魚膠原蛋白肽對高脂膳食小鼠肝臟脂肪代謝和氧化還原狀態的影響
2019-03-08 08:50:22楊玉輝王雅楠張佳紅郭海濤施用暉樂國偉
食品科學 2019年3期
田 許,楊玉輝,王雅楠,張佳紅,郭海濤,施用暉,樂國偉,*
(1.江南大學 食品科學與技術國家重點實驗室,江蘇 無錫 214122;2.江南大學食品學院,江蘇 無錫 214122)
近年來隨著物質生活水平的不斷提高,我國居民從傳統的植物性食物為主的膳食結構向高脂肪、低碳水化合物的膳食結構發展,導致肥胖的發生率逐年增加[1-2]。當前肥胖已經成為全球性的公共衛生問題而被廣泛關注[3]。許多研究已經證明了肥胖是代謝綜合征發生最常見的危險因素,而且還是糖尿病、心腦血管疾病和多種癌癥發生的重要誘因[4]。長期高脂膳食(high-fat diet,HFD)會使機體產生過量的自由基,破壞氧化還原狀態的平衡,繼而導致脂代謝的紊亂[5],最終誘使肥胖發生。已有大量研究表明通過飲食中添加抗氧化因子能夠對氧化還原狀態失衡導致的脂代謝紊亂起到良好的改善作用[6-9]。
膠原蛋白肽已被證實具有降血壓、抗氧化、抗衰老、調節腸道微生態、抗腫瘤、改善骨骼和皮膚以及免疫調節等多種活性[10-14]。近年來,陸源性膠原蛋白的開發利用遇到宗教和安全性方面的諸多問題,水產魚類加工利用后的下腳料中因富含膠原蛋白而受到研究者們的重視[15]。魚膠原蛋白肽(fish collagen peptides,FCPs)是從魚類加工后的皮、骨、鱗和鰭等廢棄物中提取膠原蛋白后經過酶法處理所得[12]。過去的研究已經發現豬骨膠原蛋白肽具有改善HFD小鼠抗氧化和血脂代謝的作用[16],丁進鋒等[17]使用海蜇膠原蛋白肽也觀察到了相似的結果,但具體的作用機制還缺乏深入研究。肝臟作為機體脂類消化、吸收、合成、分解和運輸的重要場所[18],是很多營養功能因子的作用位點。因此,本實驗以HFD C57BL/6小鼠為實驗動物模型,研究FCPs干預對HFD小鼠肝臟脂代謝和氧化還原狀態的影響,為FCPs功能的開發和利用提供實驗依據和理論支持。
1 材料與方法
1.1 動物、材料與試劑
4 周齡清潔級C57BL/6雄性小鼠54 只(18~20 g)飼養于江南大學動物實驗中心,購自南京大學生物醫藥研究院。實驗動物合格證號:SCXK(蘇)2015-0001。
FCPs(食品級,純度99%,分子質量150~2 000 Da) 韓國Geltech公司;總抗氧化能力(total antioxidant capacity,T-AOC)、谷胱甘肽過氧化物酶(glutathion peroxidase,GSH-Px)、丙二醛(malondialdehyde,MDA)、還原型谷胱甘肽(reduced glutathione,GSH)、氧化型谷胱甘肽(oxidized glutathione,GSSG)、游離脂肪酸(free fatty acid,FFA)、蘇木精-伊紅(hematoxylin and