范大學環(huán)境科學與工程學院,四川 南充 637009)
為研究SPAC1006.01基因對裂殖酵母(Schizosaccharomyces pombe)生長以及孢子數(shù)目和形態(tài)的影響,將野生型與SPAC1006.01基因敲除突變株在常溫及高溫脅迫下培養(yǎng),通過測量OD595繪制生長曲線圖,看其生長是否受到影響;同時將基因敲除突變株的h+和h-菌株結合,觀察其產生的孢子與野生型菌株接合產生的孢子的差異性。結果顯示,在25 ℃下,野生型菌株正常生長,基因敲除突變株比野生型生長緩慢;在37 ℃下,野生型菌株生長速度加速,但基因敲除突變株仍然比野生型生長緩慢,尤其在12 h后,差別明顯。產孢實驗結果表明,野生型100%產生4個孢子,在基因敲除突變菌株中,有10%的菌接合產生1個孢子;30%的菌接合產生2個孢子;15%的菌接合產生3個孢子;只有45%的菌產孢正常,產生4個孢子。但兩者所產孢子形態(tài)正常。實驗結果表明SPAC1006.01基因能影響裂殖酵母的生長和產孢數(shù)目。
裂殖酵母;基因敲除;生長曲線;子囊孢子
SPAC1006.01基因也叫psp3,在裂殖酵母中它的表達產物為液泡絲氨酸蛋白酶psp3(預測),由451個氨基酸組成,分子量為48.73 kDa。
液泡絲氨酸蛋白酶psp3具有絲氨酸型內肽酶活性[1],可通過絲氨酸親核體組成的催化三聯(lián)體機制催化多肽鏈中內部α-肽鍵的水解;具有肽酶活性[2],在液泡中起作用,可以催化肽鍵的水解,參與液泡蛋白的水解加工[3]。
科學家研究發(fā)現(xiàn),在psp3Δ中,由于psp3基因的缺失,會影響cnx1基因(cnx1是酵母中蛋白質編碼的基因)的表達,可能影響所有蛋白質或一種或多種特定蛋白質的加工[4]。psp3Δ在營養(yǎng)生長期間,細胞的體積、大小和形狀均是正常的,但其對化學品的敏感性增加。在野生型菌株能夠生長的硼替佐米濃度下,突變株psp3Δ對硼替佐米的敏感性增加,具體表現(xiàn)為細胞停止生長(并可能死亡)[5]。最近的科學研究發(fā)現(xiàn),psp3基因與SPAC4A8.04基因具有很高的同源性,SPAC4A8.04基因在裂殖酵母中的功能是誘導裂殖酵母孢子的形成,為進一步探究psp3基因是否也對裂殖酵母產孢有影響,本研究以突變株psp3Δ為材料,對其進行生長曲線測定和產孢實驗。
1 材料與方法
1.1 材料
1.1.1 酵母菌株
本研究所使用的裂殖酵母(Schizosaccharomyces pombe)野生型和突變株psp3Δ保存于西華師范大學生命科學學院西南野生動植物資源保護教育部重點實驗室,如表1所示。
表1 本研究所使用的酵母菌株
菌株敲除基因雌雄來源 PT286野生型h-實驗室保存 PT287野生型h+實驗室保存 108SPAC1006.01h+實驗室保存
1.1.2 裂殖酵培養(yǎng)基及藥品
