腐胺提高高齡小鼠卵母細胞質量及其機制的研究
2019-04-19 06:46:18胡夢婷馬薇薇徐文丹吳畏嚴正杰劉嘉茵劉金勇崔毓桂
生殖醫學雜志 2019年4期
胡夢婷,馬薇薇,徐文丹,吳畏,嚴正杰,劉嘉茵,劉金勇,崔毓桂
(南京醫科大學第一附屬醫院生殖醫學中心,南京 210029)
氧化應激是細胞衰老的共同機制之一。高齡女性卵母細胞質量下降、胚胎發育潛能差,其主要原因就是卵母細胞老化,即卵母細胞氧化應激水平升高、線粒體功能障礙。α硫辛酸、輔酶Q10、白藜蘆醇和半胱氨酸等抗氧化劑能夠降低細胞的氧化應激水平、改善線粒體功能,具有一定的延緩細胞老化的作用,但在卵母細胞的作用有待進一步研究[1-2]。因此,研究卵母細胞老化的機制和尋求新型抗氧化劑,對于改善高齡女性卵母細胞質量、提高妊娠率、改進輔助生殖技術臨床結局具有潛在的意義。腐胺是細胞內由L-精氨酸合成的聚陽離子小分子化合物,鳥氨酸脫羧酶(ODC1)催化L-鳥氨酸脫羧生成腐胺,ODC1是多胺生物合成的限速酶,也是在多胺合成中真核生物酶。研究發現,細胞內腐胺參與控制細胞代謝過程中產生的自由基、抑制細胞膜脂質過氧化[3],在細胞生長、增殖、分化、發育、免疫、遷移、基因調控、DNA穩定性以及蛋白質和核酸合成中發揮著重要作用[4]。腐胺在卵母細胞的作用如何?在本文研究腐胺是否通過抗氧化應激的機制提高體外培養的高齡卵母細胞的質量,以探討其遠期臨床應用的價值。
材料與方法
一、材料
1. 實驗動物:7月齡C56/B6J雌鼠購于北京維通利華生物公司,飼養于南京醫科大學動物中心至9月齡(相當于人類女性38~40歲年齡)。光照周期12 h/12 h;自由進食和飲水。本研究得到南京醫科大學實驗動物倫理委員會審批。
2. 實驗試劑:鳥氨酸脫羧酶(ODC1)催化L-鳥氨酸脫羧生成腐胺,且 ODC1是多胺生物合成的限速酶[5]。α-二氟甲基鳥氨酸(α-difluoromethylornithine,DFMO,sigma公司,美國)是ODC1的特異性、高效抑制劑,可明顯抑制細胞內多胺合成、顯著降低多胺水平,已見于多項研究報道[3,5-6]。孕馬血清促性腺激素(PMSG)(寧波舒生第二激素廠)。M2操作液、人表皮因子(hEGF)、重組人卵泡刺激素(rhFSH)[7-8]、透明質酸酶、腐胺、臺式酸、礦物油、γ-actin抗體(sigma,美國);兔抗大鼠SOD2多克隆抗體,兔抗大鼠SOD1多克隆抗體(proteintech,美國);JC-1熒光探針(Molecular Probes,Eugene,Oregon,美國);PVDF膜(GE Heahhcare,美國);增強化學發光檢測試劑盒(ECL detection kit,Millipore,美國)。
