沉默2b基因獲得抗黃瓜花葉病毒(CMV)的轉基因煙草
2019-03-11 06:44:42芮鵬環宋培培蔣磊江彤
中國煙草學報 2019年1期
關鍵詞:煙草
芮鵬環,宋培培,2,蔣磊,江彤
1 安徽農業大學植物保護學院,合肥 230036;
2 山東省平度市農業局,山東 平度 266700
黃瓜花葉病毒(Cucumber mosaic virus, CMV)是雀麥花葉病毒科(Bromoviridae)黃瓜花葉病毒屬(Cucumovirus)的典型成員[1],其基因組含3個RNA組分,分別為RNA1、RNA2和RNA3, RNA2 3'端ORF編碼2b蛋白,已證明是病毒基因沉默的抑制子[2]。植物通過基因沉默機制抑制病毒在寄主內的積累,抵抗病毒的侵染。為了打破寄主的沉默免疫系統,多數病毒自身可以編碼RNA沉默抑制子,抑制寄主的沉默防衛反應,導致植物發病。基于轉錄后基因沉默(post-transcriptional gene silencing, PTGS)原 理[3-4],沉默病毒的沉默抑制子,有望提高寄主的抗病性。
目前,利用dsRNA技術沉默病毒的沉默抑制子已見報道。將馬鈴薯Y病毒(Potato virus Y, PVY)Hc-Pro基因部分片段順次正反向插入植物表達載體,構建的RNAi載體具有發夾結構,利用農桿菌介導法轉化普通煙草(Nicotiana tabacum)品種云煙85,獲得T1代轉基因高抗PVY的煙草株系[5]。構建蕪菁花葉病毒(Turnip mosaic viruses,TuMV)HC-Pro基因部分片斷反向重復植物表達載體,農桿菌介導法轉化擬南芥,繼代篩選獲得T3代高抗TuMV的轉基因純合擬南芥株系[6]。水稻條紋病毒(Rice stripe virus,RSV)NS2基因和NS3基因編碼的蛋白均具有沉默抑制子功能,將串聯的NS2+NS3分別正反向插入含有1個intron的RNAi載體,轉化水稻獲得31株T0代轉基因水稻植株,接種RSV發現,轉基因水稻發病時間推遲,病毒積累量下降[7]。目前,利用dsRNA技術沉默CMV沉默抑制子2b基因,獲得轉基因抗病毒煙草植株尚未見報道。本研究擬構建CMV2b基因反向重復表達載體,轉化普通煙草品種云煙85,以獲得能夠穩定遺傳的抗CMV轉基因煙草株系,為利用基因工程技術防治CMV提供新的策略。
