蜂房哈夫尼亞菌對(duì)粘質(zhì)沙雷氏菌群體感應(yīng)信號(hào)分子產(chǎn)生與生物被膜形成的調(diào)控
2019-04-12 05:34:24李婷婷高娜娜于海鳳國(guó)競(jìng)文王當(dāng)豐勵(lì)建榮黃建聯(lián)
食品科學(xué) 2019年6期
李婷婷,高娜娜,于海鳳,國(guó)競(jìng)文,王當(dāng)豐,勵(lì)建榮,*,黃建聯(lián)
(1.大連民族大學(xué)生命科學(xué)學(xué)院,生物技術(shù)與資源利用教育部重點(diǎn)實(shí)驗(yàn)室,遼寧 大連 116600;2.渤海大學(xué)食品科學(xué)與工程學(xué)院,遼寧省食品安全重點(diǎn)實(shí)驗(yàn)室,生鮮農(nóng)產(chǎn)品貯藏加工及安全控制技術(shù)國(guó)家地方聯(lián)合工程研究中心,遼寧 錦州 121013;3.福建安井食品股份有限公司,福建 廈門 361022)
群體感應(yīng)是一種微生物細(xì)胞與細(xì)胞間的通訊方式,細(xì)菌通過自身產(chǎn)生的一些信號(hào)分子感知環(huán)境中與其共存細(xì)菌產(chǎn)生的信號(hào)分子,當(dāng)信號(hào)分子質(zhì)量達(dá)到一定濃度時(shí)啟動(dòng)細(xì)菌中相關(guān)靶標(biāo)基因的表達(dá),調(diào)控微生物的特定功能,如運(yùn)動(dòng)性介導(dǎo)的銅綠假單胞菌與根癌土壤桿菌在生物膜共培養(yǎng)中的相互作用、熒光假單胞菌胞外酶的分泌及霍亂弧菌毒力因子的表達(dá)等[1-3]。革蘭氏陰性菌一般由LuxR/LuxI型系統(tǒng)調(diào)控,以N-酰基高絲氨酸內(nèi)酯(N-acyl homoscrinc lactones,AHLs)為信號(hào)分子;革蘭氏陽(yáng)性菌的信號(hào)分子一般為寡肽類化合物(autoinducing peptides,AIPs);AI-2是細(xì)菌種間交流的信號(hào)分子,其結(jié)構(gòu)是一種呋喃硼酸二酯類物質(zhì)[4-6]。此外,細(xì)菌還存在其他群體感應(yīng)系統(tǒng)[7]。
蜂房哈夫尼亞菌(Hafnia alvei)命名為H. alvei-14,屬于腸桿菌科哈夫尼亞屬,革蘭氏陰性菌,主要存在于污水、乳制品、土壤、人和動(dòng)物糞便中,是一種細(xì)菌性污染菌[8]和條件致病菌[9]。粘質(zhì)沙雷氏菌(Serratia marcescens)命名為S. marcescens-01,屬于沙雷氏菌屬,革蘭氏陰性菌,是腸桿菌科的一種條件致病菌,存在于水、土壤、動(dòng)植物及人類的腸道和呼吸道中,能夠產(chǎn)生核酸酶、蛋白酶、脂肪酶等次級(jí)代謝產(chǎn)物,對(duì)人類存在潛在傷害[10]。
導(dǎo)致水產(chǎn)品腐敗的實(shí)際環(huán)境為多菌種混雜共存并相互影響,單菌落存在的可能性非常小[11-12]。但目前多集中在單菌種的研究上,對(duì)基于群體感應(yīng)的2 株菌共培養(yǎng)體系的研究缺乏有效的方法和理論指導(dǎo),對(duì)菌與菌之間生態(tài)關(guān)系及群體感應(yīng)方面的共生機(jī)理仍不清楚。而微生物生長(zhǎng)特性的表達(dá)受生長(zhǎng)階段、營(yíng)養(yǎng)狀態(tài)等環(huán)境因素的影響,環(huán)境條件的改變會(huì)調(diào)控細(xì)菌群體感應(yīng)活性,進(jìn)而影響AHLs的分泌和生物被膜的形成。本實(shí)驗(yàn)擬從腐敗鱸魚中分離到的2 株具有群體感應(yīng)現(xiàn)象的H. alvei-14和S. marcescens-01為研究對(duì)象,探討在不同NaCl濃度、碳源、氮源、起始pH值條件下,與蜂房哈夫尼亞菌共培養(yǎng)的粘質(zhì)沙雷氏菌AHLs產(chǎn)生和生物被膜形成能力,旨在研究菌株在共培養(yǎng)條件下AHLs的分泌規(guī)律和生物被膜的形成能力,為水產(chǎn)品的保鮮和腐敗菌的防治提供理論依據(jù)。
