家兔β干擾素啟動子質粒的構建及其啟動子活性鑒定
2019-03-03 02:43:42陳萌萌范志宇胡波宋艷華魏后軍仇汝龍朱偉峰徐為中王芳
江蘇農業科學 2019年23期
陳萌萌 范志宇 胡波 宋艷華 魏后軍 仇汝龍 朱偉峰 徐為中 王芳


摘要:通過家兔體內感染兔出血癥病毒(RHDV)后發現體內干擾素-β(Interferon-β,IFN-β)基因表達水平明顯降低。為體外研究RHDV感染宿主后IFN-β啟動子的調控機制,首先以家兔肝細胞總cDNA為模板,利用PCR擴增IFN-β基因啟動子區的序列,經測序確定全長為550 bp;預測軟件分析,與人源IFN-β基因啟動子的轉錄上調區域(positive regulation domain,PRD)高度同源;隨后將啟動子全長和一系列轉錄元件克隆至pGL6-Basic熒光素酶表達載體中,成功構建重組體報告基因質粒后,瞬轉RK-13細胞,利用試劑盒檢測熒光素酶的相對活性。結果顯示,克隆的IFN-β基因啟動子區和AP-1、IRF3、NF-κB和IRF7等轉錄元件具有顯著的活性,為研究兔出血癥病毒對宿主 IFN-β 基因的轉錄調控機制提供了有效的工具。
關鍵詞:家兔;兔出血癥病毒;IFN-β基因;啟動子;熒光素酶;轉錄調控
中圖分類號: S858.291 文獻標志碼: A 文章編號:1002-1302(2019)23-0078-03
干擾素是一類在機體內廣泛存在的糖蛋白,可分為Ⅰ型(IFN-α、IFN-β和ω)和Ⅱ型(IFN-γ)干擾素[1-2]。干擾素-β (interferon-β,IFN-β)是由干擾素誘生劑(微生物和病毒等)刺激生物成纖維細胞和白細胞等產生的高活性多功能蛋白,具有多種生物活性,如抗病毒和免疫調節[1-4]等。近來的研究發現,多種病毒感染機體后能夠采取不同的措施顯著改變IFN-β的表達水平,從而逃避早期由IFN-β介導的抗病毒天然免疫反應,如在豬偽狂犬病毒[5]、圓環病毒[6]和豬繁殖與呼吸綜合征病毒[7]的感染導致的疾病中。目前,對兔天然免疫系統了解很少,而且缺乏有效的檢測天然免疫分子的工具,這嚴重阻礙了病毒與兔宿主相互作用的深入研究。
筆者發現RHDV感染家兔能夠顯著下調兔體內IFN-β基因的轉錄水平,推測IFN-β基因水平的降低可能與該病毒的致病性相關。為了分析兔源IFN-β基因的轉錄調控,筆者所在課題組以家兔IFN-β為研究對象,通過制備一系列含有IFN-β基因啟動子和轉錄元件的重組體報告基因質粒,采用雙報告基因系統鑒定IFN-β和轉錄元件的轉錄活性,為深入揭示啟動子功能研究奠定了基礎,同時也為研究RHDV調控IFN-β基因的表達提供有效的工具。
1 材料與方法
1.1 試驗材料
兔腎RK-13細胞株由筆者所在實驗室保存。DMEM培養基購自美國Hyclone公司。胎牛血清(FBS)和青鏈霉素溶液均購自美國GIBCO BRL公司。Lip3000脂質體以及無血清培養基均購自美國英杰生命技術有限公司。真核表達載體pcDNA3.1-6his購自南京金斯瑞生物科技有限公司。pGL6-Basic螢火蟲熒光素酶報告載體以及pTL-TK海參熒光素酶的內參載體購自上海碧云天生物技術有限公司。
1.2 試驗方法
1.2.1 RHDV攻毒 選用3月齡健康易感新西蘭兔,分攻毒組和對照組,每組5只。攻毒用毒種為兔出血癥病毒皖阜株肝毒,由筆者所在實驗室保存。1 mL病毒(含量 105 LD50/mL,LD50為半數致死劑量)為攻毒劑量;等量的PBS(磷酸緩沖鹽溶液)作為對照。