eosin,HE)、油紅O試劑盒 南京建成生物工程研究所;甘油三酯(triglyceride,TG)、總膽固醇(total cholesterol,TC)試劑盒 上海豐匯醫學科技有限公司;TRIzol裂解液 美國Biomiga公司;M-MLV逆轉錄酶、5×逆轉錄緩沖液 美國Promega公司;dNTPs 碧云天生物技術公司;Oligo(dT)、基因引物 上海捷瑞生物工程公司;SYBR Green Master Mix 南京諾唯贊生物科技有限公司;其余試劑均為分析純。
1.2 儀器與設備
5804R臺式高速冷凍離心機 德國Eppendorf公司;Epoch微孔板分光光度計 美國BioTek公司;組織切片機、包埋機、脫水機 德國Leica公司;光學顯微鏡 日本Olympus公司;ETC811 聚合酶鏈式反應(polymerase chain reaction,PCR)擴增儀 東勝興業科學儀器有限公司;900HT Fast實時熒光定量PCR(quantitative real-time PCR,qPCR)儀 美國ABI公司;立式超低溫冷凍冰箱 日本Sanyo公司。
1.3 方法
1.3.1 實驗動物飼喂與分組
飼養條件:SPF級動物房,所有小鼠共處一室,每組6 籠,一籠3 只,自由攝食和飲水,環境溫度(23f2)℃,相對濕度(50f10)%,12 h/12 h晝夜循環光照。
小鼠以正常飼料適應性飼喂一周后,按體質量隨機分為3 組,每組18 只:正常膳食組(CON組,含4%(質量分數,下同)脂肪);高脂膳食組(HF組,含20%脂肪);FCPs干預高脂膳食組(PHF組,含20%脂肪+1%FCPs)。各組飼料參考AIN93純合氨基酸日糧標準和Hasek等[19]的配方進行配制,PHF組的純合氨基酸添加量扣除所加FCPs中各種氨基酸的量。其中添加的脂肪均為豬油,正常飼料的供能總量為16.20 kJ/g,脂肪供能比為11.63%;HF組和PHF組飼料的供能總量為19.55 kJ/g,脂肪供能比為38.54%。
1.3.2 組織樣品的采集
所有小鼠每周稱質量并記錄體質量,每組按體質量隨機分為兩批各9 只,分別于11、22 周后禁食12 h,摘眼球取血于肝素鈉抗凝管中(4 ℃、3 000 r/min離心10 min后取血漿-80 ℃保存備測),斷頸處死。
冰浴條件下迅速摘取肝臟,稱質量記錄后取部分肝臟組織按1∶9(m/V)加入4?℃預冷的生理鹽水制備質量分數10%組織勻漿,4 ℃、3 000 r/min離心10 min取上清液-80 ℃保存備測;取相同部位和大小的兩塊肝臟組織分別放入體積分數4%多聚甲醛固定液和冷凍固定液中備用;再取100 mg左右肝臟組織于TRIzol中剪碎,液氮速凍后-80 ℃保存備用。其余肝臟樣品直接-80 ℃保存。1.3.3 體質量增長率、采食量和表觀脂肪消化率的測定
將小鼠稱質量后放入提前消毒好的代謝籠中適應3 d,開始實驗后,收集每只小鼠72 h的糞便待測。整個實驗流程中所有小鼠均自由采食和飲水。