裂殖酵母完全培養(yǎng)液YES(1 L):5 g酵母提取物(Thermo Fisher Scientific 公司),30 g葡萄糖(Thermo Fisher Scientific 公司),225 mg組氨酸(本實驗所使用的氨基酸、嘌呤、嘧啶均購自Sigma公司),225 mg腺嘌呤,225 mg賴氨酸,225 mg亮氨酸,225 mg尿嘧啶,如果為固體培養(yǎng)基則加入17 g瓊脂[6](Thermo Fisher Scientific公司)。
裂殖酵母產孢培養(yǎng)基EMM-N(1 L):7.32 g脫水EMM-N培養(yǎng)基粉末(生物風資源技術服務公司),20 g葡萄糖,20 g瓊脂,225 mg亮氨酸,225 mg腺嘌呤,225 mg尿嘧啶,225 mg組氨酸,225 mg賴氨酸。
藥品:PBS緩沖液(pH 4.4)、纖維素酶(阿拉丁公司)。
1.1.3 主要儀器
主要儀器:LDZX-30FA全自動壓力蒸汽滅菌鍋(上海申安醫(yī)療器械廠),SE-CJ-1F型超凈工作臺(蘇州安泰空氣技術有限公司),THZ-Q恒溫搖床(上海百典儀器設備有限公司),Epoch酶標儀(美國基因有限公司),DPX-9272B-1電熱恒溫培養(yǎng)箱(上海福瑪實驗設備有限公司),AR2130電子分析天平(上海奧豪斯儀器有限公司),BCD-208K/A型﹣80 ℃冰箱(青島海爾股份有限公司),Eppendorf移液槍(艾本德(上海)國際貿易有限公司),HHS型水浴鍋(上海光地儀器設備有限公司),LD4-2A型離心機,MD50光學顯微鏡(OLYMPUS公司)。
1.2 實驗方法
1.2.1 菌種復蘇
將用無菌甘油保存于﹣80 ℃的108、PT287和PT286菌種取出;挑一小塊菌液冰塊,均勻涂布到YES固體培養(yǎng)基上,置于25 ℃恒溫培養(yǎng)箱培養(yǎng)(約48 h)。
1.2.2 菌種活化
從YES固體培養(yǎng)基上刮取PT286和108菌體,接種至4 mL完全培養(yǎng)液中,置于25 ℃,120 r/min恒溫搖床振蕩培養(yǎng)過夜(約24 h);測得OD595(酶標儀)為0.5~0.8,表示菌種已經活化好。
1.2.3 生長曲線的測定
將活化好的裂殖酵母菌液稀釋到OD595為0.1左右;分別置于25 ℃和37 ℃恒溫搖床中培養(yǎng),每2 h取200 uL菌液用酶標儀檢測OD595,重復3次;收集數(shù)據(jù),并用Excel進行處理,以培養(yǎng)時間為橫坐標,OD595為縱坐標,繪制生長曲線[7]。
1.2.4 突變株108 h-菌株的構建
挑取活化好的PT286和108菌種,充分混合均勻后涂布在EMM-N培養(yǎng)基上;置于25 ℃的恒溫培養(yǎng)箱中培養(yǎng)(約24 h);24 h后鏡檢,觀察有無孢子產生。確定有子囊孢子產生后,從EMM-N固體培養(yǎng)基上收集子囊孢子,加入1 mL纖維素酶工作液(PBS緩沖液配制),置于37 ℃水浴鍋中裂解過夜(24 h左右);鏡檢,觀察細胞壁是否裂解完全; 3 000 r/m離心10 min,收集菌體;加入1 mL蒸餾水重懸菌體進行洗滌,3 000 r/m離心10 min,收集菌體,重復洗滌一次;將收集到的菌體均勻涂布在YES培養(yǎng)基上,挑取單克隆,擴大培養(yǎng),再與野生型PT286接合,如果產孢,則確定其配型為h+;如果不產孢,則為h-。