二、實驗方法
1. 卵母細胞體外培養:9月齡C56/B6J雌鼠腹腔注射PMSG 10 U,48 h后斷頸處死。立即取出卵巢,在顯微鏡下用1 ml無菌注射器戳破卵泡,用負壓管虹吸卵丘細胞包裹完整的GV-COCs,平均置入IVM、0.5 mmol/L DFMO、0.5 mmol/L腐胺組中。采用微滴式培養,10枚GV-COCs置于一滴,每滴20 μl,覆蓋2 ml礦物油以避免蒸發。在37℃、5%CO2、21%O2條件下培養16~18 h。
2. 卵母細胞收集:將培養后的COCs置于透明質酸酶中吹打,使包裹的卵丘細胞脫落。轉移卵母細胞,置于M2微滴中,觀察裸卵并計數完成第一次減數分裂及排出第一極體的卵母細胞(即MⅡ期卵母細胞)。
3. 單卵母細胞線粒體拷貝數:卵母細胞的選取及裂解:MⅡ期卵母細胞以臺式酸酸化、吹吸,去除透明帶,PBS液洗滌卵母細胞3次。制備終濃度為30 mmol/L Tris-HCL、5 mmol/L EDTA、10 mmol/L KCL、體積分數 1.5% Tween-20及2 mg/ml 蛋白酶K的細胞裂解液,將卵母細胞移入裝有10 μl 細胞裂解液的 0.2 ml PCR 管中;55℃加熱2 h,95℃10 min,滅活蛋白酶K,裂解卵細胞。然后置于-80℃用于后續PCR擴增模板。
PCR模板制備及引物設計:根據相關文獻,合成引物(由南京銳真生物技術有限公司合成)ND2 Forward 5’-CAC GAT CAA CTG AAG CAG CAA-3’,Reverse 5’-ACG ATG GCC AGG AGG ATA ATT-3’。
標準品制備:1 μl模板,10 μl SYBR Green PCR Master Mix,0.4 μl ROX Reference Dye,引物各0.8 μl,余下體積用雙蒸水補齊至20 μl。反應條件為:95℃預變性5 min:95℃變性30 s,65℃退火30 s,72℃延伸30 s,共40個循環;DNA提純:擴增產物加入500 μl無水乙醇,4℃,12 000 rpm離心10 min,棄上清;加入500 μl 75%乙醇12 000 rpm離心10 min,棄上清,將提純擴增產物進行瓊脂糖凝膠電泳,檢測擴增產物分量,計算出分離純化的DNA樣品即標準品的拷貝數;將標準品按照 1∶10的比例進行等倍比稀釋而形成梯度濃度標準品,標準品等比自稀釋:10-3~10-8。