記錄飼料投入量和投入時間,每24 h稱量并記錄食槽中剩余飼料質量以及漏入代謝籠內的飼料質量。連續監測3 d計算平均數。小鼠體質量增長率及采食量分別按式(1)、(2)計算。

分別將每只小鼠的糞便樣品和飼料磨碎后各取1 g,精確稱質量,分別包入濾紙后在鋁盒中105?℃烘干至恒質量,記錄質量,將濾紙包放入索氏抽提器中進行脂肪抽提4 h以上,至回流液滴在濾紙上揮發后沒有油印為終點。再次將濾紙包烘至恒質量后記錄質量,前后兩次恒質量后的質量差即為糞便和飼料中的粗脂肪質量。表觀脂肪消化率根據式(3)計算。

1.3.4 肝臟脂質水平的測定
按照試劑盒說明書進行TC、TG和FFA水平的測定。
1.3.5 肝臟組織切片觀察
按照試劑盒說明書對固定液中的肝臟組織樣品進行切片和染色,鏡檢,用Image-Pro圖像軟件對切片進行分析,計算肝臟脂肪空泡和脂肪浸潤面積的比例。
1.3.6 肝臟脂代謝相關基因表達水平的測定
采用TRIzol法提取肝臟總RNA,甲醛變性電泳鑒定RNA質量,測定濃度后反轉錄得到cDNA,qPCR測定乙酰輔酶A羧化酶1(acety1 CoA-carboxylase 1,ACC1)、脂肪酸合成酶(fatty acid synthase ,FAS)、固醇調節元件結合蛋白1c(sterol regulatory element-binding protein 1c,SREBP1c)、膽固醇7α-羥化酶1(cholesterol 7α-hydroxylase 1,CYP7A1)、過氧化物激活受體α(peroxisome proliferator activated receptor α,PPARα)和肉毒堿棕櫚酰基轉移酶1(carnitine palmity1 transferase 1,CPT1)的mRNA表達水平,以β-actin為內參,引物序列見表1。最終結果以目的基因相比內參基因的相對表達量來表示。

表1 qPCR引物序列Table1 Sequences of the primers used in qPCR
1.3.7 氧化還原指標的測定
活性氧自由基(reactive oxygen species,ROS)水平的測定采用Lumino電化學發光法[20];T-AOC水平、MDA水平、GSH-Px活力和還原型/氧化型谷胱甘肽(ratio of reduced and oxidized glutathione,GSH/GSSG)水平的測定按照試劑盒說明書操作。
1.4 數據統計分析
數據結果以fs表示,采用IBM SPSS Statistics 22.0軟件對實驗數據進行統計學分析。各組間的比較采用單因素方差分析(One-way ANOVA),對滿足方差齊性的結果用Duncan檢驗進行多重比較,方差不齊時采用Tamhane檢驗。P<0.05時認為差異具有統計學意義。
2 結果與分析
2.1 FCPs對HFD小鼠體質量的影響