1 材料與方法
1.1 材料與試劑
1.1.1 原料與菌株
鮮活海鱸魚購(gòu)于錦州市林西水產(chǎn)市場(chǎng),平均質(zhì)量(1.5±0.5)kg。
供試菌株:從腐敗鱸魚中分離所得,經(jīng)16S rRNA序列比對(duì)鑒定為H. alvei-14和S. marcescens-01;報(bào)告菌株:紫色桿菌CV026(Chromobacterium violaceum CV026)、根癌農(nóng)桿菌A136(Agrobacterium tumefaciens A136),均由本實(shí)驗(yàn)室保藏。菌株均在LB液體培養(yǎng)基中,28 ℃、160 r/min搖床振蕩培養(yǎng),CV026活化時(shí)需添加20 μg/mL卡那霉素,A136需涂布20 mg/mL 5-溴-4-氯-3-吲哚-β-D-半乳糖苷(5-bromo-4-chloro-3-indolyl β-D-galactopyranoside,X-gal)。
1.1.2 試劑
信號(hào)分子標(biāo)準(zhǔn)品(C4-HSL、C6-HSL、C8-HSL、C10-HSL、C12-HSL、C14-HSL) 美國(guó)Sigma公司;卡那霉素國(guó)藥集團(tuán)化學(xué)試劑有限公司;細(xì)菌基因組DNA提取試劑盒、聚合酶鏈?zhǔn)椒磻?yīng)(polymerase chain reaction,PCR)引物 生工生物工程(上海)股份有限公司;其他試劑均為分析純。
1.2 儀器與設(shè)備
PCR儀 德國(guó)Eppendorf公司;Quantity One凝膠成像系統(tǒng)、imark酶標(biāo)儀 美國(guó)Bio-Rad公司;HZQ-X300C型恒溫振蕩器、BPS-100CA恒溫恒濕培養(yǎng)箱 上海一恒科技有限公司;MS105UD電子分析天平 瑞士梅特勒-托利多有限公司;Purifier Logic生物安全柜 美國(guó)Labconco公司;MLS-3020高壓蒸汽滅菌機(jī) 日本三洋公司;Biofuge Stratos冷凍高速離心機(jī)美國(guó)Thermo公司。
1.3 方法
1.3.1 菌落分離和純化
購(gòu)買鮮活鱸魚碎冰猝死,用無菌蒸煮袋包裝,置于4 ℃冰箱冷藏至腐敗,于無菌條件下稱取10 g腐敗魚肉剪碎,加90 mL無菌生理鹽水搖勻制成菌懸液,進(jìn)行梯度稀釋,選擇合適的稀釋度吸取1 mL于無菌平皿中,及時(shí)將15~20 mL平板計(jì)數(shù)瓊脂培養(yǎng)基傾注平皿,并轉(zhuǎn)動(dòng)平皿使其混合均勻,待瓊脂凝固后置于37 ℃進(jìn)行培養(yǎng)。挑選各典型菌落,反復(fù)連續(xù)劃線,得到單菌落。初篩具有群體感應(yīng)現(xiàn)象的菌株。
1.3.2 細(xì)菌16S rRNA序列測(cè)定
取初篩菌株過夜活化,取1 m L新鮮菌液離心(10 000 r/min、10 min),留菌體。按照細(xì)菌基因組D N A快速抽提試劑盒方法提取基因組D N A作為1 6 S r R N A序列擴(kuò)增反應(yīng)模板,2 7 F(5′-AGAGTTTGATCCTGGCTCAG-3′)和1492 R(5′-ACGGCTACCTTGTTACGACTT-3′)為引物進(jìn)行PCR擴(kuò)增。PCR程序:94 ℃、4 min,94 ℃、30 s,58 ℃、30 s,72 ℃、95 s,共30 個(gè)循環(huán)后,72 ℃、10 min。PCR產(chǎn)物進(jìn)行0.1%瓊脂糖凝膠電泳,然后送至生工生物工程(上海)股份有限公司測(cè)序,將測(cè)序結(jié)果與NCBI數(shù)據(jù)庫(kù)進(jìn)行比對(duì)。