mtDNA 拷貝數檢測:使用Quanti Nova SYBR Green PCR 試劑盒,在實時熒光定量PCR 擴增儀上檢測mtDNA 拷貝數。根據試劑盒說明書制備20 μl PCR反應體系:1 μl模板,10 μl SYBR Green PCR Master Mix,0.4 μl ROX Reference Dye,上下游引物各0.8 μl,余下體積用雙蒸水補齊至20 μl。反應條件為:95℃預變性5 min:95℃變性30 s,65℃退火30 s,72℃延伸30 s,共40個循環;設定溶解曲線,檢測 PCR 反應特異性。每次反應的標準曲線通過已知拷貝數的標準品確定,每個樣品的起始mt DNA拷貝數通過此標準曲線來確定。
4. 氧化應激水平檢測:檢測高齡卵母細胞的ROS水平。使用50-羧基-20,7-二氟二氫熒光素二乙酸酯(carboxy-DFFDA;Molecular Probes),一種能夠檢測強氧化劑如H2O2和過氧亞硝酸鹽的熒光探針。將去除卵丘細胞的MⅡ期卵母細胞置入預熱10 min的熒光探針微滴中,將培養皿放入培養箱中孵育30 min,然后在預熱的M2操作液中洗滌3次,將其放置于預熱的玻璃底培養皿中進行掃描拍攝。
5. 線粒體膜電位檢測:膜電位敏感染料JC-1用于評估線粒體的功能活性。JC-1是一種廣泛用于檢測線粒體膜電位的理想熒光探針。用DMSO制備2 mM JC-1儲備溶液,保存于-20℃。使用前,將儲備溶液用M2稀釋至工作濃度2 μM。將工作液于培養皿中制成微滴,用礦物油覆蓋防止工作濃度改變,置于培養箱中預熱10 min。將卵母細胞置于微滴中放入培養箱孵育30 min,然后在M2操作液中洗滌3次,置入預熱的玻璃底培養皿微滴中,后進行共聚焦觀察。在相同的暴露強度下捕獲所有卵母細胞。通過測量每個通道中整個卵母細胞的總熒光來進行熒光分析。使用校正的總細胞熒光(CTCF)方法將每個通道的平均灰度值標準化為測量區域。線粒體膜電位為總紅色CTCF除以總綠色CTCF。