圖1 FCPs對HFD小鼠體質量和體質量增長率的影響(n=9)Fig.1 Effect of FCPs on body mass and mass gain rate in mice fed with HFD (n = 9)
在整個飼喂周期中,各組小鼠的體質量從初始時沒有差異(P>0.05),到第11周時為快速增長的階段,隨后進入體質量增長相對平緩的時期(圖1A)。HF組小鼠的體質量增長從高脂飼喂開始后一直高于CON組,第11、22周時均有顯著差異,并且結果有統計學意義(P<0.05);與HF組相比,PHF組的體質量和體質量增長率沒有出現統計學差異(P>0.05)(圖1B)。
2.2 FCPs對HFD小鼠采食量和日均采食總能量的影響
第7周時,HF組的采食量相比CON組有減少趨勢,而PHF組則有所增加,但均未出現統計學差異(P>0.05)。至第14周,與CON相比,HF組的采食量顯著減少(P<0.05);與HF組相比,PHF組的采食量增加,且差異有統計學意義(P<0.05)。到21 周終末,HF組的采食量相比CON組仍保持顯著減少(P<0.05);而PHF的采食量相比HF組明顯增加(P<0.05)(圖2A)。
由于各組飼料配方的不同,同質量的正常飼料與高脂飼料所提供的能量并不相同,因此在采食量的基礎上又統計分析了各組小鼠的日均采食總能量。可見HF組的日均采食總能量與CON組相比在第7、14周均有增加趨勢,但沒有統計學差異(P>0.05);與HF組相比,PHF組的日均采食總能量均有增加趨勢,第7周時無統計學差異(P>0.05),而第14、21周均顯著增加(P<0.05)(圖2B)。
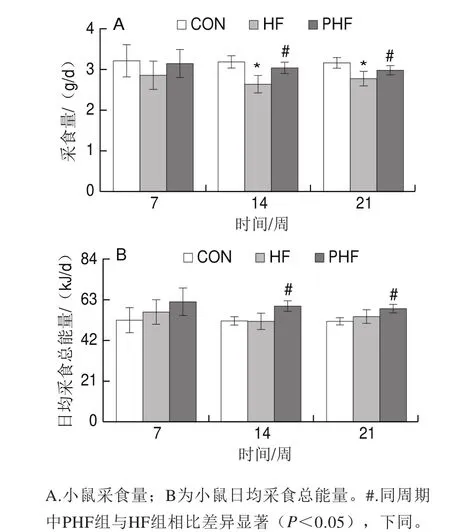
圖2 FCPs對HFD小鼠采食量和日均采食總能量的影響(n=9)Fig.2 Effect of FCPs on daily food intake and total energy intake in mice fed with HFD (n = 9)
2.3 FCPs對HFD小鼠脂肪消化吸收的影響
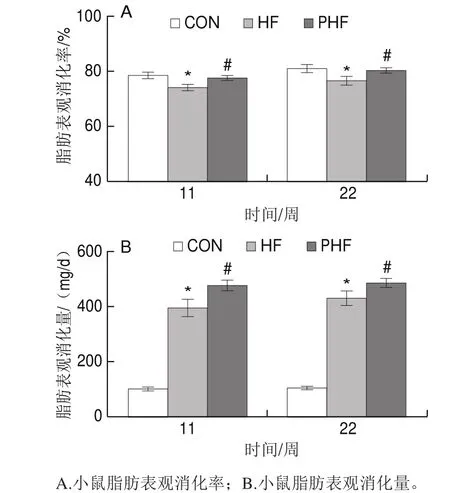
圖3 FCPs對HFD小鼠脂肪消化吸收的影響(n=9)Fig.3 Effects of FCPs on fat digestibility in mice fed with HFD (n = 9)
第11、22周時,與CON組相比,HF組的脂肪表觀消化率顯著降低(P<0.05);相比HF組,PHF組的脂肪表觀消化率顯著升高(P<0.05)(圖3A)。
用脂肪表觀消化率和采食量相乘,計算并統計了各組小鼠的日均脂肪表觀消化量。第11和22周時,相比CON組,HF組的日均脂肪表觀消化量顯著降低(P<0.05);與HF組相比,PHF組的脂肪表觀消化率在第11、22周時均顯著增加(P<0.05)(圖3B)。