1.2.2 IFN-β和病毒基因相對表達量的檢測 采用Primer 5.0軟件設計針對兔IFN-β基因啟動子和RHDV基因的引物(表1),送上海英駿生物技術有限公司合成。稱取 0.5 g 肝臟組織勻漿提取RNA,反轉錄cDNA,以此為模板進行熒光定量PCR檢測。體系:SYBR Mix 10 μL,PCR上下游引物(10 μmol/L)各0.4 μL,DNA模板2 μL,ddH2O 7.2 μL,共 20 μL。程序:95 ℃預變性30 s;95 ℃ 5 s,55 ℃ 30 s,循環40次。采用2-ΔΔCT法進行數據分析,計算基因相對表達水平。
1.2.3 IFN-β基因啟動子的克隆 采用苯酚-氯仿法提取健康兔肝臟組織DNA,用超微量核酸分析儀檢測DNA的完整性、純度和濃度。以IFNb-F:5′-ATGATCAACAGGTGTATCCTTCAA-3′和IFNb-R:5′-TTAGAGGTAATCTATGAGCATGATGAT-3′為引物,以兔肝組織基因組DNA為模板,PCR擴增兔IFN-β啟動子區550 bp序列,體系:50 μL反應體系含模板2 ng,上、下游引物(10 μmol)各1 μL,其他試劑按使用說明加入。程序:95 ℃預變性5 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 30 s,35個循環;72 ℃ 10 min。PCR產物經1%瓊脂糖凝膠電泳并回收目的片段,與pMD18-T載體4 ℃連接過夜,轉化DH5α感受態細胞,挑取陽性菌,液體LB培養基中過夜培養后,提取質粒,送上海英駿生物技術有限公司測序。
1.2.4 IFN-β基因啟動子序列的生物信息學 利用NCBI中的Blast同源性比對兔和人IFN-β基因啟動子的轉錄上調區域PRD序列;通過Promoter 2.0 Prediction軟件分析 IFN-β 基因的啟動子序列。
1.2.5 IFN-β啟動子和順式作用元件報告基因重組體的構建 以重組IFN-β啟動子載體為模板,采用PCR方法克隆IFN-β基因啟動子,雙酶切后,經1%瓊脂糖凝膠電泳后回收酶切片段后連接到pGL6-Basic載體內,構建含IFN-β基因啟動子序列的重組質粒pIFN-β-Luc并進一步雙酶切和測序鑒定。另外,由公司合成了IRF-3和NF-κB結合位點序列,并構建了重組質粒pIRF-3-Luc和pNF-κB-Luc。
1.2.6 質粒轉染 在試驗前24 h,將兔腎細胞RK-13接種于24孔板,待細胞達80%融合度時進行轉染。采用Invitrogen公司轉染試劑lip3000按試劑說明書操作,轉染 RK-13 細胞。試驗設轉染pGL6-Basic載體組(對照組)和pGL6-IFN-β啟動子等陽性載體組(試驗組),每組3個重復,在24孔板中共轉染內參質粒(pRL-TK)與報告質粒,轉染24 h后收取細胞。
1.2.7 熒光素酶活性測定 在轉染24 h后,棄去培養液,用PBS液洗3次。每孔加入100 μL細胞裂解液,充分裂解細胞20 min后,取25 μL裂解液加入96孔白板,然后按照雙熒光素酶檢測試劑盒上的說明書操作,在微孔板發光分析儀上檢測螢火蟲熒光素酶活性(pGL6)和海參熒光素酶活性(pRL-TK)。以螢火蟲和海參熒光素酶活性的比值即為相對熒光強度(relative luciferase activity,簡稱RAL)來指示試驗各組的轉錄活性。計算各組熒光素酶的相對熒光強度RAL。
1.3 數據分析
數據分析采用SPSS 13.0統計學軟件,運用統計學分析方法t檢驗(α=0.05),數據以“平均值±標準差”表示。
2 結果與分析
2.1 肝臟組織病毒和IFN-β mRNA水平測定
Real-time PCR檢測結果表明,與對照組相比,兔出血癥病毒感染兔體肝臟組織中病毒基因相對表達量快速上升(圖1-A);同時,發現在病毒感染48 h以后IFN-β mRNA水平明顯降低(圖1-B)。
2.2 PCR擴增IFN-β基因啟動子序列