1 材料與方法
1.1 試驗材料
1.1.1 材料采集安徽省蕪湖縣煙區表現出CMV癥狀的煙草,鑒定后摩擦普通煙草(N.tabacum)作為CMV毒源,烤煙品種云煙85由云南省煙草科學研究所劉勇研究員惠贈,含CMV2b基因的重組載體(pBIN-2b(r, i))由宋培培構建[8],本實驗室保存。
1.1.2 試劑
CMV 抗血清檢測試劑盒購自上海貝優生物技術公司,TaqDNA聚合酶購自上海生工生物工程公司,T4連接酶和克隆載體pMD18-T購自大連寶生物(TaKaRa)公司,DNA消化酶和限制性內切酶購自美國Promega公司,RNA抽提試劑TRIzol購自Invitrogen生物技術有限公司;質粒抽提試劑盒購自天根生化科技(北京)有限公司,反轉錄試劑盒購自北京諾博萊德科技有限公司,抗生素購自美國Sigma公司;其他藥品均購自國藥集團化學試劑公司。
1.2 試驗方法
1.2.1 pBIN-2b(r, i) T0代轉基因煙草的獲得及PCR檢測
反向重復重組載體pBIN-2b(r, i) 凍融法轉化農桿菌GV3101,篩選陽性克隆。將云煙85無菌葉片切成0.8×0.8 cm2大小葉盤,置于MS固體培養基上預培養,農桿菌浸潤侵染新鮮葉盤組織,侵染過的葉盤在分化培養產生愈傷組織,長出抗性芽后轉入生根培養基生長,最后移栽盆缽,獲得轉基因云煙85植株。提取轉基因煙株總RNA,RT-PCR法檢測靶標基因(2b-F: 5'-ATGGAATTGAACGAAGGCGC-3’, 2b-R:5'-T-CAGAACGACCCTTCCG-3’)。
1.2.2 T0代轉基因煙草的抗病性鑒定
稱取CMV感病煙草葉片,按1:10重量體積比加入0.02 mol/L pH 7.2的PBS緩沖液,研磨后作為接種毒源。分別選取5株抗病和感病轉基因煙株,以5株非轉基因煙株作為對照,機械摩擦法接種CMV,15 d后記錄發病癥狀表型,三抗夾心ELISA(Tas-ELISA)測定接種煙株的病毒增殖量[9]。
1.2.3 T1代轉基因煙草的抗病性鑒定