2.4 FCPs對HFD小鼠肝臟質量和組織形態的影響
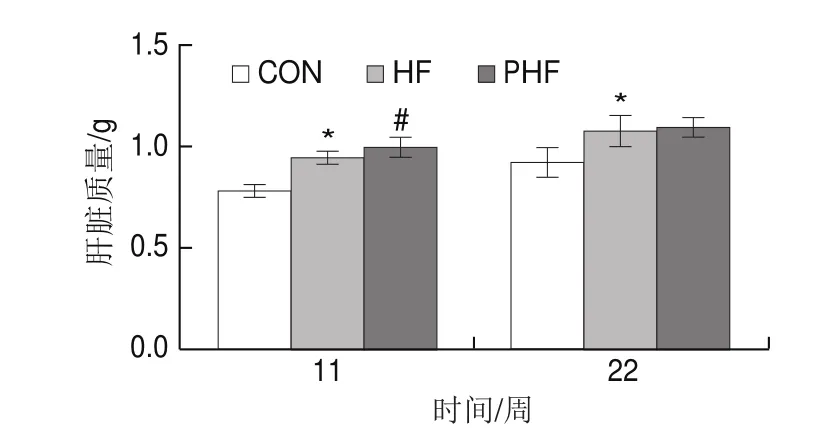
圖4 FCPs對HFD小鼠肝臟質量的影響(n=9)Fig.4 Effect of FCPs on liver mass of mice fed with HFD (n = 9)
由圖4可見,在第11、22周時,相比CON組,HF組的肝臟質量顯著增加(P<0.05);與HF組相比,PHF組的肝臟質量在第11周時顯著增加(P<0.05),至22周時兩者差異沒有統計學意義(P>0.05)。

圖5 FCPs對HFD小鼠肝臟組織形態的影響Fig.5 Effect of FCPs on hepatic morphology in mice fed with HFD
對HE染色的肝臟石蠟切片進行鏡檢,相比CON組,可見第11、22周時HF組小鼠的肝細胞均出現大量脂肪空泡,細胞形狀和排列出現紊亂,空泡面積隨飼喂時間的增長有明顯變大的趨勢;與HF組相比,第11周時,PHF組也出現了較多的脂肪空泡和肝細胞形態紊亂,但空泡較小,第22周時,PHF組的空泡大小和數量都比HF組有明顯改善(圖5A)。
通過油紅O染色小鼠肝臟冰凍切片,其中的脂肪被染為紅色,可以觀察到,相比CON組,HF組小鼠的肝臟脂肪浸潤程度在第11、22周時都明顯增加;與HF組相比,PHF組的肝臟脂肪浸潤程度在第22周時明顯降低(圖5B)。

圖6 FCPs對HFD小鼠肝臟脂肪蓄積的影響(n=4)Fig.6 Effect of FCPs on fat accumulation in liver of mice fed with HFD (n = 4)
由圖6可見,分別統計HE染色切片中的脂肪空泡面積和油紅O切片中的脂肪浸潤面積占視野總面積的比例,來衡量肝臟脂肪蓄積的程度。相比CON組,第11、22周時,HF組小鼠的肝臟脂肪空泡面積比和脂肪浸潤面積比均顯著增加(P<0.05);與HF組相比,PHF組小鼠在第11、22周時的脂肪空泡面積比顯著降低(P<0.05),脂肪浸潤面積比在第11周時沒有顯著差異(P>0.05),第22周時顯著降低(P<0.05)。
2.5 FCPs對HFD小鼠肝臟脂肪含量的影響
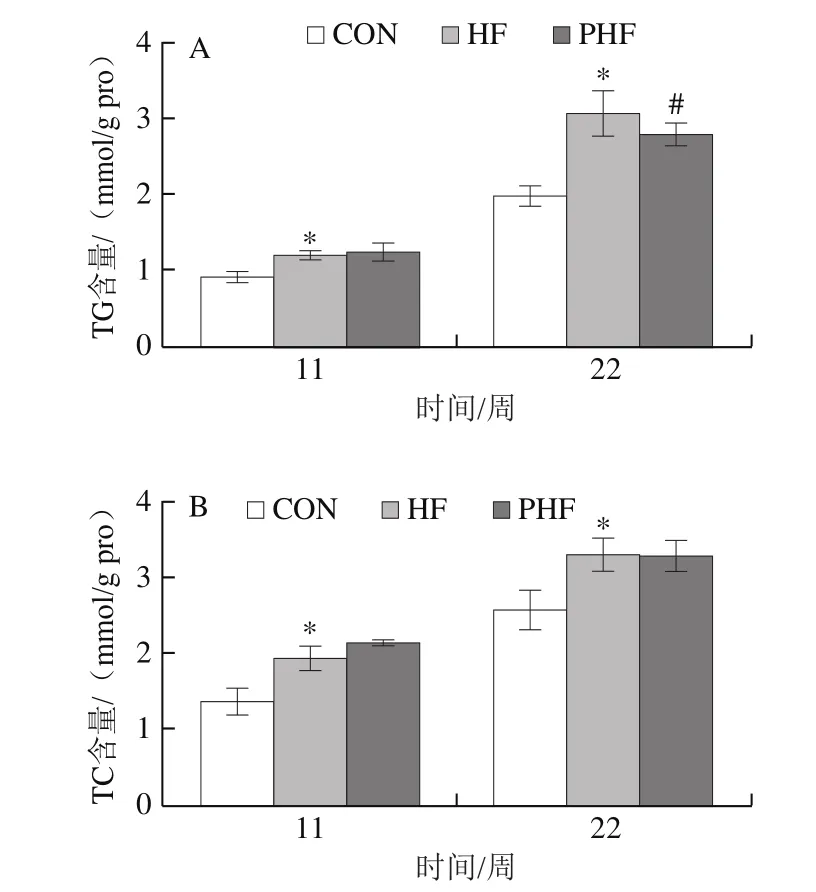

圖7 FCPs對HFD小鼠肝臟脂肪含量的影響(n=9)Fig.7 Effect of FCPs on lipid pro fi les in liver of mice fed with HFD (n = 9)
由圖7可見,與CON組相比,HF組第11、22周時TG、TC和FFA水平均顯著升高(P<0.05);與HF組相比,PHF組在第11周時TG、TC和FFA水平均無統計學差異(P>0.05),至第22周時,PHF組TG和FFA水平顯著下降(P<0.05),TC水平仍無統計學差異(P>0.05)。
2.6 FCPs對HFD小鼠肝臟脂肪合成相關基因表達的影響