設計合成引物以兔肝組織提取的DNA為模板擴增兔IFN-β基因5′UTR區,PCR擴增出約550 bp特異性目的片段(圖2),將擴增產物克隆至pMD18-T載體中,篩選陽性質粒后經雙酶切鑒定,最后進一步進行測序。測序表明,PCR擴增兔IFN-β基因啟動子序列長度為550 bp。
2.3 IFN-β基因啟動子及轉錄因子結合位點分析
在線預測軟件分析表明,IFN-β基因啟動子的ISRE包含4個轉錄上調區域(positive regulation domain,PRD)分別是Ⅰ~Ⅳ區(圖3-A);此序列包括轉錄起始位點(+1)上游500 bp,具有典型的真核生物啟動IFN-β基因啟動子具有AP-1、IRF3、IRF7、NF-κB和TATA-box等典型的真核生物啟動子元件[10](圖3-A)。Blast序列比對發現,擴增得到的IFN-β基因PRD與人源IFN-β基因啟動子PRD序列相似性達98%以上(圖3-B)。
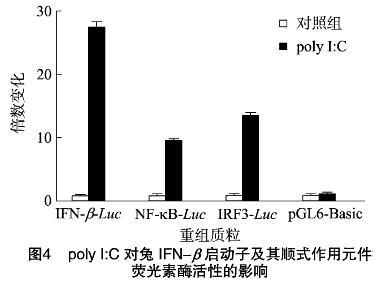
2.4 重組報告質粒的的構建和活性鑒定
以攜帶有IFN-β啟動子序列的pMD18-T載體為模板,PCR擴增啟動子序列,經雙酶切后與酶切的pGL6-Basic載體4 ℃連接,成功構建啟動子報告基因載體(pIFN-β-Luc),另外構建轉錄因子結合元件的熒光素酶報告基因載體(pIRF3-Luc和pNF-κB-Luc),經測序正確。
將構建好的重組質粒pIFN-β-Luc、pIRF3-Luc和 pNF-κB-Luc分別轉染至RK-13細胞中,同時轉染內參質粒(pRL-TK),通過檢測熒光素酶的相對活性來反映啟動子和順式作用元件的活性,分別設陰性和陽性polyI:C處理組。熒光素酶活性分析表明,與轉染pGL6-Basic對照組相比,polyI:C能明顯誘導兔IFN-β基因啟動子和順式作用元件IRF3和NF-κB相對熒光素酶活性(圖4), 由此說明成功構建了報告基因重組體。
3 討論
Ⅰ型干擾素是由病毒直接刺激被感染細胞所轉錄產生的抵御病毒感染的第一防線,為了逃避宿主的免疫反應,人類乳頭狀病毒[5]、EB病毒[8]、埃博拉病毒[9]、丙肝病毒[10]等病毒均可通過其編碼的病毒蛋白抑制干擾素順式作用元件的活性從而抑制IFN-β的表達。本研究發現,在兔體感染兔出血癥病毒后,會顯著下調IFN-β基因的表達量,這也許與兔出血癥病毒的致病性密切相關。然而目前對兔天然免疫系統的研究很少,缺乏有效的研究工具。
本研究成功獲得了IFN-β的轉錄起始位點上游550 bp啟動子序列,將兔IFN-β啟動子及IRF3等順式作用元件亞克隆進pGL6-Basic中,通過熒光素酶的表達活性來間接表明兔IFN-β基因啟動子和順式作用元件在RK13細胞中的活性,為研究IFN-β轉錄起始位點上游啟動子的生物學活性提供了依據。預測軟件分析出IFN-β的啟動子含AP-1、IRF3、IRF7和NF-κB等多個轉錄因子結合序列,暗示IFN-β基因的轉錄受到多個轉錄因子的調控。同時,同源性分析發現兔IFN-β基因啟動子與人源IFN-β基因啟動子[11]序列高度相似,尤其是IRF3和NF-κB轉錄調控元件,這進一步說明本試驗克隆序列的正確性。然而,具體哪些轉錄因子影響IFN-β的轉錄活性,還需要對其啟動子區進行序列的缺失等試驗來證實[12]。總之,本研究構建了IFN-β啟動子和順式作用元件的重組報告質粒,并分析了IFN-β啟動子區域含有的AP-1、IRF3、IRF7和NF-κB等多個轉錄因子結合序列,為研究IFN-β轉錄調控以及其啟動子功能提供了有效工具,也為進一步探究兔出血癥病毒的致病機制奠定了基礎。
參考文獻:
[1]Bogdan C. The function of type I interferons in antimicrobial immunity[J]. Current Opinion in Immunology,2000,12(4):419-424.