1.3.3 AHLs活性檢測(cè)
1.3.3.1 初步檢測(cè)AHLs
平行劃線法[13]:報(bào)告菌株CV026和A136與供試菌株活化后,在LB營(yíng)養(yǎng)瓊脂平板上平行劃線,以報(bào)告菌株做陰性對(duì)照,C6-HSL作陽(yáng)性對(duì)照。28 ℃過夜培養(yǎng),觀察顏色變化。
平板擴(kuò)散法[14]:報(bào)告菌株CV026和A136過夜活化,與LB營(yíng)養(yǎng)瓊脂混勻并倒入已放好牛津杯的平板中,待其凝固后,加入150 μL/孔的AHLs粗提物,28 ℃恒溫培養(yǎng)24 h,觀察顏色變化。
1.3.3.2 氣相色譜-質(zhì)譜(gas chromatography-mass spectrometry,GC-MS)聯(lián)用技術(shù)定性檢測(cè)AHLs
分別將6 種AHLs標(biāo)準(zhǔn)品(C4-HSL、C6-HSL、C8-HSL、C10-HSL、C12-HSL和C14-HSL)溶于甲醇中配制為質(zhì)量濃度200 μg/mL的混合溶液,-20 ℃保存?zhèn)溆茫糜贕C-MS檢測(cè)。
制備AHLs粗提物:參考綦國(guó)紅等[15]方法,將供試菌株在LB液體培養(yǎng)基中28 ℃、160 r/min振蕩培養(yǎng)至對(duì)數(shù)期,離心(10 000 r/min、10 min)取上清液,用含0.01%冰乙酸的乙酸乙酯溶液等體積混合萃取3 次,混合有機(jī)相旋蒸至干,甲醇溶解,-20 ℃保存?zhèn)溆谩?/p>
1.3.4 不同培養(yǎng)條件下產(chǎn)AHLs能力檢測(cè)
將H. alvei-14和S. marcescens-01活化后,分別接種到10 mL LB培養(yǎng)基中,并在28 ℃振蕩(160 r/min)培養(yǎng)到光密度(OD595nm)為0.5左右。然后按S. marcescens-01和H. alvei-14體積比1∶1和1∶3混勻后以1%的總接種量接種在不同液體培養(yǎng)基中,28 ℃、160 r/min搖床培養(yǎng)24 h后,取200 μL培養(yǎng)菌液測(cè)定AHLs誘導(dǎo)的變色圈直徑(mm),每組3 個(gè)平行。
不同液體培養(yǎng)基分別為:不同NaCl質(zhì)量分?jǐn)?shù)(0%、0.5%、1%、1.5%、2%、3%、4%)的液體培養(yǎng)基,不同pH值(5、6、7、8、9)的LB液體培養(yǎng)基,不同碳源(乳糖、蔗糖添加量為5.0 g/L,葡萄糖、果糖、木糖、麥芽糖添加量為5.26 g/L)的AB培養(yǎng)基(3.0 g/L K2HPO4,1.0 g/L NaH2PO4,1.0 g/L NH4Cl,0.3 g/L MgSO4·7H2O,0.15 g/L KCl、0.01 g/L CaCl2,0.002 5 g/L FeSO4·7H2O,酪蛋白水解氨基酸5.0 g/L),不同氮源(氯化銨、胰蛋白胨、酵母浸粉、尿素)的液體培養(yǎng)基。
1.3.5 S. marcescens-01生物被膜的形態(tài)觀察
將菌懸液稀釋100 倍添加在滅菌后的培養(yǎng)皿中,然后分別放入打磨后的鋅片,28 ℃靜置培養(yǎng)適宜的時(shí)間后取出鋅片,用無菌蒸餾水沖洗數(shù)遍,置于4 ℃預(yù)冷的2.5%戊二醛溶液中浸泡4 h,進(jìn)而梯度乙醇脫水,再經(jīng)醋酸異戊酯置換2 次,15 min/次,取出后放入紫外滅菌后的超凈臺(tái)內(nèi)自然風(fēng)干。樣品噴金后,經(jīng)過掃描電子顯微鏡觀察。