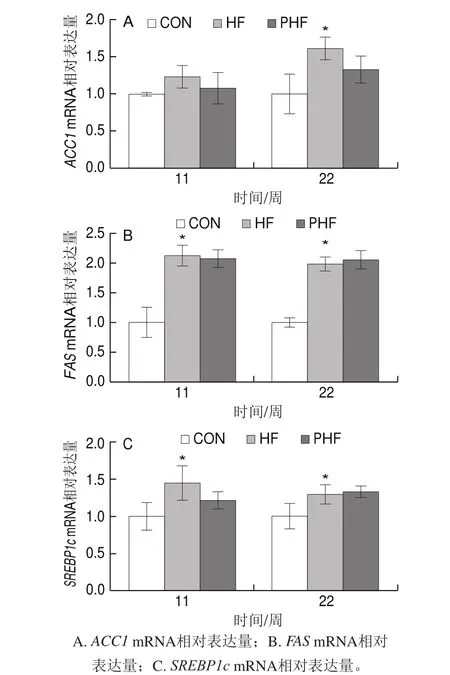
圖8 FCPs對HFD小鼠肝臟脂肪合成相關基因表達的影響(n=9)Fig.8 Effect of FCPs on gene expression related to hepatic lipogenesis in mice fed with HFD (n = 9)
由圖8可見,與CON相比,第11周時HF組的肝臟脂肪合成關鍵基因表達均有上調的趨勢,其中ACC1 mRNA水平沒有統計學差異(P>0.05),FAS和SREBP1c mRNA水平均顯著上調(P<0.05),至第22周時,三者均有顯著上調(P<0.05);相比HF組,PHF組兩個周期中的ACC1 mRNA水平和第11周時的SREBP1c mRNA水平均有下調趨勢,但沒有統計學差異(P>0.05),其他周期中的各脂肪合成關鍵基因表達均沒有明顯變化(P>0.05)。
2.7 FCPs對HFD小鼠肝臟脂肪分解相關基因表達的影響
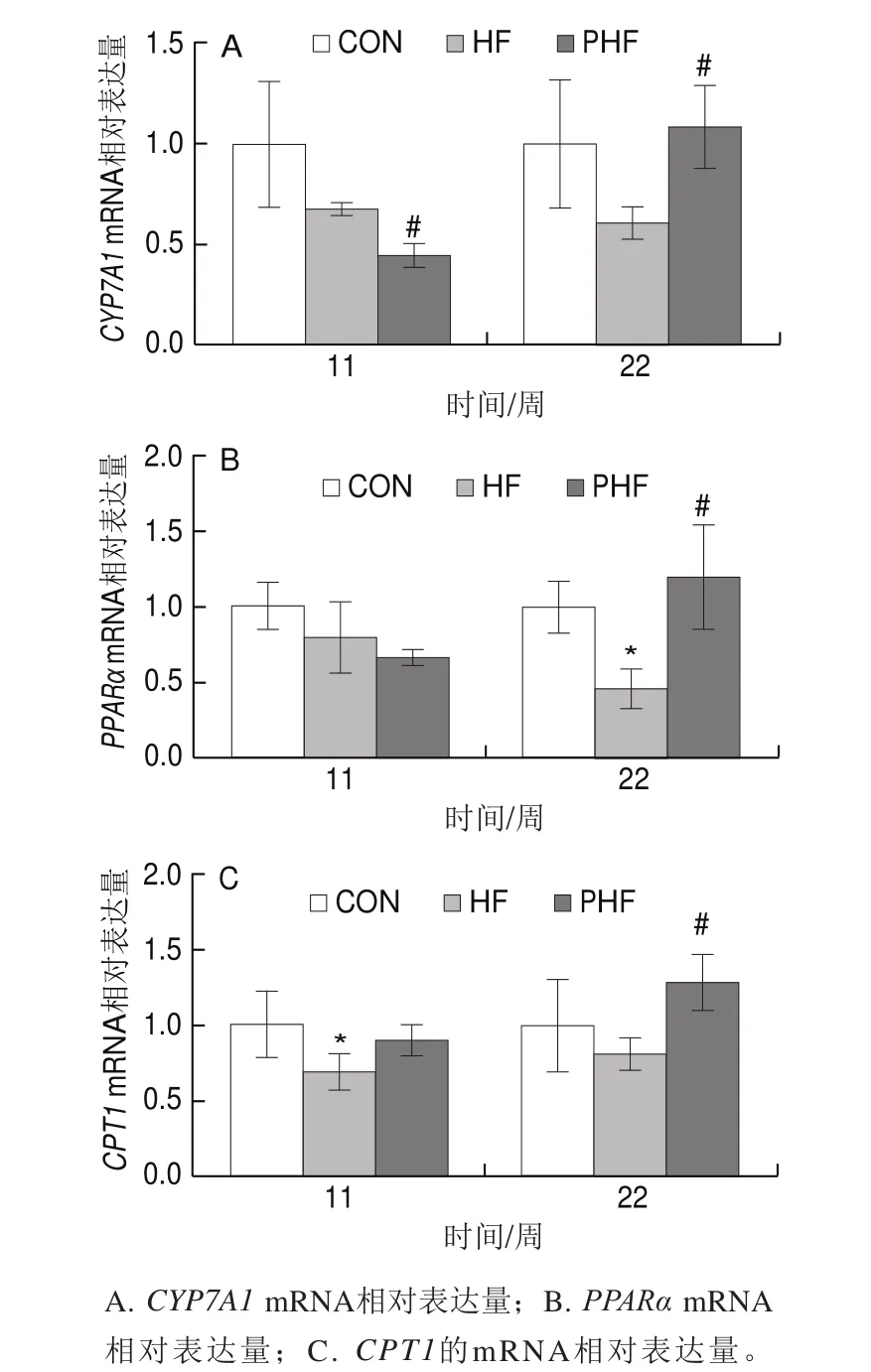
圖9 FCPs對HFD小鼠肝臟脂肪分解相關基因表達的影響(n=9)Fig.9 Effect of FCPs on gene expression related to hepatic lipolysis in mice fed with HFD (n = 9)
由圖9可見,與CON組相比,HF組各周期的脂肪分解關鍵基因表達均有下調的趨勢,其中第11周時的CPT1 mRNA水平和第22周時的PPARα mRNA水平顯著下調(P<0.05),其他周期中的各基因表達沒有統計學差異(P>0.05);相比HF組,第11周時PHF組的CYP7A1 mRNA水平顯著下調(P<0.05),PPARα mRNA水平有下調趨勢但差異沒有統計學意義(P>0.05),CPT1 mRNA水平有上調趨勢但也沒有統計學差異(P>0.05),第22周終末,PHF組的CYP7A1、CPT1和PPARα的mRNA水平均顯著上調(P<0.05)。
2.8 FCPs對HFD小鼠肝臟氧化還原狀態的影響
由圖10可見,與CON組相比,HF組的ROS水平在兩個周期中均顯著增加(P<0.05);相比HF組,PHF組的ROS水平表現出減少趨勢,其中第11周時差異沒有統計學意義(P>0.05),第22周時顯著減少(P<0.05)。

圖10 FCPs對HFD小鼠肝臟ROS水平的影響(n=9)Fig.10 Effect of FCPs on the level of liver ROS in mice fed with HFD (n = 9)

圖11 FCPs對HFD小鼠肝臟氧化還原狀態的影響(n=9)Fig.11 Effect of FCPs on liver redox state in mice fed with HFD (n = 9)
