馬鈴薯StPR-1b蛋白的原核表達及抑菌活性
2019-03-03 02:43:42盧美歡李利軍馬英輝
江蘇農業科學 2019年23期
關鍵詞:融合
盧美歡 李利軍 馬英輝


摘要:為探索馬鈴薯病害生物防治和抗病育種的有效途徑,構建馬鈴薯病程相關蛋白StPR-1b的pET30A原核表達體系,用鎳柱分離純化該融合蛋白,結果表明,經0.5 mmol/L 異丙基硫代-β-D-半乳糖苷誘導后,StPR-1b蛋白在大腸桿菌中得到了高效表達;純化得到預期大小(約15 ku)的融合蛋白;StPR-1b蛋白對馬鈴薯軟腐病病原菌胡蘿卜軟腐果膠桿菌有一定的抑制作用,蛋白濃度越大,抑制效果越強。
關鍵詞:StPR-1b蛋白;原核表達;馬鈴薯;抑菌作用;胡蘿卜軟腐果膠桿菌
中圖分類號: S435.32 文獻標志碼: A 文章編號:1002-1302(2019)23-0085-03
病程相關蛋白(pathogenesis-related proteins,簡稱PRP或PRs)是指植物在病理或病理相關的環境下誘導產生的,普遍存在的具有廣譜抗性的誘導的可溶性蛋白質,是植物產生誘導抗病性的一種生化機制。PRP相對分子質量為10~40 ku,積累于病原侵染寄主植物部位和未侵染部位的細胞內和細胞間,主要通過固化寄主細胞壁、提高抗菌活性和參與細胞信號轉化等方式參與植物的誘導抗病性[1-2],能夠抵抗蛋白酶、糖苷酶、重金屬、尿素、低pH值和高溫(60 ℃)等逆境環境,具有較強的穩定性[3-6]。大多數PRP具有幾丁質酶和β-1,3-葡聚糖酶活性,在植物體內和體外顯示出抗真菌活性,與植物過敏性壞死反應(hypersensitive response,簡稱HR)和系統獲得性抗性(systematic acquired resistance,簡稱SAR)有密切關系,PRP在寄主中的誘導表達常作為SAR建立的標志[7],因此人們可通過構建PRP基因的方法獲得抗菌譜廣、作用持久的抗病新品種。
馬鈴薯軟腐病是馬鈴薯產區發生最嚴重的細菌性病害,在苗期和窖藏期容易發生,馬鈴薯是我國重要的經濟作物和蔬菜品種,營養豐富,產量很高,但目前我國多數馬鈴薯主產區都受到了馬鈴薯軟腐病的嚴重威脅,對我國馬鈴薯的生產造成極大的破壞,2000年福建省福鼎市馬鈴薯軟腐病發生面積為25107 hm2,是總種植面積的82%,造成了嚴重的經濟損失[8]。
本研究對病程相關蛋白StPR-1b 進行原核表達,通過構建原核表達系統將融合蛋白大量表達和分離純化,探索該蛋白對馬鈴薯軟腐病病原菌的抑制作用,以期開發安全無害的抗生素,并為該基因在馬鈴薯抗病育種中的應用提供理論基礎。
1 材料與方法
1.1 試驗材料
1.1.1 生物材料 馬鈴薯軟腐病病原菌、表達載體pET30a載體、pET30A-StPR-1b質粒,均為筆者所在實驗室保存;表達宿主為大腸桿菌感受態細胞,購自Invitrogen公司。
1.1.2 酶和試劑 T4連接酶、DNA Marker、NdeⅠ和XhoⅠ內切酶,均購自TaKaRa公司;凝膠回收試劑盒,購自生工生物工程(上海)股份有限公司;異丙基硫代-β-D-半乳糖苷(IPTG),購自Sigma公司;PVDF膜,購自Millpore公司;其他試劑均為國產分析純。
1.2 試驗方法
1.2.1 引物設計 根據StPR-1b基因序列,利用引物設計軟件Rrimer 5.0設計合成引物,引物序列:上游5′-GACACGACACCATATGCAGAACTCTCCGCAGGATTAT-3′,下游5′-GTGTCCTCGAGGTACGGACGCTGACCAACCCAG-3′,以含StPR-1b全長的pET30A-StPR-1b質粒為模板進行聚合酶鏈式反應(polymerase chain reaction,PCR)擴增。
1.2.2 PCR反應條件 以pET30A-StPR-1b質粒為模板,在總體積為50 μL的EP管中依次加入1 μL dNTP(25 mmol/L)、5 μL高溫聚合酶(Pfu) Buffer(10×)、2 μL上游引物、2 μL下游引物、0.4 μL Pfu(5 U/μL),用ddH2O補至50 μL。擴增程序:95 ℃預變性3 min,95 ℃變性22 s,53 ℃退火20 s,72 ℃ 延伸60 s,24個循環,72 ℃修復延伸5 min。
