HIV-1流行株去糖基化修飾gp41突變體的構建及其表達和純化①
2019-04-01 05:35:34邵繼平
中國免疫學雜志 2019年5期
邵繼平 符 健
(海南醫學院海南省藥物臨床前藥理毒理學研究重點實驗室,海口 571199)
HIV-1病毒(Human immunodeficiency virus type 1,HIV-1)是艾滋病的致病媒介,目前在全球造成3 500萬人感染。HIV-1能攻擊人體免疫系統CD4+T淋巴細胞,使免疫功能喪失,引起各種機會感染,病死率高。HIV-1感染者要經過長達10年或更長的潛伏期后才發展成艾滋病,后期常常發生惡性腫瘤,以至全身衰竭而死。雖然抗病毒藥物可以控制疾病的進程,延長患者的存活時間,但藥物不能清除病毒。而且HIV病毒高度變異,患者要不斷更換藥物,這也增加了艾滋病的治療難度。因此至今尚無根治艾滋病的特效藥物,也無有效的預防疫苗[1-2]。
糖基化作為一種主要的翻譯后修飾在病毒顆粒的包裝、成熟、感染力和免疫逃避中起著重要作用。Doores研究了不同細胞來源的病毒粒子糖基化特點,他們的結果顯示Env糖基化相當簡單,幾乎完全由高甘露糖型聚糖組成,與產生病毒的細胞來源無關[3,4]。以往的研究表明一半以上的HIV-1包膜糖蛋白(Envelope glycoprotein,Env)由聚糖組成,聚糖參與了Env的正確折疊,有助于糖蛋白功能的發揮,并通過多種機制維持病毒的生物學特性[5-6]。比如通過產生糖盾(Glycan shield)來逃避免疫系統的攻擊[7,8];或掩蓋潛在的蛋白抗原表位,使其免受機體免疫系統監視;或以無定形的塊狀物(Amorphous masses)來改變蛋白結構,使病毒蛋白表面免受抗體攻擊;而且絕大部分Env是免疫沉默蛋白(Immunologically silent),為免疫逃避創造了條件[9]。糖基化有助于HIV病毒的感染,但Env 僅糖基化不足以形成天然的病毒粒子,其前提是聚糖要對號入座,不能錯位。
病毒刺突(Spike)是中和抗體的唯一靶點,因此作為疫苗免疫原,Env是該領域的研究熱點[10,11]。HIV-1 Env基因全長約2.6 kb,是變異性最強的基因。不同的HIV-1 Env含有20~30個不等的N糖基化位點。糖基化位點的數量與分布會影響HIV-1 Env的結構和感染力。N糖基化位點少的病毒感染力強,研究顯示包膜蛋白V2區或V4區低糖基化與HIV-1的傳播能力相關[12-15]。為了進一步證實糖基化位點的數量和分布與病毒感染及致病性的關系,本文利用基因定點突變的方法,將HIV-1 ⅢB、CRF-BC 和CRF-AE 流行株野生型gp41基因中特定的N611Q糖基化位點天冬酰胺Asn突變為谷氨酰胺Gln,使該位點去糖基化,構建了HIV-1 ⅢB、CRF-BC和CRF-AE 流行株野生型和突變型pcDNA3.1-gp41的表達載體。挑選測序正確的陽性克隆轉染293 T 細胞,重組蛋白經Ni-NTA柱親和層析純化后采用SDS-PAGE電泳分析。本文為研究去糖基化修飾gp41蛋白與疾病發生發展的分子機制奠定了基礎。
1 材料與方法
1.1材料
1.1.1實驗材料 大腸桿菌DH5a感受態細胞、哺乳動物細胞表達載體pcDNA3.1和HEK 293T細胞由本實驗室保存。限制性內切酶XhoⅠ和NheⅠ購自NEB有限公司;Ex Taq試劑盒、T4DNA連接酶等工具酶以及點突變試劑盒均購自TaKaRa公司;質粒小提試劑盒、DNA膠回收試劑盒及PCR產物純化試劑盒均購自Promega公司;蛋白相對分子質量標準購自Ferments公司;RPMI1640、DMEM 培養基購自Gibco公司;胰酶、脂質體轉染試劑盒Lipofecta-mine 2000購自Invitrogen公司;Ni-NTA Resins購自Novagen公司;辣根過氧化物酶標記羊抗鼠IgG(HRP-IgG)和辣根過氧化物酶標記羊抗兔IgG(HRP-IgG)二抗均購自美國Sigma-Aldrich公司。