由圖11可見,第11周時,與CON組相比,HF組的肝臟MDA水平顯著增加(P<0.05)、GSH-Px活力和GSH/GSSG比值顯著降低(P<0.05),T-AOC水平沒有統計學差異(P>0.05);相比HF組,PHF組的MDA水平有減少趨勢,但沒有統計學差異(P>0.05),GSH-Px活力和GSH/GSSG比值均顯著升高(P<0.05),T-AOC水平沒有統計學差異(P>0.05)。至第22周終末,與CON組相比,HF組的T-AOC水平、GSH-Px活力和GSH/GSSG比值顯著降低(P<0.05),MDA水平顯著增加(P<0.05);相比HF組,PHF組的T-AOC水平和GSH-Px活力顯著升高(P<0.05),MDA水平和GSH/GSSG比值均有改善,但沒有統計學差異(P>0.05)。
3 討 論
本研究顯示,在HFD中添加質量分數1% FCPs并沒有抑制小鼠體質量的增加,而李亞欣、朱琳玲等采用2%豬骨膠原蛋白肽干預則觀察到了小鼠體質量增長的抑制[14,16],這與膠原蛋白肽的來源、添加量和模型的不同有一定的關系。一般飼料中油脂質量分數超過3%時就會由于膽汁酸分泌量的限制影響禽畜對飼料中脂肪的吸收,而通常HFD飼料中的脂肪含量高達20%,勢必造成其中脂肪不會完全吸收。本研究在FCPs干預后觀察到,在自由攝食、飲水和運動的情況下,PHF組小鼠的采食量、脂肪攝入和能量攝入相比HF組顯著增加,這可能與FCPs作為兩親性物質參與了脂肪的乳化吸收有關。動物攝入脂肪和能量的增加勢必會引起其體質量的增加,然而在本研究中PHF組相比HF組體質量并沒有統計學差異,提示FCPs可能通過增加了機體對脂肪的利用來改善HFD造成體內脂肪積蓄和脂代謝紊亂。
HFD導致的肥胖,造成脂代謝紊亂通常表現為體內脂肪水平的失衡,特別是肝臟的異常增大和脂肪積累,已有研究表明膠原蛋白肽可以顯著改善機體的血脂水平[16-17]。本研究表明,FCPs干預可以顯著改善HFD導致的肝臟脂肪空泡增多、脂肪浸潤面積增大、以及甘油三酯和游離脂肪酸水平的升高,通過減少肝臟的脂肪蓄積對脂代謝紊亂進行調節。
ACC1是催化脂肪酸合成第一步反應的限速酶[21];FAS是脂肪酸合成反應的另一個限速酶,在肝臟組織中的表達較高[22];SREBP1c則是作為轉錄因子選擇性激活脂質合成相關酶的表達,以此來調控肝臟脂質代謝[23]。從實驗結果來看,除FAS mRNA水平外,相比HF組,PHF組FCPs干預對肝臟脂肪合成關鍵調節物ACC1和SREBP1c的mRNA表達均有一定的下調,但是都沒有統計學差異,說明FCPs主要作用途徑可能不是通過抑制脂肪合成實現的。