6. Western Bolting:收集30枚MⅡ期卵母細胞置于5 μl 蛋白裂解液中,與4~10% SDS-PAGE梯度膠電泳分離,轉移到PVDF膜。用TBS(含5%脫脂奶)37℃處理l h后,加入SOD2抗體(1∶500稀釋)或SOD1抗體(1∶500稀釋)孵育,內參是兔抗鼠γ-actin多克隆抗體(1∶1 000稀釋),4℃過夜。TBST(TBS+0.1%Tween 20)洗脫后,二抗(驢抗兔IgG,1∶1 000稀釋)37℃孵育1 h,TBST洗脫。用增強化學發光檢測試劑盒(ECL)測定熒光強度。條帶灰度值用天能凝膠成像系統軟件(Tanon Gis Image system ID Danalysis,上海)分析。
三、統計學分析
采用SPSS 20.0統計軟件對數據進行分析。計數資料用均數±標準差表示,采用獨立樣本t檢驗進行分析。P<0.05為差異有統計學意義。
結 果
一、腐胺增加高齡卵母細胞的成熟率
三組高齡卵母細胞成熟率比較(圖1)。腐胺組卵母細胞成熟率(87.57±1.70)%,顯著高于對照組(66.02±4.05)%,平均升高了22%(P<0.01);DFMO組卵母細胞成熟率為(66.96±3.40)%,與對照組差異沒有統計學意義(P>0.05)。
二、腐胺減少高齡卵母細胞內活性氧
腐胺減少高齡卵母細胞內的活性氧(圖2)。腐胺組卵母細胞內活性氧水平顯著減少,DFMO組卵母細胞內活性氧水平顯著升高(圖2A)。與對照組(18 714 873±1 641 482)相比,腐胺組活性氧水平(11 827 188±1 290 525)下降了37%(P<0.01);DFMO組(52 119 088±4 178 825)活性氧水平升高179%(P<0.01)(圖2B)。
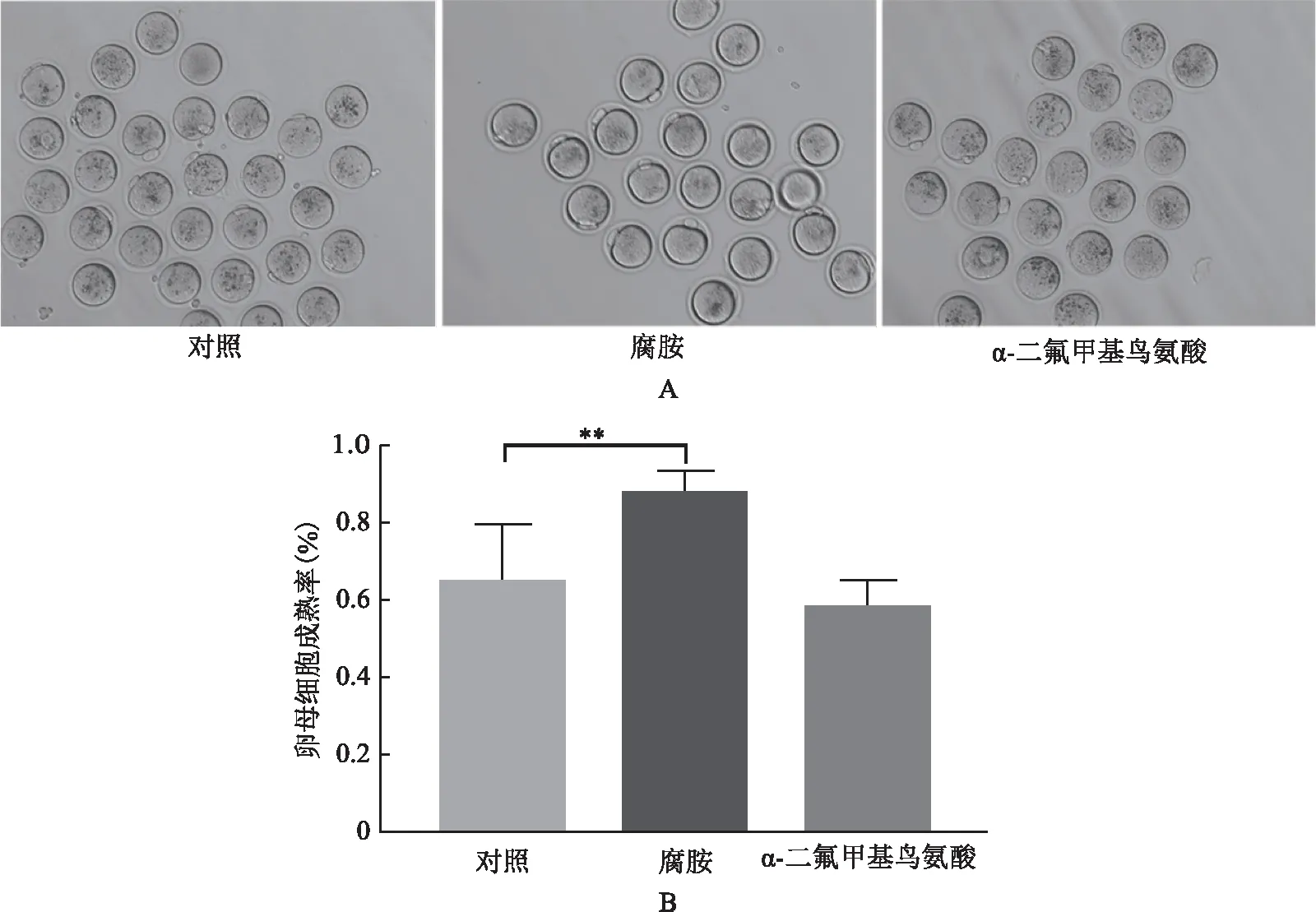
**P<0.01圖1 腐胺提升高齡卵母細胞的成熟率
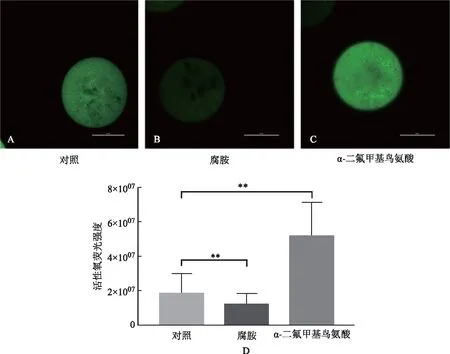
A、B、C:熒光顯示卵母細胞內ROS,carboxy-DFFDA 綠色染料標記氧自由基,標尺50 μm;D:卵母細胞ROS水平統計分析,**P<0.01圖2 腐胺減少高齡卵母細胞內ROS
三、腐胺提升高齡卵細胞內SOD1表達
腐胺提升高齡卵母細胞內SOD1蛋白的表達(圖3A)。腐胺組卵母細胞內SOD1蛋白表達較對照組顯著升高(P<0.05);DFMO組SOD1蛋白表達較對照組顯著下降(P<0.05)(圖3B)。
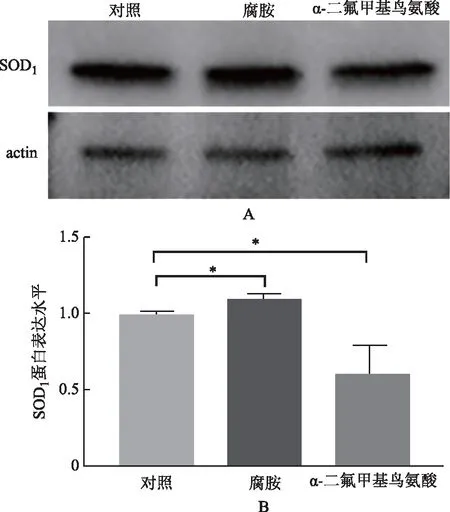
*P<0.05圖3 腐胺提升高齡卵母細胞內SOD1蛋白表達
四、腐胺對高齡卵母細胞線粒體DNA拷貝數的影響
