木材細胞壁超微構造的形成、表征及變化規律
2019-03-25 12:37:40孫海燕王玉榮
浙江農林大學學報 2019年2期
孫海燕,王玉榮
(1.中國林業科學研究院 林業新技術研究所,北京 100091;2.中國林業科學研究院 木材工業研究所,北京 100091)
木材是一種多孔性、層次狀、各向異性的非均質天然高分子復合材料,主要由纖維素、半纖維素和木質素3種高分子聚合物組成。木材的實體物質為其細胞壁,細胞壁中的纖維素通過分子鏈聚集成排列有序的微纖絲束,構成了細胞壁的基本骨架[1]。揭示木材細胞壁特別是其骨架的超微構造的形成及變化規律,對木材細胞壁的改性處理以及后續的遺傳改良等具有重要的科學意義。木材細胞壁在超微水平上主要以纖維素微纖絲及結晶區的形式體現,木材科學中常用微纖絲角表征細胞次生壁S2層中微纖絲排列方向與細胞主軸方向的夾角,用結晶度和微晶形態表征結晶區和其基本組成結構的大小[1]。木材細胞壁超微構造即微纖絲和結晶區的研究是木材科學領域的研究熱點之一[2]。近年來關于細胞壁超微構造形成的研究較多,如葡萄糖形成纖維素單鏈,繼而形成微晶、微纖絲和結晶區等過程[3];射線技術、尖端顯微鏡以及光譜類儀器在木材超微結構表征中的應用[4],進一步揭示了微纖絲和結晶區的結構特征。研究發現微纖絲角和結晶度沿軸向和徑向的變化規律不盡相同,對細胞形態的影響也不同[5-6],但對細胞壁微晶形態及變化特點方面的研究還不夠深入;細胞壁超微構造會影響木材密度、干縮性和強度等物理力學性能[7-8],而探究微纖絲角、結晶度和微晶形態的形成及變化規律,是了解木材的基礎性質的重要途徑之一。以往對木材細胞壁超微構造的研究多集中于某一超微構造或某種表征方法方面,對不同種類木材及同種木材不同生長部位細胞壁纖維素微纖絲及結晶區的形成和變化及其表征方法未見系統報導。筆者詳述了木材細胞壁微纖絲和結晶區的形成、表征方法、變化規律及其對細胞形態的影響,以期為今后木材的超微構造的深入研究和為基于細胞壁微纖絲和微晶結構特征來預測木材基礎性質、早期良種選育以及材料的高效利用等方面提供詳細的科學資料。
1 細胞壁超微構造的形成過程
微晶、微纖絲和結晶區均屬于木材細胞壁的超微構造,是纖維素的結構組成部分。纖維素是植物細胞壁的主要組成成分,也是自然界中分布最廣、含量最多的一種多糖,對高等植物細胞壁中天然纖維素結構和形成過程的研究發現,細胞壁超微構造的形成過程并非孤立,而是按照 “微晶—微纖絲—結晶區”的順序形成的。
1.1 細胞壁微晶結構
天然狀態下,纖維素合成酶合成線性葡聚糖鏈的聚合度十分龐大(≥104),即上萬葡萄糖殘基通過β-1,4-糖苷鍵相連形成無分支結構的纖維素單鏈(圖1A)[2-3]。由于含有大量羥基,新合成的相鄰纖維素分子鏈間可產生大量分子間氫鍵,形成有序自組織聚集體;特別是相鄰糖鏈間形成的氫鍵,可使纖維素分子形成穩定的片層結構(圖1B)[9];這些片層結構在范德華力和疏水力等次級鍵作用下自發有序地緊密堆積(圖1C)[10],即為天然結晶纖維素,其中有序結晶的程度可通過X射線粉末衍射法測定的結晶度來表征[11]。由圖1C可知:纖維素基元纖絲中的36根糖鏈聚集形成8個糖鏈片層,片層經氫鍵網絡和范德華力作用堆積為空間結構呈現相對規則的六面體;理論模型橫切面長約5.30 nm,寬3.20 nm,即常說的纖維素微晶[10]。