CYP7A1是膽汁酸合成的限速酶,其上調可以減少腸道對膽汁酸的重吸收并增加機體排出膽固醇[24];PPARα是過氧化物酶體增殖物激活受體的一個亞型,可以改善機體對脂肪酸的轉運和利用[25];CPT1是脂肪酸β氧化的限速酶[26]。本研究表明,與HF組相比,PHF組在FCPs干預11 周時,會顯著下調CYP7A1的mRNA表達,PPARα的mRNA表達水平也有下調趨勢,說明雖然在FCPs的干預下,由于PHF組在中短期內攝入比HF組更多的脂肪和能量,HFD對PHF組小鼠脂肪分解代謝的干擾更為明顯,這與肝臟脂肪水平的結果也是一致的;但到第22周時,FCPs顯著上調了三者的mRNA表達水平。因此,盡管PHF組小鼠的脂肪攝入量和能量攝入量在整個飼喂周期中都顯著高于HF組,并導致中短期飼喂(11 周)時PHF組的CYP7A1和PPARα的基因表達水平下降,但長期飼喂(22 周)時的相關結果說明FCPs對HFD小鼠肝臟脂代謝紊亂的調節,可能與長期飼喂時其促進脂肪分解代謝密切相關。
已有大量研究表明功能因子對HFD導致肥胖的影響與其抗氧化作用密切相關[8-9,17]。長期HFD使機體脂肪攝入處于過量狀態,肝臟作為能量代謝的重要器官,代謝水平持續增高,導致肝細胞需氧量增加,進而誘發更多ROS的生成,引起肝臟中生物大分子的氧化損傷,肝細胞失去完整性影響肝臟功能[27],這是肝臟脂代謝失衡的重要原因。
由于膠原蛋白肽是酶解蛋白得到的混合物,可分離純化得到的具有抗氧化活性的單體肽數量甚多[28-29],所以其抗氧化機制也相對復雜,目前已經報道的有直接淬滅自由基、金屬離子螯合以及提供還原力等多種抗氧化途徑[30]。有研究表明膠原蛋白酶解產物的自由基清除能力與其疏水性氨基酸和亞氨基酸含量有關,而還原力更依賴于羥脯氨酸的含量,也有研究報道羥脯氨酸的游離羥基具有清除超氧陰離子自由基和羥自由基的能力[31-32]。
從本研究的結果來看,FCPs干預HFD小鼠11 周時,相比HF組,PHF組顯著提高了肝臟GSH-Px活力和GSH/GSSG比值,對肝臟MDA水平也有降低的趨勢;第22周后,可以觀察到PHF組肝臟T-AOC水平、MDA水平和GSH-Px活力相比HF組的顯著改善,同時對GSH/GSSG比值也有改善的趨勢,但沒有統計學差異。提示FCPs可能首先通過調節肝臟氧化還原狀態,繼而調節了HFD小鼠肝臟的脂肪分解代謝,但是其中具體的作用機制還需要進一步的研究和驗證。
綜上,質量分數1%的FCPs能夠減少HFD小鼠肝臟中ROS的產生,改善肝臟的抗氧化能力,促進肝臟脂肪分解代謝,減少肝臟中脂肪的蓄積。因此飲食中添加1%FCPs具有改善機體氧化還原狀態和脂代謝紊亂的作用。本研究為FCPs的開發利用提供了更進一步的理論依據,具有一定的研究意義。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
音樂探索(2022年2期)2022-05-30 21:01:37
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
人大建設(2019年12期)2019-05-21 02:55:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