[2]Lazear H M,Schoggins J W,Diamond M S. Shared and distinct
functions of type Ⅰ and type Ⅲ interferons[J]. Immunity,2019,50(4):907-923.
[3]Zinzula L,Tramontano E. Strategies of highly pathogenic RNA viruses to block dsRNA detection by RIG-I-like receptors:hide,mask,hit[J]. Antiviral Research,2013,100(3):615-635.
[4]Uehara J,Ohkuri T,Kosaka A,et al. Intratumoral injection of IFN-β induces chemokine production in melanoma and augments the therapeutic efficacy of anti-PD-L1 mAb[J]. Biochemical and Biophysical Research Communications,2017,490(2):521-527.
[5]Wei J Y,Ma Y M,Wang L,et al. Alpha/beta interferon receptor deficiency in mice significantly enhances susceptibility of the animals to pseudorabies virus infection[J]. Veterinary Microbiology,2017,203:234-244.
[6]Huang B,Zhang L L,Lu M Q,et al. PCV2 infection activates the cGAS/Sting signaling pathway to promote IFN-β production and viral replication in PK-15 cells[J]. Veterinary Microbiology,2018,227:34-40.
[7]Luo R,Xiao S,Jiang Y,et al. Porcine reproductive and respiratory syndrome virus(PRRSV)suppresses interferon-β production by interfering with the RIG-I signaling pathway[J]. Molecular Immunology,2008,45(10):2839-2846.
[8]Bentz G L,Liu R,Hahn A M,et al. Epstein-Barr virus BRLF1 inhibits transcription of IRF3 and IRF7 and suppresses induction of interferon-β[J]. Virology,2010,402(1):121-128.
[9]Gupta M,Mahanty S,Ahmed R,et al. Monocyte-Derived human macrophages and peripheral blood mononuclear cells infected with Ebola virus secrete MIP-1α and TNF-α and inhibit Poly-IC-induced IFN-α in vitro[J]. Virology,2001,284(1):20-25.
[10]Abe H,Imamura M,Hiraga N,et al. ME3738 enhances the effect of interferon and inhibits hepatitis C virus replication both in vitro and in vivo[J]. Journal of Hepatology,2011,55(1):11-18.
[11]Ford E,Thanos D. The transcriptional code of human IFN-β gene expression[J]. Biochimica et Biophysica Acta-Gene Regulatory Mechanisms,2010,1799(3/4):328-336.
[12]王艷艷,金 蕊,徐華國,等. 干擾素基因刺激因子啟動子的克隆、鑒定及活性分析[J]. 江蘇醫藥,2015,41(13):1489-1491.
收稿日期:2019-09-19
基金項目:國家自然科學基金(編號:31702274);現代農業產業技術體系建設專項資金(編號:CARS-43-C-1)。
作者簡介:陳萌萌(1987—),女,江蘇連云港人,博士,助理研究員,主要從事家兔疾病防治與獸醫生物技術研究。E-mail:moonchen2010@yeah.net。
通信作者:王 芳,博士,研究員,主要從事畜禽疫病防控及免疫機理研究。E-mail:rwangfang@126.com。