1.2.3 PCR電泳及回收 將PCR產物用2%瓊脂糖凝膠在1×TAE緩沖液,電壓為150 V,電流為100 mA條件下,20 min電泳觀察。將目的條帶切下,按試劑盒回收StPR-1b片段,回收純化好的片段備用酶切。酶切體系為1 μg純化回收好的片段、5 μL酶切Buffer(10×)、1 μL限制性內切酶NdeⅠ(10 U/μL)和1 μL限制性內切酶XhoⅠ(10 U/μL),用ddH2O補至50 μL,在溫度為37 ℃恒溫水浴鍋中反應2 h。
1.2.4 目的片段與載體連接、轉化和篩選 回收純化好的目的DNA片段8 μL、T4連接酶Buffer 2 μL、pET30A載體4 μL、T4連接酶1 μL,用ddH2O補至20 μL,在16 ℃條件下反應 4 h;然后將其轉入oneshort感受態細胞中,檢測篩選出陽性克隆送生工生物工程(上海)股份有限公司進行測序。測序正確的菌液使用質粒提取試劑盒SK8191 SanPrep柱式質粒DNA小量抽提試劑盒提取質粒DNA。
1.2.5 原核表達載體的構建 用NdeⅠ和XhoⅠ內切酶對測序正確的pET30A-StPR-1b質粒和表達載體pET30A分別進行雙酶切,切膠回收目的片段用T4 DNA連接酶連接到pET30a(+)載體上,將連接產物轉化到oneshort感受態細胞中,提取轉化子的質粒DNA,經PCR檢測和酶切檢測證實含目的基因的片段已經插入原核表達載體中并成功轉入表達宿主菌中,得到重組表達載體。
1.2.6 融合蛋白的誘導表達及純化 挑取表達菌株的單菌落于含卡那霉素的LB培養基中,在37 ℃、220 r/min下過夜培養。然后按1 ∶ 100稀釋培養液,在37 ℃、220 r/min下培養至D600 nm為0.6時,添加終濃度為 0.5 mmol/L 的IPTG,在 220 r/min,溫度分別為20、37 ℃條件下誘導過夜;37 ℃誘導 4 h,以未加IPTG誘導劑的處理作為陰性對照。4 000 r/min離心10 min收集菌體,棄上清,菌體用500 μL PBS(pH值為7.4)緩沖液懸浮,添加終濃度為 0.5 mmol/L 的溶菌酶,超聲破碎6 min,破碎0.5 s停1.5 s,分別離心收集上清和沉淀,沉淀用500 μL包涵體溶解液(8 mol/L 尿素,50 mmol/L Tris-HCl,300 mmol/L NaCl,pH值為8.0)溶解,分別取40 μL樣品和10 μL 5×protein loading buffer混勻,沸水浴10 min,離心后取上清,進行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(簡稱SDS-PAGE)檢測。準備12%的SDS-PAGE,Tris-Gly電泳緩沖液(Tris 3.0 g,甘氨酸14.4 g,SDS 1.0 g,定容至1 L),上樣量為10 μL,濃縮膠條件:80 V 20 min,分離膠條件:120 V 60 min,凝膠電泳結束用考馬斯亮藍染色20 min,脫色后觀察蛋白條帶。
將培養的菌液按1 ∶ 100比例接種于3 L含有卡那霉素的LB液體培養基中,在37 ℃、220 r/min條件下培養,當D600 nm達到 0.6 時,添加終濃度為0.5 mmol/L的IPTG,在 20 ℃、220 r/min條件下誘導過夜,離心收集細胞菌體,采用鎳瓊脂糖親和層析進行蛋白純化后,-80 ℃保存。
1.2.7 蛋白免疫印記試驗(Western Blot) 將純化好的 HIS-StPR-1b蛋白進行Western Blot檢測,蛋白經過SDS-PAGE轉移至PVDF膜后,用5%的脫脂奶粉,在37 ℃條件下緩慢振蕩2 h,加兔抗his標簽的一抗(1 ∶ 500稀釋)在37 ℃條件下緩慢振蕩 60 min。加羊抗兔的二抗孵育60 min,最后用3,3′,5,5′-四甲基聯苯胺(TMB)顯色鑒定。
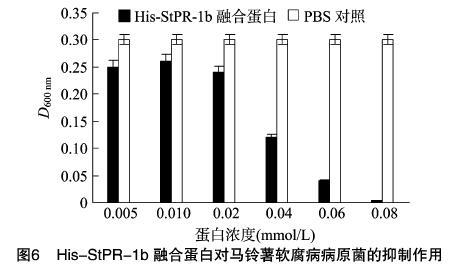