1.2 細胞壁微纖絲和結晶區
植物細胞壁微纖絲是在纖維素微晶的基礎上形成的。受環境等因素影響,在合成過程中纖維素微晶結構沿著不同晶面聚集生長或沿著某一軸向扭轉,形成大小不同、形狀各異的微纖絲結構(圖1D)[12]。電子顯微鏡觀測可知,微纖絲中晶胞數目不同,晶面聚集方向不一致[13],微纖絲間不能進一步緊密聚集,因而可認為微纖絲是細胞壁中的基本結構單元(圖1D)。定向排列的微纖絲幾何結構發生螺旋狀扭曲,造成微纖絲寬度改變的同時,也形成了纖維素結晶和非晶2種晶態[11],即為纖維素的結晶區和非結晶區。
2 細胞壁超微構造的表征方法
2.1 微纖絲角表征方法
微纖絲角的表征方法隨著儀器制造及分析水平的不斷發展而發展。直接觀測法是最原始的表征方法,適用于細胞壁局部區域微纖絲取向的精細表征,包括直接觀測微纖絲傾角的偏振光顯微鏡法、碘結晶法、激光共聚焦顯微鏡法、紋孔法、原子力顯微鏡法以及電子顯微鏡法等。偏振光顯微鏡法是最早應用的直接觀測法[14],垂直入射的完全偏振光通過試樣后出現消光,此消光角即為木材微纖絲角[15]。原子力顯微鏡法則是通過表征細胞壁中微纖絲聚集體的排列來測定其傾角[16]。
間接法是基于木材光譜特征通過數學計算得到微纖絲角的X射線衍射法、近紅外光譜預測法和拉曼光譜法線法等,適用于大量試樣的平均微纖絲角的研究[4]。目前比較常用的是X射線衍射法[13-14]。X射線衍射法的微纖絲角計算方法中,Turley法是常用的一種方法,它是通過晶面衍射強度曲線最低2點畫切線去除背景的方法計算微纖絲角[17-18]。共聚焦拉曼顯微技術和偏振拉曼顯微技術則是通過分析與纖維素取向密切相關的拉曼光譜峰來獲得細胞壁的纖維素取向[19-20]。
應用直接觀測法還是間接觀測法應當依據測試要求和內容而定。
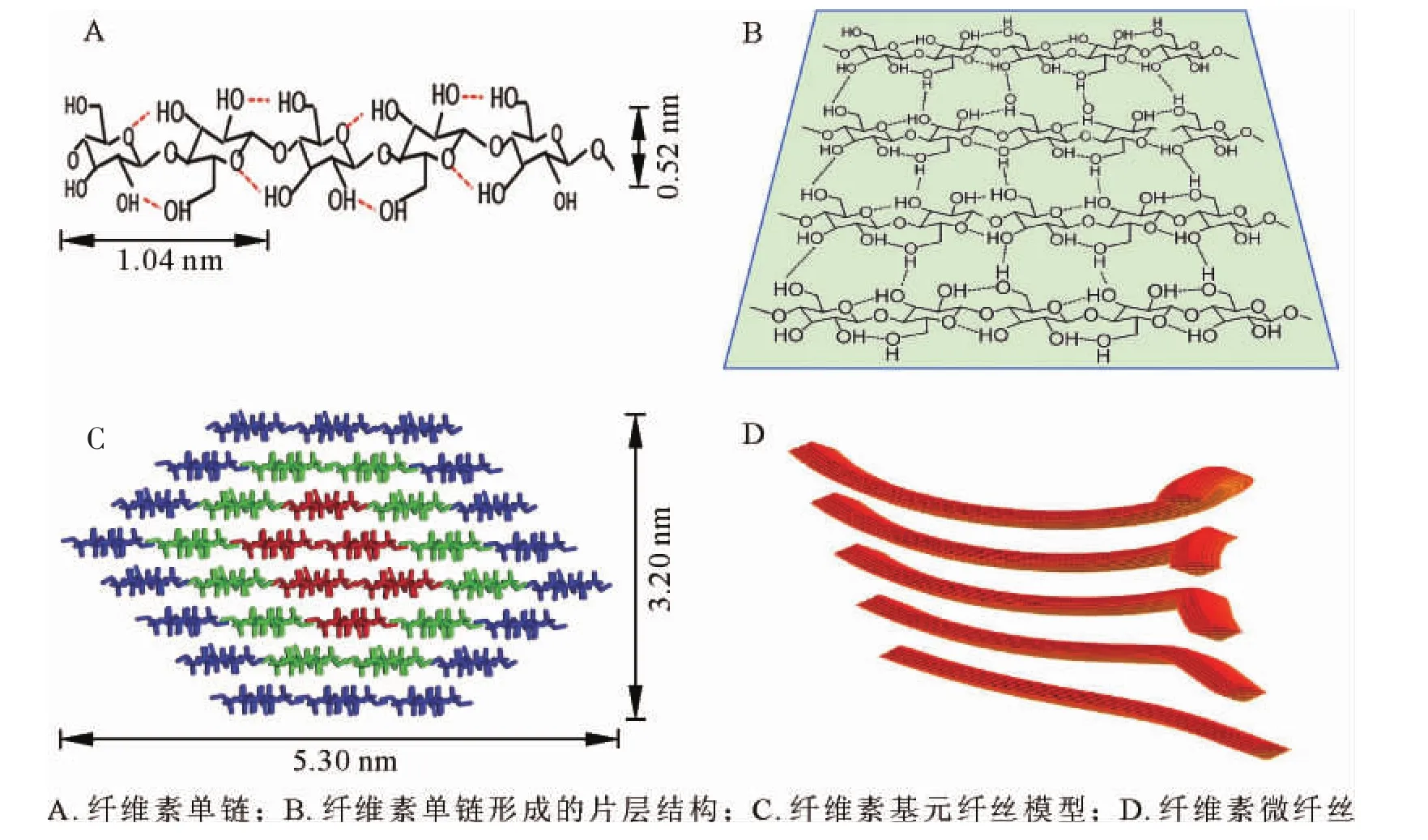
圖1 微晶、微纖絲及結晶區的形成Figure 1 The form diagram of microcrystalline,microfibril and crystalline area
2.2 結晶度和微晶形態表征方法
隨著尖端顯微鏡、射線類以及光譜類儀器不斷地應用于木質纖維材料結構與性能表征中,木質纖維材料細胞壁纖維素結晶度、微晶形態等精細構造特征也不斷地被揭示。檢測方法主要分為原位檢測和非原位檢測兩大類。