1.1.2主要儀器和設備 PCT-2000型PCR基因擴增儀購自美國Bio-Rad公司;GeneGenius 全自動凝膠成像儀購自美國Syngene公司;NANO核酸定量儀購自美國Thermo公司;5430R型冷凍離心機購自德國Eppendorf公司;電子天平購自梅特勒-托利多儀器(上海)有限公司;搖床購自哈爾濱市東明醫療儀器廠;電熱恒溫水槽購自上海一恒科技有限公司。
1.2方法
1.2.1野生型gp41引物設計 根據GenBank數據庫中已發表的HIV-1 gp41基因序列,利用Primer5.0 軟件,設計并合成了PCR引物(見表1)。同時根據表達載體pcDNA3.1的序列,在gp41基因的N-端引入CD14信號肽,使外源基因以分泌形式表達于胞外,在C-端添加His標簽序列、酶切位點和終止密碼子。在上游引物5′端加上NheⅠ酶切位點,下游引物5′端加上XhoⅠ酶切位點。經引物軟件分析,引物自身不互補或形成發夾結構。設計好的gp41基因長度約為700 bp。
1.2.2去糖基化gp41引物設計 運用定點突變的方法去除gp41 N-糖基化位點,將天冬酰胺突變為谷氨酰胺。破壞這些位點的糖基化修飾能力。設計并合成了一系列相應的DNA引物(表1),利用CO-PCR的方法進行目的基因的擴增。

表1 PCR 引物Tab.1 PCR primers
1.2.3野生型和去糖基化gp41表達載體的構建及鑒定 分別以表1所列的引物對作為上、下游引物,以env質粒為模板,擴增野生型和去糖基化gp41基因片段,再進行目的基因和載體雙酶切,12 h后瓊脂糖凝膠電泳鑒定、切膠回收酶切產物。PCR產物克隆到pcDNA3.1 的NheⅠ與XhoⅠ位點之間,連接產物轉化大腸桿菌DH5α的感受態細胞,在氨芐青霉素選擇壓力下挑取單菌落至LB培養液中,37℃振蕩培養過夜,用堿裂解法小量提取質粒后進行PCR和雙酶切鑒定,挑選正確的陽性克隆送上海英駿公司測序,采用酶切和Sanger 測序的方法鑒定插入片段。并進行目的基因的序列比對。測序正確的重組質粒分別命名為pcDNA3.1/gp41-ⅢB、pcDNA-3.1/gp41-BC、pcDNA3.1/gp41-AE。
運用定點突變的方法去除野生型gp41 N-糖基化位點,破壞這些位點的糖基化修飾能力。應用PCR方法,以鑒定好的pcDNA3.1/gp41質粒為模板,以表1所列的引物對作為上、下游引物,按照TaKaRa 點突變試劑盒方法對N 611的糖基化位點進行突變,經測序驗證突變正確后,將此突變型重組質粒命名為pcDNA3.1/Mgp41-ⅢB、pcDNA3.1/Mgp41-BC、pcDNA3.1/Mgp41-AE。
1.2.4序列比對分析 進入網頁:http://www.ncbi.nlm.nih.gov/,打開Blast,點擊“Nucleotide-nucleotide BLAST(blastn)”,將測序序列直接拷貝到方框里,點擊“BLAST”,新頁面完全出來以后,點擊“Format”。將目的基因序列和測序序列進行比對。
1.2.5野生型和去糖基化gp41重組蛋白的表達 選擇HIV-1 ⅢB 標準株、中國大陸地區主要流行株CRF-BC 和海南省流行株CRF-AE gp41序列構建了重組質粒。293T細胞用含10%FBS、100 U/ml青霉素、100 U/ml鏈霉素的DMEM培養基在37℃、5%CO2條件下常規培養。采用Lipofectamine 2000進行轉染,具體操作按試劑盒說明書進行。轉染前24 h 以2×105ml-1的密度將細胞接種于60 mm培養皿中,當貼壁細胞密度達到90%融合度時即可進行轉染。將1 mg質粒與100 ml PBS混合,再將1 ml脂質體與10 ml PBS混合,將兩者輕輕混勻后,室溫靜置15 min,滴加入培養皿中。以pcDNA3.1/gp41和pcDNA3.1/Mgp41重組質粒轉染293T細胞,進行重組蛋白的真核表達。