1.2.8 純化的HIS-StPR-1b蛋白對馬鈴薯軟腐病病原菌的抑制試驗 將HIS-StPR-1b蛋白用無菌雙蒸水配制成 1 mg/mL 蛋白溶液,再稀釋成不同濃度梯度備用。將保存的馬鈴薯軟腐病病原菌胡蘿卜軟腐果膠桿菌活化,在LB液體培養基中培養24 h。在已滅菌的EP離心管中加入880 μL液體LB培養基,20 μL軟腐病病原菌菌液,同時加入稀釋好的HIS-StPR-1b蛋白溶液100 μL,32 ℃振蕩培養12 h,每個濃度做3次重復,測定D600 nm。同時做空白對照。
2 結果與分析
2.1 StPR-1b原核表達載體的構建
以pET30a-StPR-1b質粒為模板進行PCR擴增,擴增出與預期大小一致的417 bp條帶(圖1),對目標條帶進行瓊脂糖電泳、回收。回收產物亞克隆于pET30a載體中。對含有StPR-1b基因的重組pET30a質粒進行NdeⅠ和XhoⅠ雙酶切,PCR檢測和酶切鑒定結果顯示,StPR-1b基因已成功導入到原核表達載體pET30a中,目標片段大小與該基因編碼區大小一致。
2.2 StPR-1b蛋白的誘導表達
將原核表達載體pET30a-StPR-1b轉化至大腸桿菌oneshort感受態細胞,在IPTG誘導下StPR-1b能成功表達,菌體中檢測到高表達量的蛋白條帶(圖2的泳道3和泳道5)。考察不同溫度誘導StPR-1b蛋白表達的影響,結果顯示,在20、37 ℃ 2個溫度處理中,37 ℃誘導更有利于StPR-1b蛋白的表達。表明不同誘導溫度對StPR-1b蛋白表達的影響較明顯。
2.3 純化蛋白SDS-PAGE分析
采用鎳瓊脂糖親和層析對在大腸桿菌中大量表達的重組StPR-1b蛋白進行純化,采用不同濃度的咪唑基(Imidazole)洗脫液洗脫,發現20 mmol/L Imidazole洗脫組分能較好地將StPR-1b蛋白分離純化(圖3),收集蛋白洗脫液冷凍干燥,經檢測蛋白濃度為84%。StPR-1b融合蛋白經過純化,SDS-PAGE 電泳分析在約15 ku出現明顯條帶,表明融合蛋白成功得到了純化(圖4)。
2.4 StPR-1b融合蛋白的Western Blot分析
為了進一步確定StPR-1b融合蛋白是否在大腸桿菌中成功表達,用抗his標簽的一抗和加羊抗兔的二抗進行Western Blot檢測,用TMB顯色鑒定。結果表明,在預期的 15 ku 位置有特異性條帶出現(圖5),證明StPR-1b融合蛋白表達成功。
2.5 StPR-1b蛋白對馬鈴薯軟腐病病原菌的抑制作用
配制不同濃度StPR-1b融合蛋白溶液,研究其對軟腐病病原菌的抑制作用。結果(圖6)顯示,在0.005~0.08 mmol/L蛋白溶液濃度范圍內,His-StPR-1b融合蛋白對馬鈴薯軟腐病病原菌胡蘿卜軟腐果膠桿菌有一定的抑制作用,蛋白濃度越大,抑制效果越強,當His-StPR-1b融合蛋白濃度為 0.08 mmol/L 時,抑制率達到90.0%。
3 結論
馬鈴薯軟腐病在世界范圍內廣泛發生,是馬鈴薯產區發生最嚴重的細菌性病害,嚴重影響馬鈴薯產量。積極培育抗病品種,是緩解馬鈴薯軟腐病的有效途徑。本研究通過構建原核表達載體,將StPR-1b基因成功在大腸桿菌中得到表達。在原核表達系統中,不同誘導條件對StPR-1b蛋白表達的影響較明顯,本研究經多次誘導及SDS-PAGE分析后發現,StPR-1b蛋白最佳誘導溫度為37 ℃,采用鎳親和層析進行蛋白純化,最佳純化條件為20 mmol/L Imidazole洗脫組分,獲得了純度為84%的蛋白,這為進一步研究StPR-1b蛋白的生物活性及在馬鈴薯抗病方面的生物學功能奠定了基礎。
植物在受到真菌、細菌和病毒等病原體入侵時合成抗毒素、抗菌肽或一些病程相關蛋白,PRs與寄主植物抗病性和系統獲得性具有密切關系,是植物自我抗御機制中的可誘導組分,目前已知的蛋白根據序列同源性、血清學關系和生物活性被劃分為17個家族。抗病防衛反應中表達的基因主要是編碼病程相關蛋白和植保素合成相關酶基因,而PRs在植物抗病性和系統獲得抗性中起重要作用,是因為不同PRs基因的表達水平和抗病性之間緊密相關。StPR-1b蛋白屬于 PR-1 家族成員,StPR-1b基因編碼1個含有159個氨基酸的蛋白質,在N端存在長為24個氨基酸的信號肽。本研究通過PCR技術擴增了馬鈴薯病程相關蛋白 StPR-1b基因,并使克隆的StPR-1b基因在大腸桿菌中得到成功表達,優化表達體系為純化產物驗證體外生物活性及后期培育抗病轉基因馬鈴薯提供了參考。
參考文獻:
[1]Loon L V,Strien E V. The families of pathogenesis-related proteins,their activities,and comparative analysis of PR-1 type proteins[J]. Physiological & Molecular Plant Pathology,1999,55(2):85-97.
[2]Christensen A B,Cho B H,Nsby M,et al. The molecular characterization of two barley proteins establishes the novel PR-17 family of pathogenesis-related proteins[J]. Molecular Plant Pathology,2002,3(3):135-144.
[3]王勇剛,曾富華,吳志華,等. 植物誘導抗病與病程相關蛋白[J]. 湖南農業大學學報(自然科學版),2002,28(2):177-182.
[4]Hoffmann-Sommergruber K. Pathogenesis-related (PR)-proteins identified as allergens[J]. Biochemical Society Transactions,2002,30(6):930-935.
[5]Bertini L,Leonardi L,Caporale C,et al. Pathogen-responsive wheat PR4 genes are induced by activators of systemic acquired resistance and wounding[J]. Plant Science,2003,164(6):1067-1078.
[6]Ziadi S,Poupard P,Brisset M N,et al. Characterization in apple leaves of two subclasses of PR-10 transcripts inducible by acibenzolar-S-methyl,a functional analogue of salicylic acid[J]. Physiological and Molecular Plant Pathology,2001,59(1):33-43.
[7]范志金,劉秀峰,劉鳳麗,等. 植物抗病激活劑誘導植物抗病性的研究進展[J]. 植物保護學報,2005,32(1):87-92.
[8]馬 宏. 我國馬鈴薯軟腐病防治的研究進展[J]. 生物技術通報,2007(1):42-44.
收稿日期:2018-10-31
基金項目:陜西省科學院科技計劃(編號:2015K-05);陜西省科技廳農業攻關計劃(編號:2014K02-13-02、2016NY-197);西安市科技局農業創新計劃[編號:2017050NC/NY009(4)]。
作者簡介:盧美歡(1981—),女,廣東羅定人,碩士,副研究員,主要從事植物病害生物防治研究。E-mail:lu_meihuan@sina.com。
通信作者:李利軍,研究員,主要從事微生物物資開發與利用研究。E-mail:lijun_lli@163.com。
猜你喜歡
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
數學年刊A輯(中文版)(2022年4期)2022-02-16 08:17:34
今日農業(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
無線電通信技術(2021年4期)2021-07-13 08:58:28
無線電通信技術(2021年3期)2021-06-08 03:33:48
中學生數理化(高中版.高考數學)(2021年1期)2021-03-19 08:28:38
無線電工程(2020年11期)2020-10-29 01:25:46
現代出版(2020年3期)2020-06-20 07:10:34
福利中國(2015年4期)2015-01-03 08:03:38