原位檢測技術不改變樣品纖維素原本的位置和形態,常用表征方法如原子力顯微鏡技術和X射線法。原子力顯微鏡技術通過監測探針與試樣表面的作用力來表征纖維素結晶區等大分子結構特征[21]。X射線法作原位檢測時通常以1.0~1.5 mm厚的薄木片為樣品,偶有4.0 mm厚的樣品[18,22],作非原位檢測以80~100目的木粉壓制成的薄片為樣品[5,7],根據衍射最強點的強度和位置,測出纖維素纖維晶體分子鏈中的晶區大小和結晶度等,能直接獲得較為準確的結晶度值。其他非原位檢測技術如核磁共振法,以木材硫酸鹽漿為實驗材料,通過區分纖維素無定形區和結晶區的信號得到結晶度值,其值與X衍射方法得到的結晶度值一致[23]。拉曼光譜法通過拉曼特征峰的相對強度來表征結晶度的大小[24],但因目前無法完全去除半纖維素、木質素等對結晶相關特征峰的干擾,該方法還沒有直接應用到木質纖維材料細胞壁微晶形態表征中。
3 木材細胞壁超微構造的變化規律
3.1 微纖絲角的變化規律
樹木木質部細胞次生壁在形成過程中,每一薄層的微纖絲沉積方向和排列密度都在不斷發生變化,因此木材不同位置的微纖絲角不同[25]。微纖絲角決定材料微觀和宏觀的各項性能,直接關系到木材加工利用,被認為是影響木質纖維材料性質的重要指標。關于微纖絲角的株內變異規律目前有較多研究。
3.1.1 徑向變化規律 研究認為,徑向方向上同一年輪中早材的微纖絲角大于晚材;從髓心(幼齡材)到樹皮(成熟材)平均微纖絲角逐漸減小,到一定年齡后趨于穩定。以長白落葉松Larix olgensis為例,從髓心到樹皮微纖絲角在生長的前5 a急劇下降,第5年到第25年呈微小的波動變化,與銀杏Ginkgo biloba,黑楊Populus nigra,垂枝樺Betula pendula等的微纖絲角變化規律一致[8,25]。研究發現:云杉Picea aspoerata,垂枝樺和輻射松Pinus radiata等幼齡材的平均微纖絲角約為30°,幼齡材至成熟材變異幅度一般在10°左右,之后基本穩定[26-28]。目前認為:微纖絲角在徑向產生這種變異的原因有2種。一種認為樹木生長過程中,幼齡期細胞的直徑增長快于長度生長,微纖絲軸向伸長受抑制,微纖絲角較大;進入成熟期后細胞長度生長快于直徑生長,微纖絲在軸向得以延伸,微纖絲角較小[29]。另一種認為原生質流動方向及原生質體分生的纖維素含量越豐富,微纖絲的排列方向越接近細胞軸的方向;隨樹齡的增長,光合產物積累越多,分生細胞細胞壁的纖維素含量增多,微纖絲角越小[6]。
3.1.2 軸向變化規律 木材軸向方向微纖絲角的變化規律表現為基部最大,從基部向上先減小后增加的變化趨勢,但不同材種變化規律不盡相同。如刺楸Kalopanax septemlobus,油松Pinus tabulaeformis,毛白楊Populus tomentosa中最小的微纖絲角分別出現在1.3 m,3.3 m,5.3 m處;輻射松樹高7.0 m以上、毛白楊高9.0 m以上時,微纖絲角趨于穩定,但在梢部的心材中微纖絲角有所增加[6,30-31]。總體來說,微纖絲角軸向變異模式屬于 “大—小—大”的形式。目前關于微纖絲角產生軸向變異的原因尚缺乏明確的解釋。