1.3.6 不同培養(yǎng)條件下生物被膜的培養(yǎng)及測(cè)定
將H. alvei-14和S. marcescens-01活化后,分別接種到10 mL LB培養(yǎng)基中,并在28 ℃振蕩(160 r/min)培養(yǎng)到光密度(OD595nm)為0.5左右。H. alvei-14和S. marcescens-01用不同液體培養(yǎng)基(同1.3.4節(jié)方法)稀釋100 倍后,按S. marcescens-01與H. alvei-14體積比1∶1和1∶3混勻后在96 孔板中28 ℃培養(yǎng),每組3 個(gè)平行,以液體培養(yǎng)基為對(duì)照。培養(yǎng)48 h后采用結(jié)晶紫染色的方法在595 nm波長(zhǎng)處測(cè)定光密度。
1.4 數(shù)據(jù)處理
2 結(jié)果與分析
2.1 16S rRNA序列分析與系統(tǒng)發(fā)育樹的構(gòu)建
圖1 菌株H. alvei- 14和S. marcescens-01的16S rRNA PCR擴(kuò)增產(chǎn)物電泳圖Fig. 1 Electrophorograms of 16S rRNA PCR ampli fi cation products from H. alvei-14 and S. marcescens-01
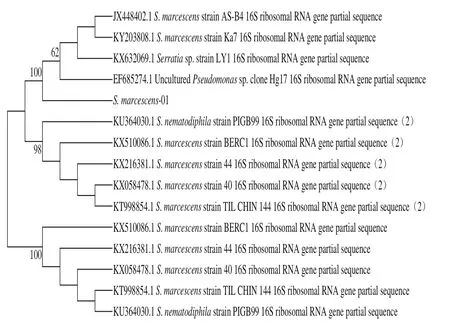

圖2 基于16S rRNA基因序列H. alvei- 14和S. marcescens-01的系統(tǒng)發(fā)育樹Fig. 2 Phylogenic tree based on 16S rRNA sequence analysis of H. alvei-14 and S. marcescens-01
首先以菌株H. alvei-14和S. marcescens-01的總基因組DNA為模板,以細(xì)菌的通用引物擴(kuò)增16S rRNA序列,經(jīng)PCR特異性擴(kuò)增,對(duì)所獲得的PCR產(chǎn)物進(jìn)行測(cè)序(圖1)。所測(cè)得的菌株H. alvei-14和S. marcescens-01的16S rRNA序列與NCBI數(shù)據(jù)庫(kù)進(jìn)行BLAST比對(duì),發(fā)現(xiàn)與蜂房哈夫尼亞菌和粘質(zhì)沙雷氏菌的相似度高達(dá)99%。利用MEGA 5.0軟件構(gòu)建系統(tǒng)發(fā)育樹如圖2所示。
2.2 AHLs檢測(cè)結(jié)果

表1 平行劃線法和瓊脂擴(kuò)散法對(duì)AHLs的檢測(cè)Table 1 Identification of AHLs production by parallel streak and agar diffusion methods
由表1可知,供試菌株都能誘導(dǎo)CV026變紫色,誘導(dǎo)A136水解X-gal產(chǎn)生藍(lán)色。H. alvei-14分泌AHLs的能力較S. marcescens-01強(qiáng)。
2 種菌株對(duì)不同酰基側(cè)鏈長(zhǎng)度及不同取代基AHLs敏感性不同,平行劃線和瓊脂擴(kuò)散法檢測(cè)與樣品中信號(hào)分子提取量和濃度有關(guān),僅用于AHLs初步分析,不能確定AHLs種類和定量分析,而GC-MS可用于多種AHLs的定性和定量檢測(cè),有靈敏性與準(zhǔn)確性更高的優(yōu)點(diǎn)[16]。