云煙85T0代抗病煙株留種,播種擴繁T1代煙株,CMV摩擦接種7~8葉期T1代煙株,15 d后觀察煙株發病癥狀,記錄抗、感病煙株表型,統計各表型數量。再提取部分抗、感病T1代煙株的總RNA,選擇煙草Actin蛋白作為內參基因(Nt-actin-F: 5’-TG GCTCTTGACTACGAGCAGGAGCTT-3’, Nt-actin-R:5’-ACCACTGAGCAC-AATGTTACCGTAGAGGT-3’),采用Real-time PCR方法,SYBR Green 1染料檢測轉基因云煙85煙株CMV2b基因(2b-RT-F:5’-CGTCGAACTCCAACTGGC-TCGT-3’, 2b-RT-R:5’GAATGGTAGGAAGCGGAACAGCC-3’)的RNA積累水平。
2 結果與分析
2.1 葉盤法轉化云煙85
凍融法將pBIN-2b(r, i)導入農桿菌GV3101,葉盤法轉化云煙85,卡那霉素抗性篩選,共獲得112株T0代再生煙草植株(圖1)。

圖1 葉盤法轉化煙草Fig.1 Tobacco plants transformed via leaf disc
2.2 T0代轉基因煙草陽性植株的PCR檢測
CTAB法提取112株T0代轉基因煙株DNA,PCR擴增發現,從全部T0代轉基因煙株中均能檢測出大小為530 bp的特異性條帶,轉基因煙株陽性率達100%。
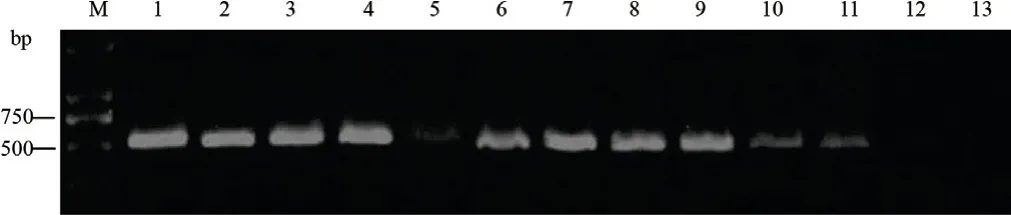
圖2 轉基因煙草植株的PCR檢測Fig.2 PCR detection of the transgenic tobacco plants
2.3 T0代轉基因煙草的抗病性鑒定
將112株T0代轉基因煙草依次編號,分別接種等量CMV,15 d后觀察癥狀。結果發現,46個T0代轉基因煙株不產生任何癥狀,表現為抗病,占41.1%;其他66個轉基因煙株產生不同程度的花葉癥狀,表現為感病,占58.9%;非轉基因煙草接種CMV 15 d后,全部出現嚴重的花葉癥狀(圖3)。

圖3 T0代轉基因煙草植株接種CMV的癥狀表現Fig.3 Symptoms of transgenic T0 transgenic tobacco plants inoculated with CMV