1.2.5 裂殖酵母產孢及孢子形態(tài)觀察
將108菌株的h+與h-接合作為實驗組,野生型PT286與PT287接合作為對照組,置于25 ℃恒溫培養(yǎng)箱培養(yǎng)過夜(約24 h);制片,使用光學顯微鏡選取不同視角觀察并統(tǒng)計20個產孢酵母的孢子數(shù),截取典型孢子形態(tài)圖。
2 結果與分析
2.1 野生型及基因敲除突變株在25 ℃條件下的生長曲線
野生型和基因敲除突變株在25 ℃條件下生長曲線如圖1所示,25 ℃條件下,0~6 h野生型和突變株108生長速度差別不明顯;6 h后,野生型生長速度加快,而突變株108一直較為平緩,在12 h后,野生型OD595已經達到0.623,而突變株108僅有0.398,與野生型差別較大。
實驗結果表明,在溫度條件為25 ℃時,SPAC1006.01基因以及其編碼的蛋白質與裂殖酵母的生長相關,基因敲除后,導致菌株生長緩慢。

圖1 野生型和基因敲除突變株在25 ℃條件下生長曲線
2.2 野生型及基因敲除突變株在37 ℃條件下的生長曲線
野生型和基因敲除突變株在37 ℃條件下生長曲線由圖2所示,溫度為37 ℃時,0~6 h內野生型與突變株生長趨勢基本一致,6 h后野生型生長速度陡增,而突變株108生長速度一直較為平緩。尤其在12 h后,野生型菌株OD595達到了0.843 4,突變株108在12 h后OD595為0.373,與25 ℃條件下基本沒有差別。
由此可知,無論是在25 ℃條件下還是在37 ℃條件下,SPAC1006.01基因及其編碼的蛋白質都會影響裂殖酵母的生長,且基因敲除后,導致菌株對溫度不敏感。但其具體的分子機制尚不清楚,有待進一步研究。

圖2 野生型和基因敲除突變株在37 ℃條件下生長曲線
2.3 野生型與基因敲除突變株的子囊孢子數(shù)目及形態(tài)比較
野生型與基因敲除突變株108產生的子囊孢子數(shù)量對比如圖3所示。
由圖3可知,野生型菌株100%都會產生4個大小均勻的子囊孢子,而突變株108產生子囊孢子的菌中有10%產生1個孢子、30%產生2個孢子、15%產生3個孢子,只45%的菌能夠正常產生4個孢子。
野生型與突變株的產孢數(shù)目對比如圖4所示,野生型PT286和突變株108產生的子囊孢子形態(tài)并沒有差別。實驗結果說明,sps基因及其編碼的蛋白質會影響裂殖酵母產生子囊孢子的數(shù)量,但不影響孢子形態(tài)。
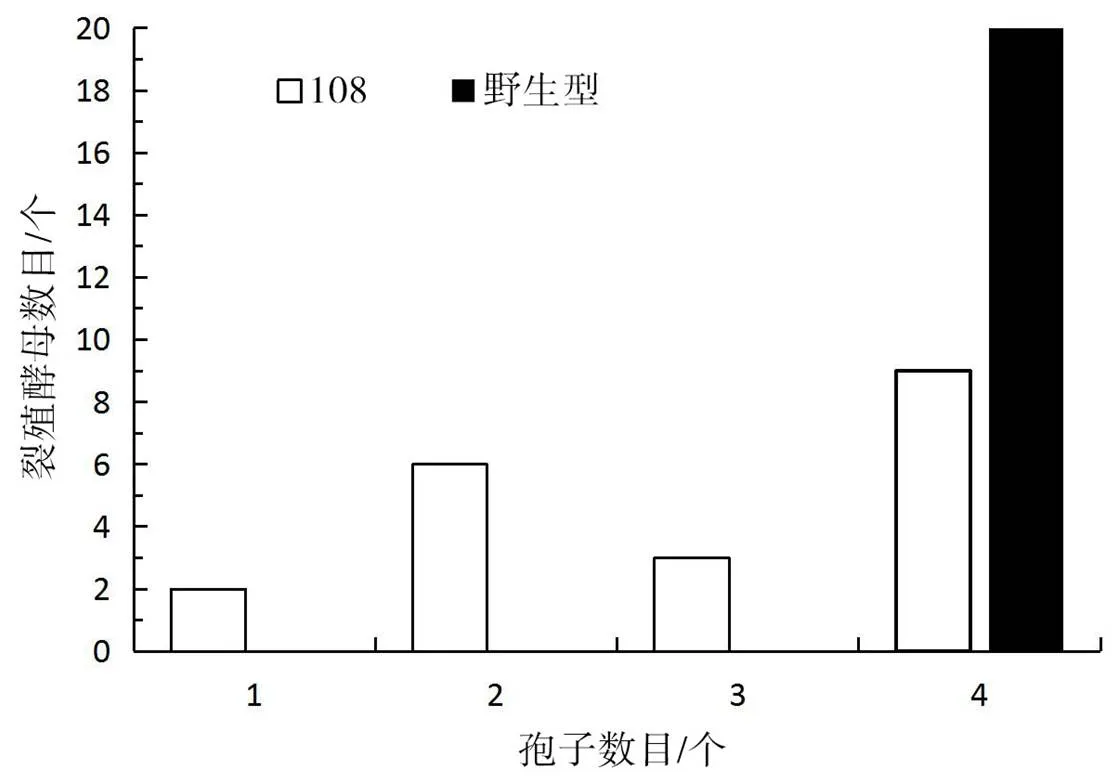
圖3 野生型與突變株的產孢數(shù)目對比
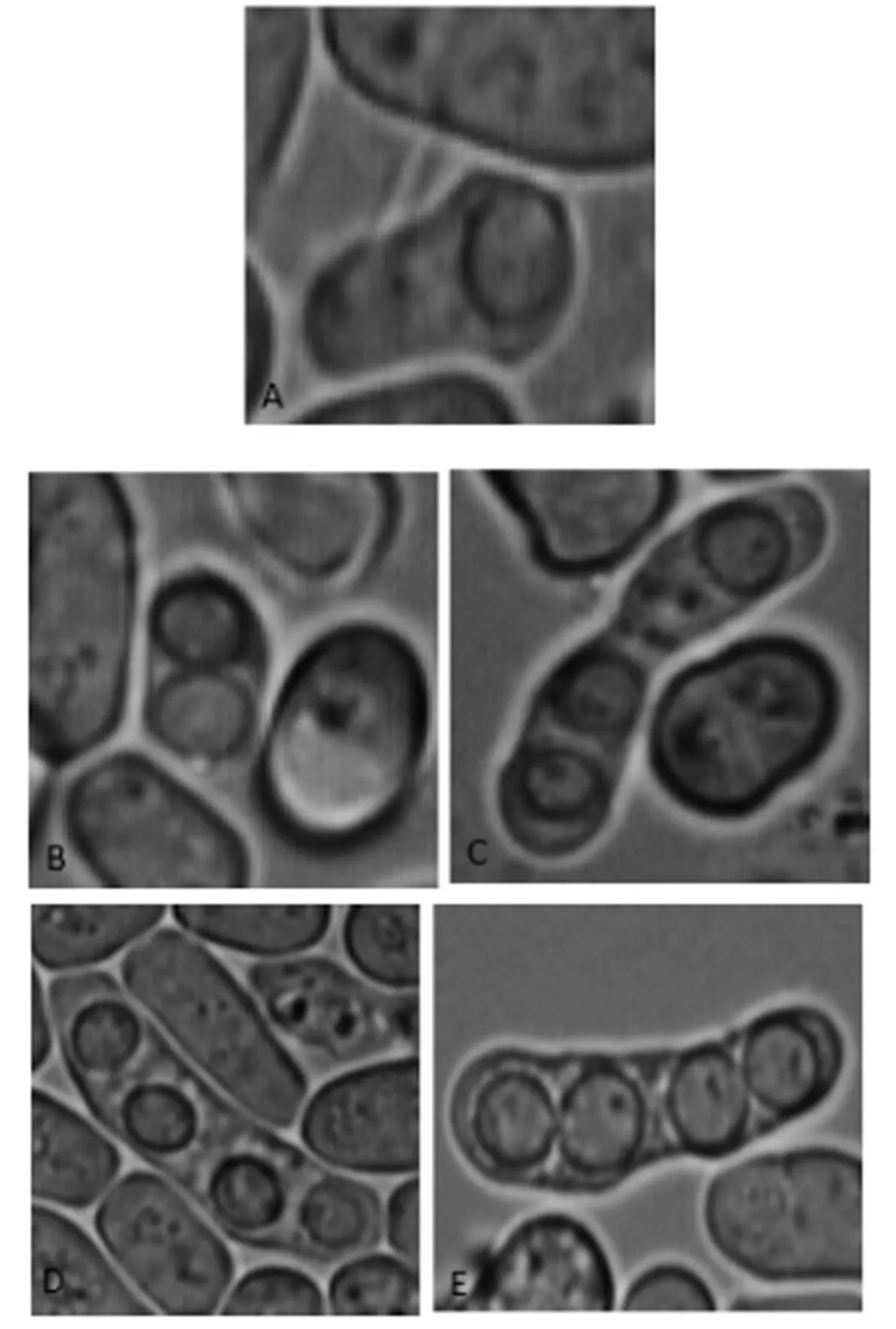
圖4 野生型與突變株的孢子形態(tài)圖(100×10)
注:A、B、C、D為突變株108的孢子;E為野生型的孢子
3 結論