AHLs標(biāo)準(zhǔn)品的出峰時(shí)間和順序如圖3A所示,各峰分離良好,峰形尖銳且對(duì)稱,完全分離開來。從左到右依次為C4-HSL、C6-HSL、C8-HSL、C10-HSL、C12-HSL和C14-HSL,保留時(shí)間分別為3.974、5.674、7.629、9.846、12.410、15.398 min。采用選擇離子監(jiān)測(cè)模式對(duì)未知樣品進(jìn)行檢測(cè)。通過菌株H. alvei-14粗提液(圖3B)、菌株S. marcescens-01粗提液(圖3C)的色譜圖與AHLs標(biāo)準(zhǔn)品色譜圖進(jìn)行比較,發(fā)現(xiàn)菌株H. alvei-14粗提液和菌株S. marcescens-01粗提液可分別檢測(cè)到C6-HSL和C4-HSL、C6-HSL。
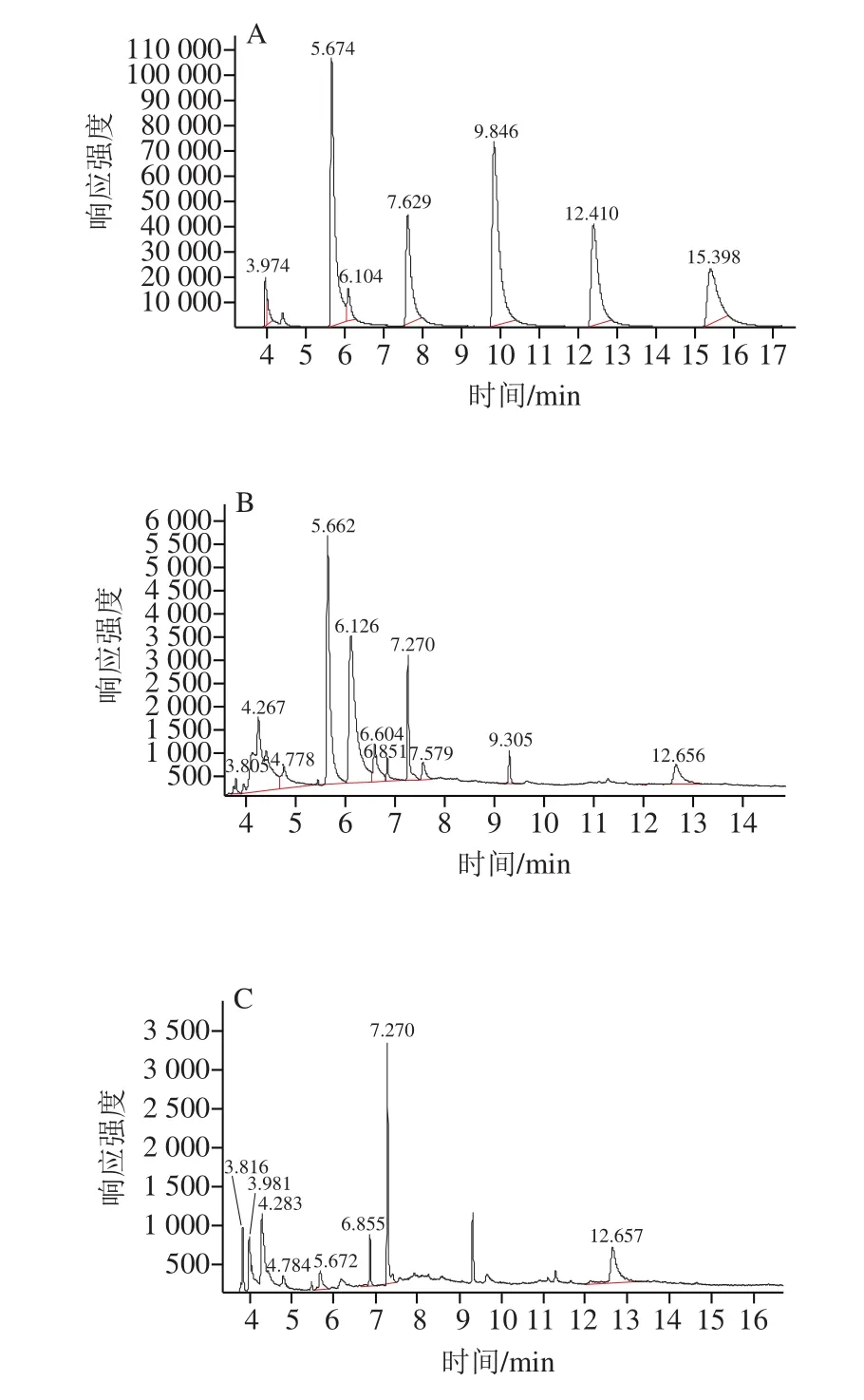
圖3 選擇離子監(jiān)測(cè)模式下標(biāo)準(zhǔn)品(A)、菌株H. alvei-14粗提液(B)、菌株S. marcescens-01粗提液(C)GC-MS圖Fig. 3 GC-MS chromatogram in selected ion monitoring (SIM) mode
2.3 生長(zhǎng)動(dòng)力學(xué)曲線
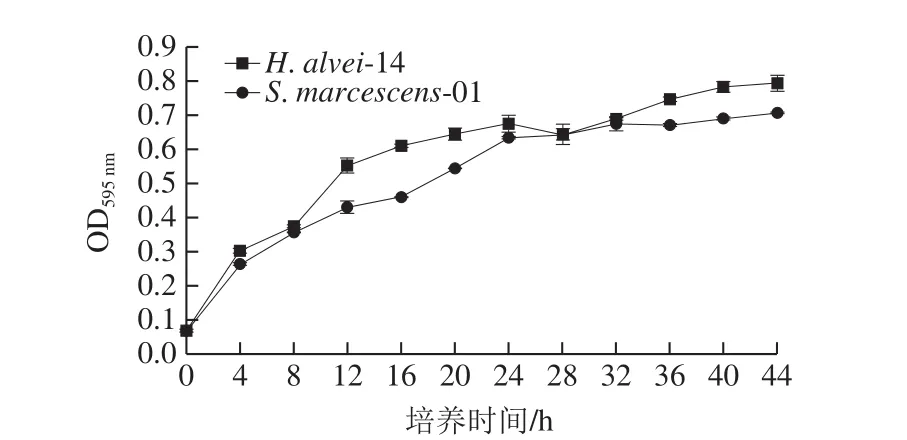
圖4 菌株H. alvei- 14和S. marcescens-01的生長(zhǎng)動(dòng)力學(xué)曲線Fig. 4 Growth curves of H. alvei-14 and S. marcescens-01
圖4 為細(xì)菌的H. alvei-14和S. marcescens-01的生長(zhǎng)動(dòng)力學(xué)曲線,0~4 h菌株呈指數(shù)增長(zhǎng)趨勢(shì),12~24 h菌株不斷生長(zhǎng),菌落數(shù)緩慢上升,24 h后隨著培養(yǎng)時(shí)間的延長(zhǎng),菌株生長(zhǎng)較平穩(wěn)。菌株H. alvei-14生長(zhǎng)能力較S. marcescens-01強(qiáng),但是基本趨勢(shì)相同。后期實(shí)驗(yàn),采用OD595nm為0.5左右的菌株,此時(shí)菌株處于對(duì)數(shù)期,仍具有較好生長(zhǎng)能力,活性較穩(wěn)定,便于實(shí)驗(yàn)研究。
2.4 不同培養(yǎng)條件下AHLs的分泌
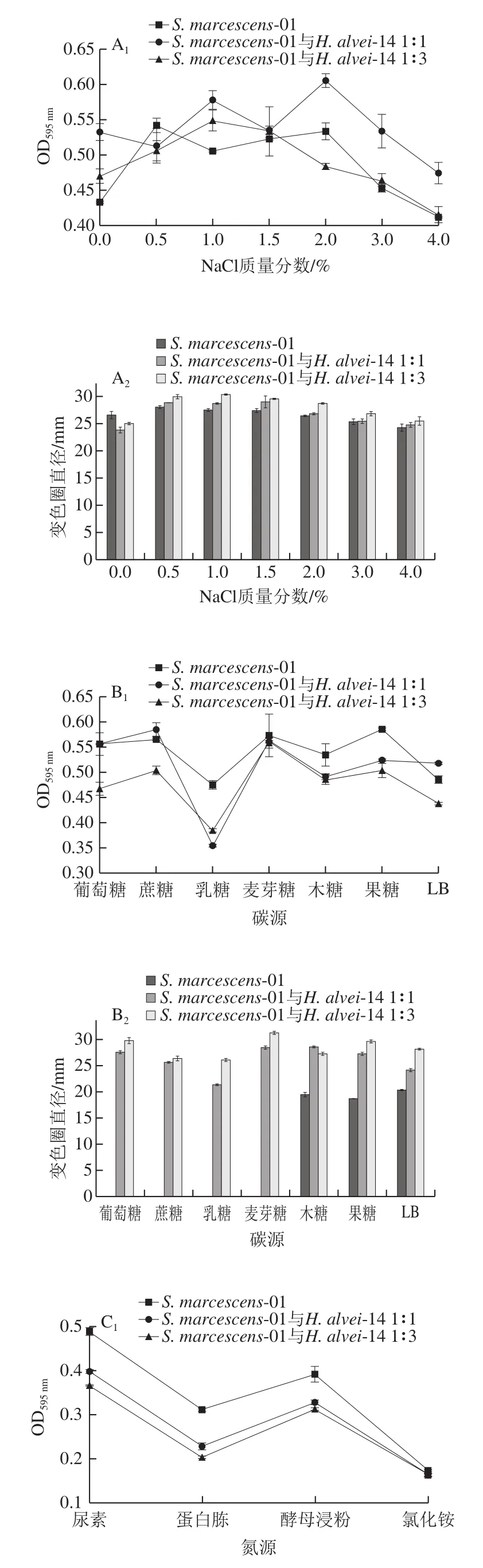
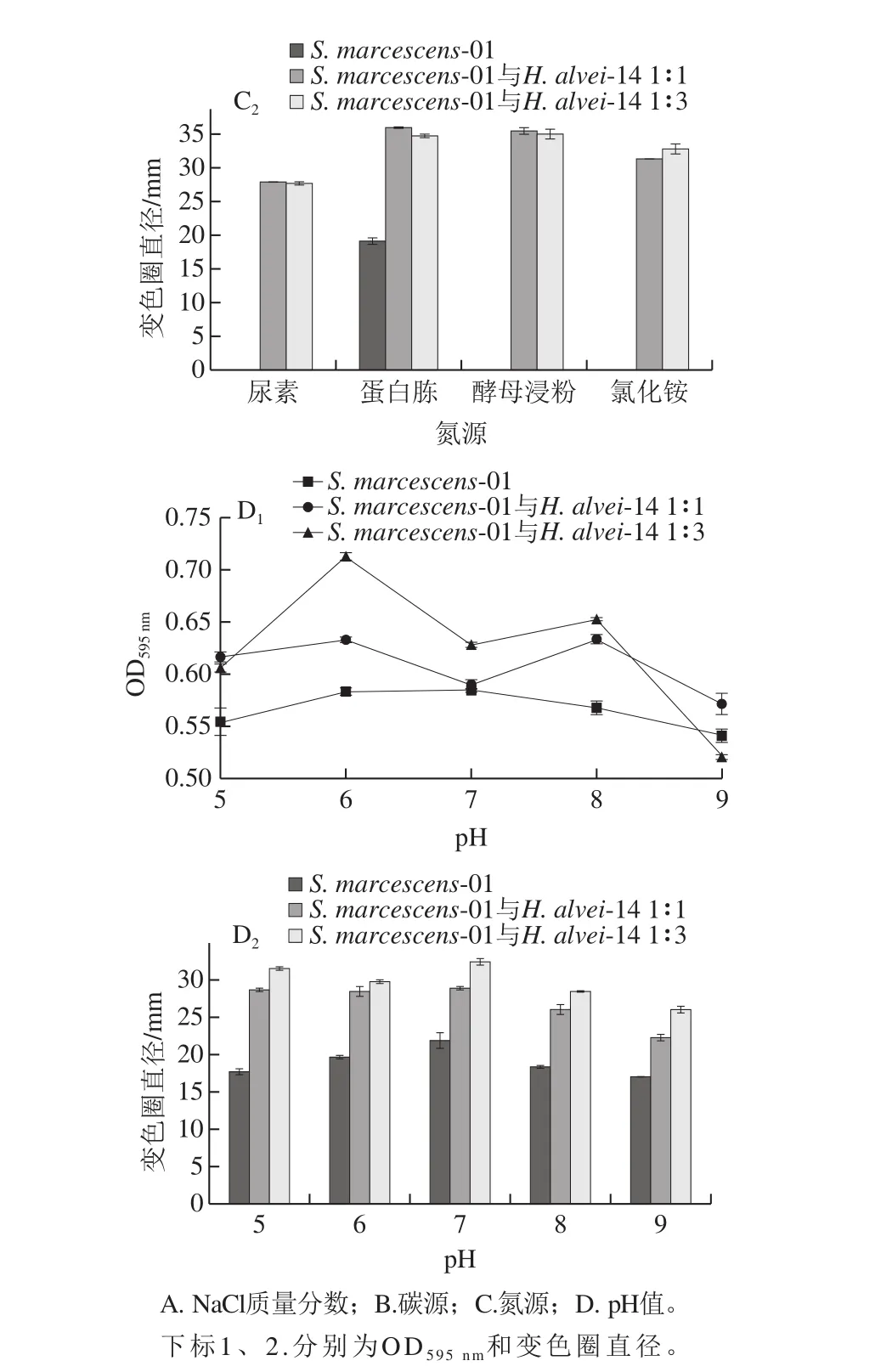
圖5 不同培養(yǎng)條件下AHLs的分泌Fig. 5 Effects of different culture conditions on secretion of AHLs
如圖5A所示,與陰性對(duì)照(NaCl為0%)相比,一定NaCl質(zhì)量分?jǐn)?shù)(0.5%~2%)可刺激菌株分泌AHLs,當(dāng)質(zhì)量分?jǐn)?shù)大于2%時(shí),菌株生長(zhǎng)受到抑制,AHLs產(chǎn)量明顯減少。與對(duì)照S. marcescens-01相比,共培養(yǎng)體系在各質(zhì)量分?jǐn)?shù)下變色圈直徑均變大,說明共培養(yǎng)后促進(jìn)AHLs的分泌。Ravn等[17]研究表明,當(dāng)NaCl質(zhì)量分?jǐn)?shù)為3%時(shí),變形斑沙雷氏菌B5a的信號(hào)分子分泌量減少,而成團(tuán)腸桿菌B6a的信號(hào)分子分泌量會(huì)增加,說明NaCl的質(zhì)量分?jǐn)?shù)會(huì)對(duì)菌株分泌信號(hào)分子產(chǎn)生不同程度的影響。