選取T0代轉基因抗病煙株和感病煙株各5株,Tas-ELISA檢測煙株樣本中的病毒含量。結果表明,5個抗病煙株的OD405平均值為0.117,顯著低于5個感病煙株的OD405平均值0.391。轉基因感病煙株的OD405和接種CMV的非轉基因煙株(CK+)OD405相近,轉基因抗病煙株的OD405接近于未接種的非轉基因煙株OD405(表1),且轉基因抗病煙株的OD405平均值顯著低于轉基因感病煙株的OD405。表明轉化反向重復的2b基因確實可以提高轉基因煙株對CMV的抗性。
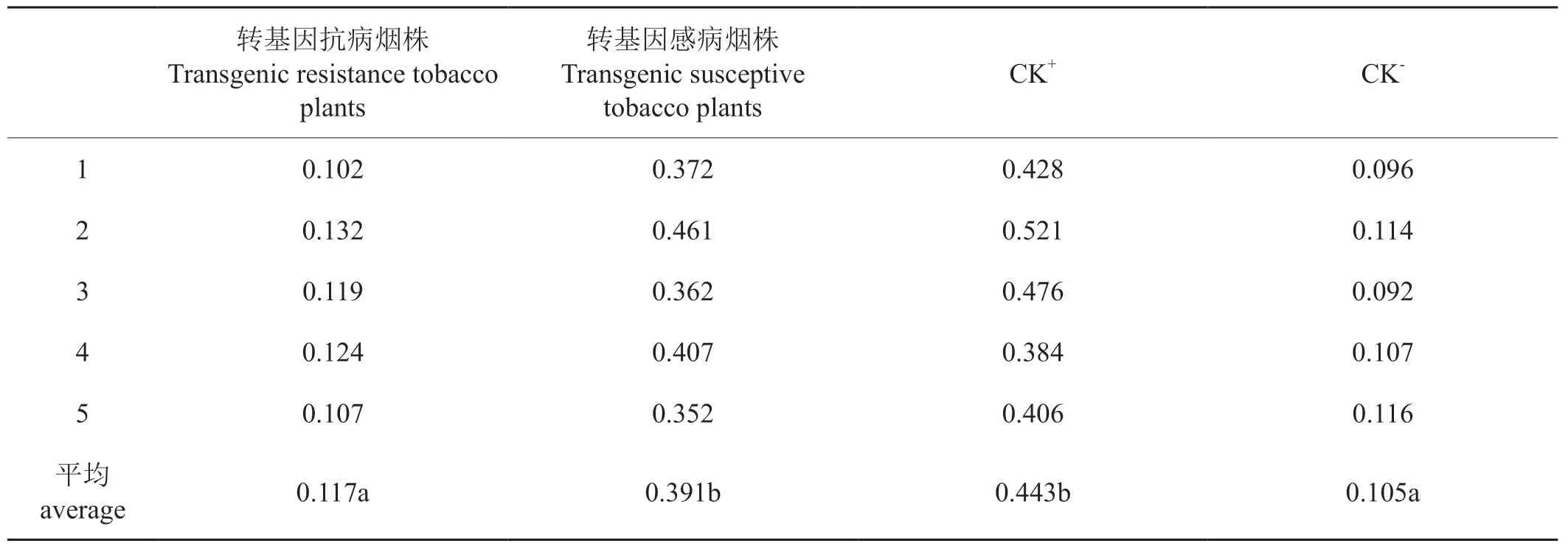
表1 T0代轉基因煙草植株接種CMV后的Tas-ELISA檢測(OD405)Tab.1 Tas-ELISA detection of transgenic T0 tobacco plants inoculated with CMV (OD405)
2.4 T1代轉基因煙草的抗病性鑒定
46株T0代轉基因抗病煙株全部留種,隨機選選擇10個株系播種,每個株系移栽30棵苗,7~8葉期接種CMV,15 d后觀察癥狀。統計結果發現,其中3個T1代煙株抗病株率達100%,7個T1代轉基因煙株發生了不同比例的抗感分離,抗病株率為46.7%~80%。非轉基因煙株接種CMV的發病率達100%(表2)。

表2 T1代轉基因煙草對CMV的抗性鑒定Tab.2 Resistance identification of transgenic T1 tobacco plants to CMV
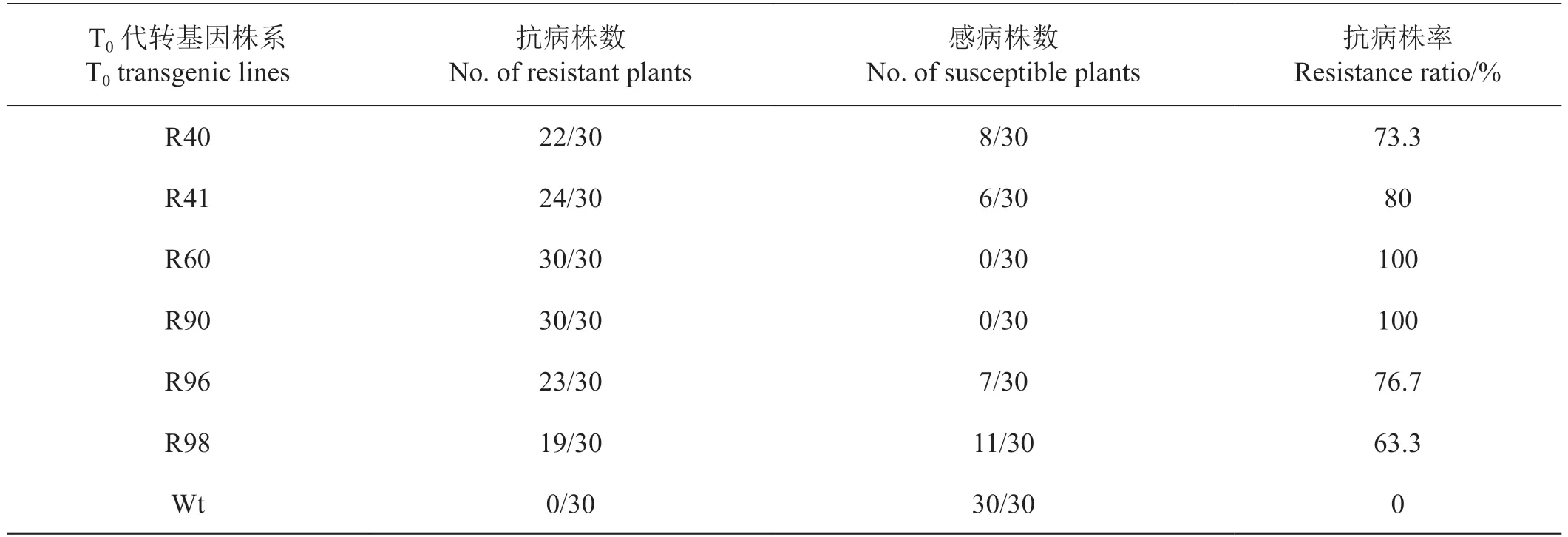
續表2
Real-time PCR檢測CMV2b基因RNA在T1代轉基因煙株的積累水平。結果發現,接種CMV后,表現為抗病的轉基因煙株和表現為感病的轉基因煙株中,CMV2b基因RNA的積累水平差異非常顯著。轉基因抗病煙株中CMV2b基因RNA的積累極低,而轉基因感病煙株中檢測到的CMV2b基因RNA積累水平很高。不接種CMV的轉基因煙株和非轉基因煙株均檢測不到CMV2b基因RNA的積累(圖4)。結果表明,T1代轉基因煙株的抗病性確實是由RNA介導的。
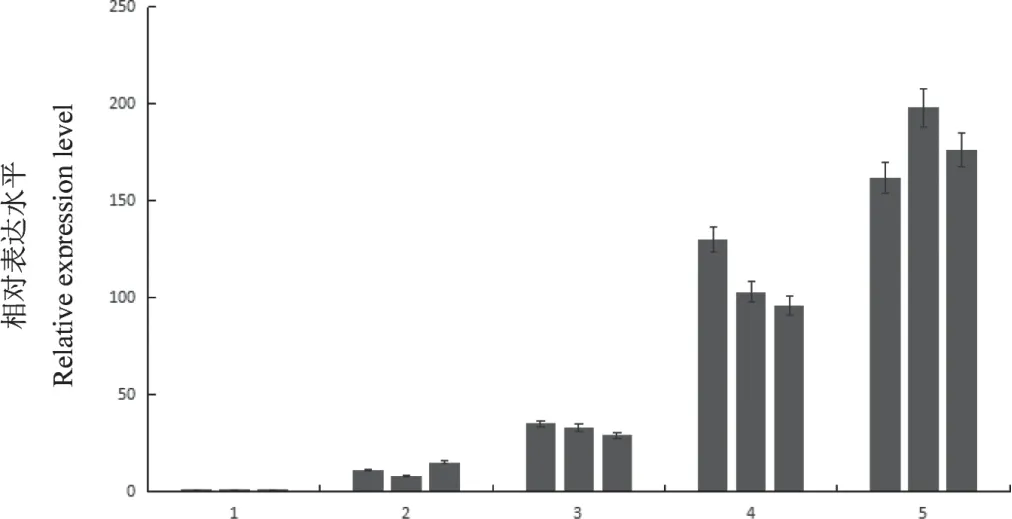