腐胺對高齡卵母細胞線粒體DNA拷貝數的影響(圖4)。以DFMO抑制卵母細胞內腐胺生成,高齡卵母細胞線粒體DNA拷貝數(48 811±1 624)較對照組(35 026±6 402)增加24.4%(P<0.05)。但是,外源性添加腐胺,高齡卵母細胞線粒體DNA拷貝數(42 137±5 772)未發生顯著改變(P>0.05)。

與對照組比較,*P<0.05圖4 抑制腐胺增加高齡卵母細胞線粒體DNA拷貝數
五、腐胺提升高齡卵母細胞線粒體膜電位
線粒體內膜上的質子泵將線粒體基質內的質子泵出膜間腔,使內膜外電位為正、膜內為負,這種膜內外的電位差稱為線粒體跨膜電位(△Ψm),其閾值變化與氧化代謝水平相關,常被用來評估線粒體功能。當線粒體膜電位較高時,JC-1聚集在線粒體的基質中,產生紅色熒光;當線粒體膜電位較低時,JC-1不能聚集在線粒體的基質中,此時JC-1為單體,產生綠色熒光[9-10](圖5A)。與對照組相比(0.62±0.05),腐胺組線粒體膜電位(2.20±0.18)升高253%(P<0.01),DFMO組(0.33±0.05)下降46.7%(P<0.01)(圖5B)。
討 論
卵巢卵母細胞和顆粒細胞內,腐胺可能參與調節卵母細胞發育成熟及排卵[5]。研究發現,在LH作用下卵巢組織ODC1升高,細胞內腐胺水平顯著升高,激發卵母細胞成熟和隨后的卵泡破裂以釋放成熟卵;LH誘導卵巢ODC1活性比FSH的作用更有效,故認為腐胺對圍排卵期的卵泡成熟是必要的[11-12]。有研究提示,高齡雌性小鼠卵巢內ODC1以及腐胺的濃度降低[11-14]。本文研究發現,在IVM培養液中添加腐胺有效提高高齡小鼠卵母細胞成熟率;添加DFMO以抑制ODC1,抑制內源性腐胺的生成,卵母細胞成熟率較腐胺組明顯降低。與對照組比較,培養液中添加DFMO后高齡小鼠卵母細胞成熟率沒有顯著下降,表明高齡小鼠卵母細胞內ODC1活性低、腐胺濃度低,DFMO作用不明顯。
線粒體是卵母細胞質中含量最為豐富的細胞器,其數量、結構和活性在卵母細胞發育成熟過程中發生顯著變化,線粒體拷貝數由原始生殖細胞中不足10個逐漸增加到成熟卵母細胞的105個以上。線粒體產生的 ATP 是卵母細胞、受精卵以及早期胚胎發育的主要能量來源[15-18]。本研究發現,培養液中添加腐胺后高齡小鼠卵母細胞的線粒體拷貝數沒有顯著改變(P>0.05);添加DFMO后,線粒體拷貝數增加,這可能是因為卵母細胞通過代償性增加線粒體數量來平衡細胞內缺乏腐胺導致的線粒體功能不足。近年研究顯示,線粒體膜電位的降低是細胞凋亡的早期事件。卵母細胞線粒體膜電位與其胚胎發育潛能相關[25]。本研究發現,高齡小鼠卵母細胞體外培養液中添加腐胺,線粒體膜電位增加,提高其線粒體功能;DFMO的作用與之相反。說明腐胺提高卵母細胞線粒體膜電位、有效地改善線粒體功能。