如圖5B所示,以LB為對(duì)照,S. marcescens-01誘導(dǎo)CV026紫色圈直徑僅達(dá)到20.35 mm;且僅以木糖、果糖為碳源時(shí)才能檢測(cè)到AHLs。共培養(yǎng)體系以葡萄糖、果糖、木糖、麥芽糖為碳源時(shí),產(chǎn)AHLs的能力較強(qiáng);而對(duì)乳糖、蔗糖利用程度較低,產(chǎn)AHLs的能力較弱。Geisenberger等[18]對(duì)于銅綠假單胞菌的研究表明,在補(bǔ)充添加0.2%葡萄糖作為唯一碳源時(shí)分泌的AHLs種類和含量最高。說明碳源不同,導(dǎo)致菌株分泌信號(hào)分子的能力不同。
如圖5C所示,S. marcescens-01僅以蛋白胨為氮源時(shí)才能檢測(cè)到AHLs,變色圈直徑為19.01 mm。共培養(yǎng)體系以尿素為氮源,菌液密度較大,但分泌信號(hào)分子的能力較差;以蛋白胨和氯化銨為氮源,菌體密度較小,但分泌AHLs的量較多。Dandekar等[19]研究比較了380 個(gè)可溶氮源中的銅綠假單胞菌AHL合成突變體加或減對(duì)AHL的生長(zhǎng),發(fā)現(xiàn)在以Glu-Tyr、His-Trp、Tyr-Gly-Gly、Tyr-Phe、Tyr-Trp和Tyr-Tyr作為氮源的信號(hào)存在下,生長(zhǎng)得到改善。說明在特定氮源存在下,可改善菌株生長(zhǎng),進(jìn)而調(diào)控AHLs的分泌。
如圖5D所示,共培養(yǎng)體系在不同pH值下產(chǎn)AHLs能力的趨勢(shì)與S. marcescens-01相似,在pH 7時(shí)分泌信號(hào)分子的能力最強(qiáng),變色圈直徑達(dá)到最大,pH過低或過高時(shí),菌株生長(zhǎng)未受到明顯影響,但AHLs分泌能力下降。
2.5 不同時(shí)間S. marcescens-01生物被膜掃描電子顯微鏡觀察
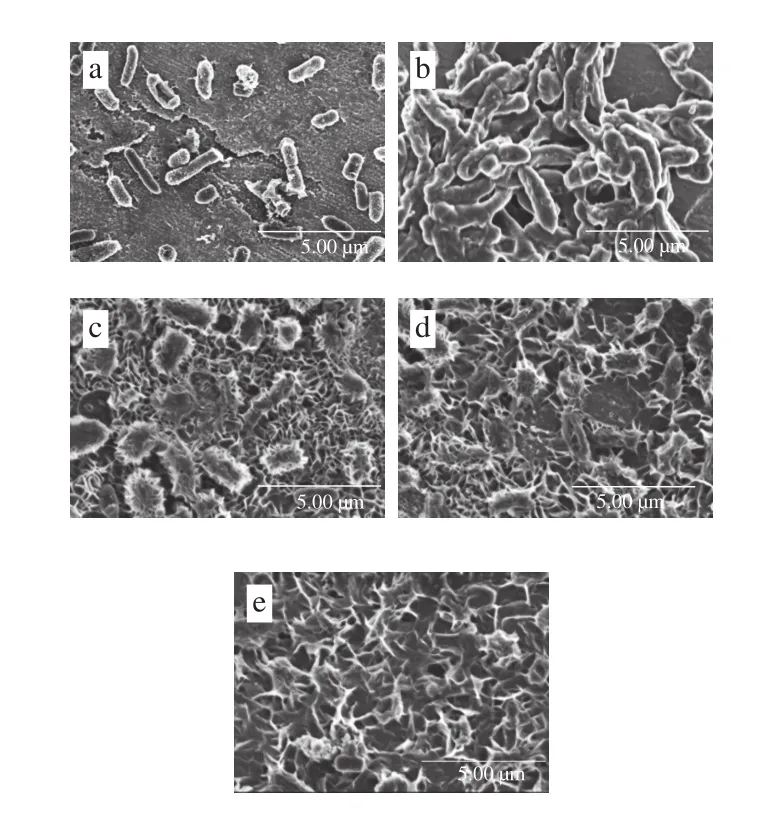
圖6 不同培養(yǎng)時(shí)間下S. marcescens-01生物被膜形態(tài)(×10 000)Fig. 6 Bio fi lm morphology of S. marcescens-01 at different culture times ( × 10 000)
生物被膜是指菌體黏附于載體表面,分泌黏性的胞外多聚物,并與其他成分組成胞外基質(zhì)將其自身包圍其中而形成復(fù)雜三維結(jié)構(gòu),能使細(xì)菌高度耐藥,抵抗不利環(huán)境[20]。細(xì)菌生物被膜的形成是一個(gè)動(dòng)態(tài)過程。細(xì)菌的生物被膜一般會(huì)經(jīng)歷:1)初始附著期;2)不可逆附著期;3)初始形成和發(fā)展期;4)成熟期;5)脫落期。如圖6所示,在12 h菌體開始聚集黏附在鋅片表面;隨著時(shí)間的延長(zhǎng),鋅片表面附著的菌體數(shù)目逐漸增多,細(xì)菌之間開始黏結(jié),且胞外聚合物將菌體包裹其中,進(jìn)而生成成團(tuán)聚集的生物被膜,36~60 h是生物被膜形成的發(fā)展成熟和穩(wěn)定階段。
2.6 不同培養(yǎng)條件下生物被膜的形成
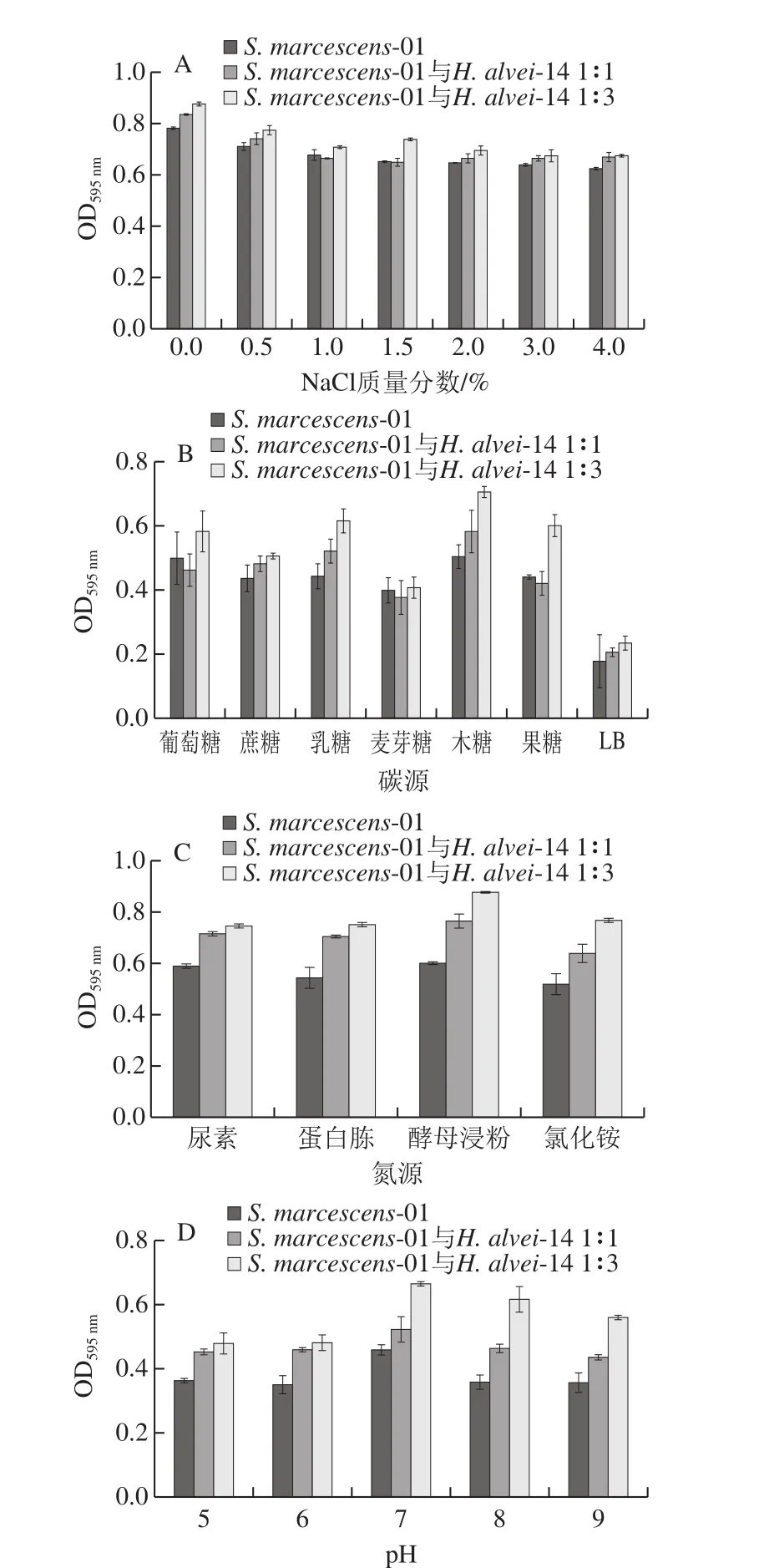
圖7 不同培養(yǎng)條件對(duì)生物被膜的影響Fig. 7 Effect of different culture conditions on the formation of biofilm