1.2.6野生型和去糖基化gp41重組蛋白的純化 轉染36~48 h后收集培養上清液,4 000 r/min低速離心5 min,去除細胞碎片。取2 ml Ni-NTA樹脂裝柱,用pH7.0磷酸鹽緩沖液洗柱,直至流出液pH為7.0。將收集的細胞上清液過Ni-NTA柱,進行gp41重組蛋白的純化。將得到的重組蛋白洗脫液進行離心超濾,將蛋白滯留在超濾管上,以達到濃縮的效果。純化的gp41融合蛋白能穩定存在于溶液中。
1.2.7免疫雜交(Westem blot)分析 取25 μg純化后的蛋白,加適量2×SDS-PAGE 上樣緩沖液混勻,100℃煮5 min,離心1 min。利用12%SDS-PACE膠電泳分離蛋白,再電轉于硝酸纖維素膜上,80 V,進行2 h免疫雜交。5%的脫脂奶粉封閉2 h,加兔抗HIV-1 gp41多肽抗體為一抗(1∶1 000)進行孵育,洗膜后再加辣根過氧化物酶標記的羊抗兔IgG(1∶5 000)為二抗孵育,進行免疫雜交分析。最后加顯色液,顯影,定影。
2 結果
2.1野生型和突變型 HIV-1 gp41的結構 gp41跨膜糖蛋白相對分子質量為41 000,由345個氨基酸組成,在HIV-1病毒進入靶細胞中起重要作用。gp41主要由三部分構成:膜外區(aa512-683)、跨膜區(aa684-705)和胞質區(aa705-856)。膜外區又分為5個功能區:融合多肽(FP,aa512-527);polar區(PR,aa525-543);N端重復序列(NHR,aa546-581)和C端重復序列(CHR,aa628-661);抗原決定環(ID環,aa598-604)。NHR和CHR可形成六聚體(6-HB),是膜外區的核心結構。gp41結構如圖1所示。
2.2野生型和突變型 HIV-1 gp41基因的擴增 運用PCR方法擴增野生型和突變型 HIV-1 gp41基因,DNA電泳結果出現了700 bp左右的目的條帶,與預期的基因大小一致,如圖2所示。
2.3野生型和突變型重組質粒的雙酶切鑒定 根據pcDNA3.1載體的圖譜特征,選用NheⅠ與XhoⅠ

圖1 HIV-1 gp41 結構功能域Fig.1 Schematic diagram of HIV-1 gp41
雙酶切重組質粒來鑒定其插入方向。圖3結果顯示雙酶切后,電泳圖上出現了大小約700 bp的基因片段和約4 500 bp的載體片段。我們初步證明所挑選的陽性克隆為正確的重組轉化子。
2.4野生型和突變型DNA序列的同源比對 用測序引物對重組質粒進行測序,進一步確定其插入方向及閱讀框的正確性。Chromas結果顯示四種堿基的信號強,峰值高,表明測序的特異性好。所測得的DNA序列經Genetool軟件比對發現插入片段寡核苷酸序列與設計的靶點序列完全吻合,gp41基因被正向插入于NheⅠ和XhoⅠ雙酶切位點之間。閱讀框分析結果證實了該重組質粒的正確性,且無任何突變,與預期一致(圖4)。由鑒定結果可以看出:重組質粒構建成功。
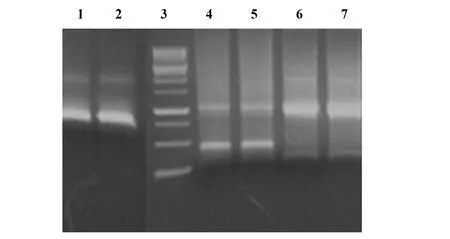
圖2 Gp41基因的PCR擴增Fig.2 PCR amplification for gp41 gene

圖3 雙酶切鑒定gp41 基因Fig.3 NheⅠand XhoⅠdouble digest for gp41 gene