3.2 結晶度的變化規律
纖維素的結晶區由纖維素大分子鏈有序排列形成,結晶區占纖維素整體的百分數即結晶度,可表征木材纖維素聚集態形成結晶的程度。木材纖維素結晶度在不同樹種及同一樹種不同部位均具有差異性。一般認為:針葉材的纖維素結晶度大于闊葉材。由表1可知:多數針葉材的平均結晶度大于40%,而闊葉材一般為30%~40%[1,7,22,32-40];但也有例外,如楊樹Populus,泡桐Paulownia等低密度闊葉材的纖維素結晶度高于翠柏Calocedrus macrolepis,樟子松Pinus sylvestrisvar.mongolica等針葉材[7,32-34]。結晶度的變化也與不同樹種細胞生長發育階段有關。通常認為隨木質部細胞的不斷發育,纖維素的結晶度會不斷增加,且呈正相關。在徑向方向的結晶度研究表明,隨生長輪齡的增加,結晶度逐漸增大,至成熟后趨于穩定;并且在同一年輪內晚材的結晶度一般比早材的大[5,36,41]。目前,對沿樹軸方向結晶度變化規律的研究不多,表現為自基部向上逐漸增加,到稍部有所減小[36]。

表1 不同樹種木材的結晶度Table 1 Crystallinity of the woods in the different tree species
3.3 微晶形態的變化規律
天然纖維素中微小尺度的晶粒統稱為微晶,常用微晶尺寸表征微晶的形態[42-43]。不同種類木材纖維素微晶的大小和形狀并不均一,一般纖維素微晶寬3.00~5.00 nm,厚2.00~5.00 nm,長十至數百納米,具體形態因樹種而異[42]。對5種針葉材樹種微晶尺寸的研究發現(表2),這些針葉材樹種的微晶寬度接近,為3.00~3.20 nm,但晶體長度則變化較大,為10.00~40.00 nm[43-46];對銀杏幼齡材研究發現,微晶的寬度、長度和樹齡相關性不大[43]。目前,關于木材微晶形態在成熟材和幼齡材中變化規律的研究較少。石江濤等[39]發現白樺Betula platyphlla和水曲柳Fraxinus mandschurica等木材早期組織中纖維素的晶型、晶胞或微晶大小與成熟材不同,但具體差別有待于進一步研究。
4 細胞壁超微構造與細胞形態的相關關系
4.1 微纖絲角與細胞形態的相關關系
微纖絲的排列方向與針葉材管胞的長度和闊葉材纖維的長度相關,微纖絲角是纖維素分子鏈取向的特征指標,與兩者呈不同程度負相關。沿徑向方向,生長的前9 a紅松Pinus koraiensis的晚材管胞長度自髓心向外急劇增加,而微纖絲角逐漸減小,兩者呈顯著負相關(-0.965);此后長度增加減緩,微纖絲角也緩慢減小[47]。同一生長輪內兩者也呈負相關關系,紅松的微纖絲角與管胞長度的相關系數約為-0.70,濕地松Pinus elliottii,油松和翠柏在同一生長輪內管胞長度與微纖絲角的相關系數均為-0.90[34,47],顯示出0.01水平的顯著負相關。由此可見,管胞長度與微纖絲角呈顯著負相關,一定條件下可以通過管胞長度推測纖絲角度。