圖4 轉基因煙株中CMV 2b基因RNA積累水平的Real-time PCR檢測Fig.4 Real-time PCR detection of CMV 2b RNA accumulation level of transgenic tobacco plants
3 討論
dsRNA誘導的RNA沉默在抗病毒轉基因工程中廣泛應用,特別是構建含有反向重復序列的載體轉化植物,很容易高效轉錄獲得dsRNA。Smith等[10]在反向重復序列之間插入一段內含子(intron),轉錄后形成dsRNA誘發基因沉默的效率將近100%。朱俊華等[11]發現誘發RNA介導抗性的最短有效基因片段介于202~417 bp之間。因此,本研究也借鑒了這種構建策略,克隆了長度為300~310 bp的2b基因正反向片段,在大豆內含子的兩側分別插入2b基因正反向片段,轉化煙草獲得高抗CMV的轉基因煙株,表明該構建策略具有較好的沉默效果。
Tas-ELISA和Real-time PCR檢測結果表明,CMV侵染轉基因抗病煙株,病毒在煙株體內的積累量及病毒RNA轉錄水平均極低,表明構建的表達載體可以高效沉默CMV2b基因,抵御CMV的侵染。T1代轉基因煙株的抗病性鑒定發現,部分煙草轉化株系發生了不同程度的抗感分離,抗感比率符合3:1,推測可能是單拷貝株系,而抗性未見分離的,推測可能是多拷貝株系。抗感分離原因很復雜,可能與靶基因反向重復片段的長度和位置的選擇有關。另外,外源基因在受體基因組中的插入方式、轉基因的甲基化狀況等也對外源基因的表達效率有影響[14]。
轉基因煙草抗CMV,一般是將CMVcp基因、Rep基因或CMV反義2b基因轉入煙草。本研究是將CMV2b基因反向重復序列插入內含子(intron)兩側,構建表達載體轉化煙草,這種重組載體在轉錄時可以剪切掉intron,構建的2b(i, r)雙臂結構更為緊湊,不易遭受核酶的攻擊,穩定性強;其次,intron剪切的過程不僅有利于互補臂的形成,還有利于轉錄水平的提高[12-14]。另外,這種構建策略還可以避免病毒異源重組帶來的生態安全性問題。本研究已得到了T1代轉基因抗CMV株系,將分析轉基因抗病性遺傳規律,繼代篩選T2代、T3代抗病株系,直到獲得高抗CMV的單拷貝純合株系。
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