2.5野生型和突變型重組蛋白的免疫印跡分析 取25 μg蛋白加入適量的2×SDS上樣緩沖液,迅速混勻后100℃加熱3~5 min按常規方法進行SDS-PAGE電泳分離總蛋白,并電轉至硝酸纖維素膜上(BIO-RAD半干轉膜,80 V,2 h)。以抗gp41抗體為一抗(1∶1 000),HRP標記的羊抗兔IgG為二抗(1∶5 000),進行Western blot分析。圖5結果表明gp41融合蛋白在293T細胞得到大量表達,表達的融合蛋白可被抗gp41的特異性抗體所識別,在26 000 處出現了特異性反應條帶,條帶的大小與預測的目的蛋白大小一致,本研究采用真核表達系統成功表達了gp41融合蛋白。從圖5中Lane 1到Lane 6融合蛋白的免疫雜交結果可見在26 kD處出現陽性條帶。
2.6野生型和突變型重組蛋白的濃度檢測 用Nano紫外分光光度計測定純化的野生型和突變型重組蛋白,濃度和含量如表2所示。可見野生型和突變型gp41蛋白能在293T細胞中大量表達。

圖4 gp41 基因的測序圖譜Fig.4 Sequencing map for gp41 gene

圖5 重組蛋白的免疫印跡分析Fig.5 Western blot for gp41 recombinant proteins

表2 gp41重組蛋白濃度Tab.2 Concentration of gp41 recombinant protein
3 討論
艾滋病已成為威脅人類健康的重大傳染病之一。中國是艾滋病感染人數眾多的國家。自1995年后我國HIV的流行進入快速增長期,截至去年9月底,全國存活的艾滋病感染者為74.7萬。目前艾滋病整體疫情處于低流行狀態,但又呈增長趨勢。
蛋白質糖基化的分子機制與疾病的發生發展密切相關,已成為生物學研究領域中的熱點之一。HIV-1 Env是一種高度糖基化的糖蛋白,其糖基化程度對蛋白穩定性和功能方面具有重要的影響,與其生物學功能的發揮密切相關。病毒包膜蛋白在加工過程中需要進行N-糖基化,即N-X-S/T(X為除P之外的任意氨基酸)修飾。N-聯糖基化是一種蛋白翻譯后修飾,合成糖基側鏈的過程,在很多方面影響蛋白的功能,比如蛋白正確折疊,維持蛋白構型、改變蛋白空間結構,增加蛋白的穩定性、調節蛋白的抗原性,影響病毒包膜蛋白與受體、輔助受體的結合、介導膜融合和病毒進入細胞,從而影響HIV病毒的傳播等。包膜蛋白過度糖基化可能會掩蓋其抗原表位,導致病毒逃逸免疫應答。
我國主要的HIV-1流行株是B′,CRF07-BC以及CRF01-AE亞型。因此,艾滋病研究要從我國的實際情況出發。本文利用基因定點突變的方法,將HIV-1 ⅢB、CRF-BC 和CRF-AE 流行株野生型gp41基因中特定的N611Q糖基化位點天冬酰胺Asn突變為谷氨酰胺Gln,使該位點去糖基化,成功構建了HIV-1 ⅢB、CRF-BC 和CRF-AE 流行株野生型和突變型pcDNA3.1-gp41的表達載體,分別命名為pcDNA3.1/gp41-ⅢB、pcDNA3.1/gp41-BC、pcDNA3.1/gp41-AE和pcDNA3.1/Mgp41-ⅢB、pcDNA3.1/Mgp41-BC、pcDNA3.1/Mgp41-AE。挑選測序正確的陽性重組質粒轉染293 T細胞,收獲的重組蛋白經Ni-NTA柱親和層析純化后采用SDS-PAGE電泳分析,蛋白條帶單一,幾乎無雜帶,說明Ni-NTA柱純化出來的蛋白純度高,經Western blot驗證其能特異性結合抗gp41抗體,而且gp41目的蛋白在真核體系的表達量分別高達2.786、1.78、3.495、2.06、2.9和1.9 mg。本文為研究去糖基化修飾gp41對病毒致病力的影響與疾病發生發展的分子機制奠定了基礎。