表2 5種針葉材的微晶尺寸Table 2 Crystal size of the woods in five tree species
闊葉材中微纖絲角與木纖維長度之間也呈負相關,但相關程度要比針葉材低。如尾巨桉Eucalyptus urophylla×E.grandis細胞壁S2層微纖絲角與纖維長度的相關系數為-0.44[48],歐美楊Populus×euramericana中兩者的相關度為-0.39[49]。這可能是因為管胞、纖維長度的變異模式不同;也可能是因為針葉材結構單一,95%以上均是管胞,而闊葉樹材中木纖維只占50%左右,組成比較復雜。
4.2 結晶度與細胞形態的相關關系
纖維素結晶度是衡量木質纖維材料細胞壁結晶程度的一個重要指標,與木質纖維材料的生長特性、組織結構等有密切關系。一般來說,結晶度與管胞、纖維長度呈顯著正相關。研究發現,翠柏的結晶度與早晚材管胞的長度和寬度相關系數在0.90以上[5];浙江桂Cinnamomum chekiange的結晶度與纖維長度和寬度的相關系數在0.95以上[41]。由此認為,利用木材結晶度可以很好地預測木材細胞形態。
總的來說,目前研究多集中在揭示纖維素微晶形態方面,未深入到對其性能影響方面,因此未來需要加強微晶形態對木質纖維材料基礎性能的影響研究。
5 結論與展望
對木材細胞壁微纖絲和結晶區的形成過程、微纖絲角和結晶度表征方法及其變化特點進行綜述發現,葡萄糖殘基最初形成纖維素單鏈,繼而在分子間氫鍵作用下形成穩定的片層結構,然后通過有序堆積方式形成纖維素微晶;微晶在不同晶面聚集成長,形成相互之間不能再緊密聚集的微纖絲結構,并通過微纖絲的扭曲構象形成纖維的結晶態和非結晶態。微纖絲角和結晶度均可以通過尖端顯微鏡、射線類以及光譜類儀器設備表征,常用X射線法,此外也用拉曼光譜法等進行表征。結果發現:木材細胞壁微纖絲角和結晶度變化特點在一定程度上表現出相反的變化規律,即徑向方向從髓心到樹皮微纖絲角逐漸減小,結晶度逐漸增大,最終均趨于穩定;軸向方向從基部向上微纖絲角先減小后增加,結晶度逐漸增加,到梢部有所減小。細胞壁微纖絲的排列和結晶區的大小與其細胞形態相關,微纖絲角越小,管胞和纖維細胞越長,兩者呈負相關關系;結晶度越高,細胞越長,兩者呈正相關關系。
目前,針對細胞壁微纖絲的形成、傾角變化規律和表征方法等已有較為充分的研究,但關于微纖絲角取向形成機制和細胞壁各層厚度分化形成機理還沒有明確的解釋;對纖維素微晶形態的研究已興起,但對從幼齡材到成熟材生長過程中晶型、晶胞及晶體尺寸等微晶形態的具體變化模式還未深入探究。因此,今后工作可以圍繞以下幾點展開:一是從分子層面探究微纖絲取向形成機理;二是加強對木材細胞壁各層厚度累積過程的研究;三是闡明晶型、晶胞及晶體尺寸等微晶形態在木材生長過程中的變化特點。