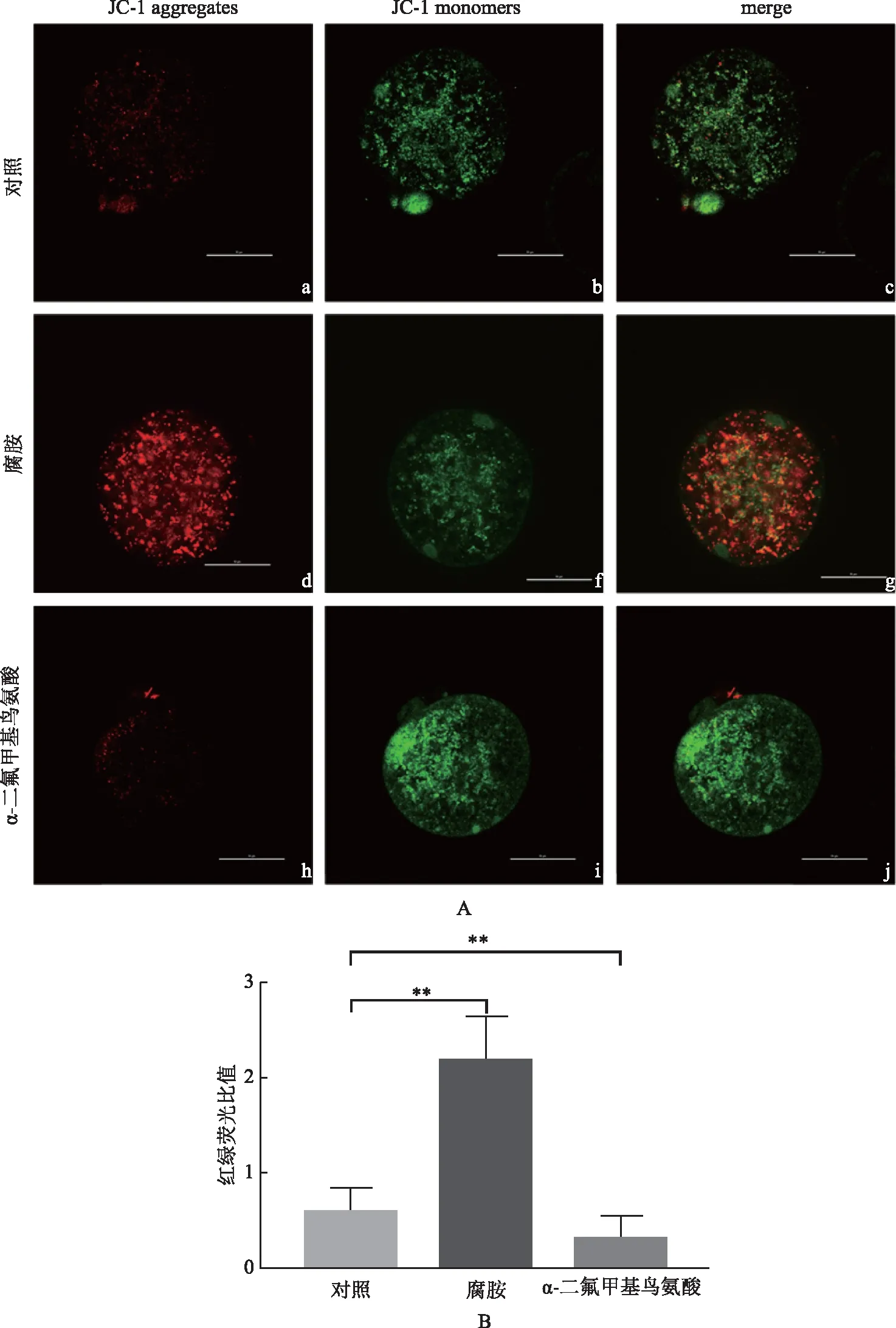
A:高齡卵母細胞線粒體膜電位的觀察,用JC-1染料檢測線粒體膜電位,紅色表示高線粒體膜電位,綠色表示低線粒體膜電位,標尺50 μm;B:卵母細胞線粒體膜電位統計分析;**P<0.01圖5 腐胺提升高齡卵母細胞線粒體膜電位
出生前卵母細胞阻滯在第一次減數分裂前期的雙線期,這一階段稱為生發泡(GV)。人GV期卵母細胞可在卵泡內停留40余年[19-20]。未成熟卵母細胞通過氧化磷酸化維持低水平的ATP,滿足卵母細胞對能量的需求;而隨著卵母細胞發育成熟,對能量需求大大增加。卵母細胞在氧化磷酸化產生ATP過程中,也產生副產物ROS。正常情況下,細胞內ROS和抗氧化之間維持動態平衡,使細胞內ROS保持一定的低水平。當線粒體功能不足,這種平衡被破壞,ROS升高,進而影響卵母細胞發育成熟、受精、胚胎發育[21-22]。有研究顯示,高齡卵母細胞內ROS水平增加,胚胎發育潛能降低[21,23]。本研究發現,體外培養高齡卵母細胞時添加腐胺,顯著減少細胞內ROS;而DFMO使ROS水平明顯增加。在細菌以及細胞相關實驗中,腐胺作為細胞內的抗氧化劑可以保護高氧化應激環境中的細菌和細胞[24]。卵母細胞的成熟依賴于活性氧與抗氧化之間的氧化還原平衡。哺乳動物細胞表達3種亞型的超氧化物歧化酶(SOD1-3)。本文實驗中,腐胺并不影響卵母細胞內SOD2水平,與文獻報道一致[26]。有文獻報道,老齡小鼠卵巢內SOD1水平降低[27];本研究發現,腐胺有效地提高卵母細胞內SOD1表達,DFMO抑制腐胺生成時明顯減少SOD1表達,表明腐胺也能夠有效地提高卵母細胞內SOD1表達,發揮抗氧化的作用,有助于達成卵母細胞內氧化-還原反應的平衡。
綜上所述,腐胺通過改善線粒體功能、以及多重抗氧化的機制,促進卵母細胞成熟,提高高齡卵母細胞質量和胚胎發育潛能。我國目前放開了二胎政策,越來越多高齡女性有生育需求,但是高齡女性的生育結局較低[18,28]。與年齡相關的女性生育能力下降主要歸因于卵巢衰老引起的卵母細胞質量下降[29]。本研究表明,腐胺改善高齡卵母細胞的質量,對改善高齡婦女的生育結局具有潛在的臨床意義。