如圖7A所示,隨著NaCl質(zhì)量分?jǐn)?shù)的增加,細(xì)菌細(xì)胞因滲透壓過高而脫水,導(dǎo)致其失水死亡,進(jìn)而抑制生物被膜的形成。H. alvei-14與S. marcescens-01共培養(yǎng),生物被膜形成能力增強(qiáng),S. marcescens-01與H. alvei1∶3產(chǎn)生物被膜的能力強(qiáng)于共培養(yǎng)體系S. marcescens-01與H. alvei1∶1體系。
如圖7B所示,以LB培養(yǎng)基為對(duì)照,添加碳源促進(jìn)了菌株S. marcescens-01生物被膜的形成。共培養(yǎng)體系以木糖、乳糖為碳源,生物被膜生成量最多,以麥芽糖為碳源時(shí),生物被膜生長(zhǎng)較差。劉晶晶等[21]研究發(fā)現(xiàn),在25 ℃條件下,麥芽糖和乳糖質(zhì)量分?jǐn)?shù)由0.2%增至1.4%時(shí),金黃色葡萄糖球菌-大腸桿菌混合生物被膜形成較快。
如圖7C所示,S. marcescens-01與共培養(yǎng)體系均以酵母浸粉為氮源時(shí),生物被膜產(chǎn)生量最多,而以氯化銨為氮源時(shí),生物被膜量形成較少。不同氮源會(huì)使菌的生長(zhǎng)速率出現(xiàn)差異,并進(jìn)一步對(duì)生物膜的黏附和代謝活性等產(chǎn)生影響[22]。
由圖7D所示,生物被膜生長(zhǎng)的最佳pH值都為7,生長(zhǎng)環(huán)境中pH值過低或過高均不利于微生物生物被膜的形成。
3 討 論
共存體系中微生物互相作用是一個(gè)復(fù)雜的過程,不僅存在菌落間競(jìng)爭(zhēng)營(yíng)養(yǎng)物質(zhì),還存在代謝產(chǎn)物對(duì)彼此菌體的影響[23]。與對(duì)照S. marcescens-01相比,2 個(gè)共培養(yǎng)體系在各不同培養(yǎng)條件下變色圈直徑變大,說明共培養(yǎng)后促進(jìn)AHLs的分泌,其原因可能在于H. alvei-14的存在被S. marcescens-01認(rèn)為是一種環(huán)境刺激,其通過某些調(diào)控機(jī)制調(diào)節(jié)自身AHLs的合成,這種調(diào)控機(jī)制可能包括對(duì)H. alvei-14的感知和對(duì)AHLs合成的調(diào)控。環(huán)境條件的改變會(huì)影響AHL的產(chǎn)生量,即在環(huán)境中可能存在影響AHL積累的因素,或者是存在一些因素抑制與AHL合成相關(guān)基因的表達(dá)。鈉離子不參與細(xì)胞的組成,但仍是微生物發(fā)酵培養(yǎng)基的必需成分,鈉離子與維持細(xì)胞滲透壓有關(guān),在培養(yǎng)基中常常加入少量鈉鹽,但是用量不能過高,否則會(huì)影響微生物的生長(zhǎng),氯離子在一般微生物中不具有營(yíng)養(yǎng)作用,但是對(duì)某些微生物生長(zhǎng)是必需的[24]。低NaCl含量和高NaCl含量均會(huì)抑制AHLs的分泌[25]。糖類在微生物正常生長(zhǎng)有著重要作用,一般來說微生物對(duì)單糖的利用程度優(yōu)于雙糖和多糖[26],可能由于木糖、葡萄糖、果糖均為單糖,共培養(yǎng)體系對(duì)其利用程度較高,產(chǎn)AHLs的能力較強(qiáng);而蔗糖、麥芽糖、乳糖均為雙糖,菌株對(duì)麥芽糖、蔗糖利用能力依次降低,AHLs活性隨著降低;而以乳糖為碳源時(shí),菌株密度較小,但產(chǎn)AHLs的能力則相反,其原因可能為乳糖為菌株的限制性碳源,不利環(huán)境反而促進(jìn)了AHLs的產(chǎn)生;因此在水產(chǎn)品加工過程中,應(yīng)盡量選用蔗糖、乳糖等為原料,以減少由AHLs介導(dǎo)的食品腐敗的發(fā)生。