本研究將裂殖酵母野生型的SPAC1006.01基因敲除,通過觀察生長曲線,發(fā)現(xiàn)12 h后野生型在37 ℃的OD595比25 ℃時更高;突變株108在25 ℃和37 ℃的OD595基本沒有差別,但都比野生型低。該實驗結果表明,基因敲除不僅會影響菌株的生長速度,導致突變株生長緩慢,同時導致突變株對溫度不敏感。在觀察子囊孢子的數(shù)目和形態(tài)實驗中,發(fā)現(xiàn)產孢菌株中有10%的菌產生1個孢子、30%產生2個孢子、15%產生3個孢子,只有45%的菌能夠正常產生4個孢子,但孢子的形態(tài)沒有差異。本實驗結果為進一步深入研究SPAC1006.01基因影響裂殖酵母生長和產孢的具體分子機制提供了理論依據(jù)和技術支持。
[1]Ladds G,Davey J.Identification of proteases with shared functions to the proprotein processing protease Krp1 in the fission yeast Schizosaccharomyces pombe[J].Mol Microbiol,2000(4).
[2]Mukaiyama H,Iwaki T,Idiris A,et al.Processing and maturation of carboxypeptidase Y and alkaline phosphatase in Schizosaccharomyces pombe[J].Applied Microbiology & Biotechnology,2011,90(1):203-213.
[3]Nú?ez Andrés,Dominic D,Mehdi J,et al.Calnexin Is Essential for Survival under Nitrogen Starvation and Stationary Phase in Schizosaccharomyces pombe[J].PLoS one,2015,10(3).
[4]Kojiro T,Ayaka M,Mitsuhiro Y,et al.Identification of Genes Affecting the Toxicity of Anti-Cancer Drug Bortezomib by Genome-Wide Screening in S.pombe[J].PLoS one,2011,6(7).
[5]王文超.裂殖酵母表現(xiàn)遺傳調控產孢及異染色質沉默的研究[D].上海:復旦大學,2014.
[6]成少寧,許先猛,馬欣,等.一株優(yōu)良果脯加工廢糖液酒精發(fā)酵酵母菌生長條件的研究[J].漯河職業(yè)技術學院學報,2016,15(2):12-15.
2095-6835(2019)01-0001-03
Q933
A
10.15913/j.cnki.kjycx.2019.01.001
〔編輯:嚴麗琴〕
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