通常,微生物生長(zhǎng)所需的氮源主要分為有機(jī)氮源和無機(jī)氮源兩大類,其中有機(jī)氮源物質(zhì)成分復(fù)雜,主要是蛋白質(zhì)及其降解物,而微生物對(duì)無機(jī)氮源的吸收利用能力較強(qiáng)。菌株對(duì)蛋白胨的吸收利用能力較差,而在脅迫環(huán)境下,菌株刺激自身分泌大量的AHLs,而氯化銨可能是為其限制性氮源不利于菌株生長(zhǎng),但可以刺激菌體群體感應(yīng)系統(tǒng),促進(jìn)AHLs的釋放。相比堿性,菌株在酸性環(huán)境下產(chǎn)AHLs能力較強(qiáng)可能是因?yàn)椴焕h(huán)境下共培養(yǎng)體系感知危機(jī)信號(hào),啟動(dòng)相關(guān)基因表達(dá),刺激群體感應(yīng)應(yīng)急機(jī)制而分泌大量的AHLs,從而達(dá)到保護(hù)自身的目的[27]。
在自然條件下,大多數(shù)生物被膜是由許多微生物組成的混合生物被膜,被膜態(tài)細(xì)菌具有浮游菌不具有的優(yōu)勢(shì),這增加了不同微生物之間的相互作用和胞外分泌物的復(fù)雜性,從而改變生物被膜的生理特性及功能。生物被膜的形成與胞外聚合物的分泌有關(guān),而胞外聚合物的分泌離不開營(yíng)養(yǎng)物質(zhì),所以營(yíng)養(yǎng)物質(zhì)對(duì)細(xì)菌生物被膜的形成至關(guān)重要[28]。Brink等[29]研究報(bào)道,影響益生菌生物被膜形成除去自身基因調(diào)控的因素外,還存在其他環(huán)境因素如培養(yǎng)基中營(yíng)養(yǎng)物質(zhì)的不足及限制性碳源的供給均有利于生物被膜的形成。高質(zhì)量分?jǐn)?shù)NaCl溶液能使細(xì)菌細(xì)胞因滲透壓過高而脫水,抑制其生長(zhǎng)繁殖,進(jìn)而抑制生物被膜的形成。可能由于麥芽糖為雙糖,菌株對(duì)其利用程度較低,生物被膜生長(zhǎng)較差。以氯化銨為氮源時(shí),菌株菌液密度較低,生物被膜生長(zhǎng)較差。pH值一般對(duì)處于生物膜形成初期的浮游菌和周圍的活菌產(chǎn)生影響,通常中性環(huán)境對(duì)生物被膜的形成有利[30]。在酸性條件下,微生物胞外分泌物可能會(huì)發(fā)生變性,而不利于其生物被膜的形成;在堿性條件下,不利于菌體的聚集,進(jìn)而抑制其生物被膜的形成。因此,生長(zhǎng)環(huán)境中pH值過低或過高均不利于微生物生物被膜的形成。
4 結(jié) 論
與蜂房哈夫尼亞菌共培養(yǎng)能促進(jìn)粘質(zhì)沙雷氏菌分泌AHLs和形成生物被膜,這種影響可能與菌群間群體感應(yīng)有關(guān)。環(huán)境條件的改變是影響細(xì)菌分泌AHLs和生物被膜形成的重要影響因子,較高鹽含量、pH值過低或過高均不利于微生物分泌AHLs和生物被膜的形成,而限制性碳源及氮源等脅迫環(huán)境均可以刺激菌體群體感應(yīng)系統(tǒng),促進(jìn)AHLs的釋放和生物被膜的形成。因此在水產(chǎn)品加工過程中應(yīng)選擇更適合的食品加工和貯藏條件,以減少由AHLs介導(dǎo)的食品腐敗的發(fā)生和生物被膜的形成。
猜你喜歡
天天愛科學(xué)(2022年9期)2022-09-15 01:12:54
天天愛科學(xué)(2022年4期)2022-05-23 12:41:48
當(dāng)代水產(chǎn)(2022年3期)2022-04-26 14:26:56
科學(xué)大眾(2021年9期)2021-07-16 07:02:54
小讀者(2021年2期)2021-03-29 05:03:48
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
