Wfs1基因敲除小鼠肝臟芯片數據的生物信息學分析
2019-04-29 02:09:36胡瑞瑋陳淑芹白寧寧張菁高新雨嚴寒方啟晨
江蘇大學學報(醫學版) 2019年2期
胡瑞瑋,陳淑芹,白寧寧,張菁,高新雨,嚴寒,方啟晨
(上海交通大學附屬第六人民醫院糖尿病研究所,上海200233)
WFS1基因于1998年由Inoue等首次發現,定位于染色體4p16,由8個外顯子組成,長33.4 kb[1]。WFS1基因編碼產物主要為內質網上的跨膜糖蛋白wolframin,該蛋白由890個氨基酸組成,相對分子質量約100 kD,含9個跨膜區,在心臟、腦、胰腺等組織有較高水平表達[2]。WFS1基因突變可導致Wolfram綜合征,臨床癥狀主要表現為尿崩癥、糖尿病、視神經萎縮和耳聾,糖尿病通常為首發癥狀[3]。Wolfram綜合征患者的壽命中位數為30歲,60%在35歲以前死亡[4]。目前WFS1基因突變的致病機制尚未完全闡明,因此,本研究對Wfs1基因敲除小鼠肝臟的芯片數據進行生物信息學分析,以探討WFS1基因突變可能的致病機制。
1 資料與方法
1.1 資料和數據
從美國國立生物技術信息中心的Gene Expression Omnibus(GEO)數據庫(https:∥www.ncbi.nlm.nih.gov/geo/)下載基因表達譜芯片數據GSE55143。該數據由Punapart等[5]于2014年上傳至GEO數據庫。該數據集使用Mouse Gene 1.0 ST(Affymetrix公司,美國)表達譜芯片。從中選取野生型和Wfs1敲除小鼠(16~18周齡雄性C57BL/6小鼠)肝臟組織RNA的全基因表達譜進行差異表達基因分析,兩組各有8份重復數據。
1.2 數據處理及差異表達基因分析
通過GEO數據庫基于R語言的交互式網絡分析工具GEO2R(http:∥www.ncbi.nlm.nih.gov/geo/geo2r/),分析GSE55143中野生型和Wfs1敲除小鼠肝臟組織中的差異表達基因。將P<0.05、|logFC|>log21.5[logFC為log2(fold change)]作為差異表達基因篩選標準。
1.3 差異表達基因的富集分析
采用上海伯豪生物技術有限公司的在線富集分析網站(http:∥enrich.shbio.com/index/ga.asp)對差異表達基因進行GO(Gene Ontology)分析以及KEGG(Kyoto Encyclopedia of Genes and Genomes)通路富集分析。采用Fisher精確檢驗,挑選標準為差異表達基因數目≥4,P<0.05,錯誤發現率(false discovery rate,FDR)<0.05。
1.4 差異表達基因的蛋白質相互作用網絡分析
將所有差異基因導入到STRING 10.5(Search Tool for the Retrieval of Interacting Genes/Proteins)[6]在線工具中(https:∥string-db.org/),并用Cytoscape軟件進行可視化[7],獲得蛋白質相互作用網絡圖,置信度閾值為0.4,以連接度大于10作為核心基因篩選標準。
2 結果
2.1 差異表達基因的篩選
通過GEO2R對GSE55143中野生型和Wfs1敲除小鼠肝臟組織RNA的全基因表達譜進行差異表達基因分析,篩選出198個差異表達基因。其中,與野生型相比,Wfs1敲除小鼠肝臟組織中有96個上調基因,102個下調基因。
2.2 差異表達基因的GO分析
為進一步了解這些差異表達基因的功能,對上調及下調基因進行GO富集分析,得到生物學過程、分子功能、細胞組分這3個方面的分析結果。按富集程度由高到低排序,取前5位列于表1中。富集程度定義:(某個詞條中的差異基因數目/總的差異基因數目)/(數據庫該詞條中總的基因數目/數據庫中總的基因數目)。上調基因主要涉及有機陰離子跨膜轉運體活性等生物學過程和分子功能,而下調基因主要涉及芳香化酶活性等分子功能。
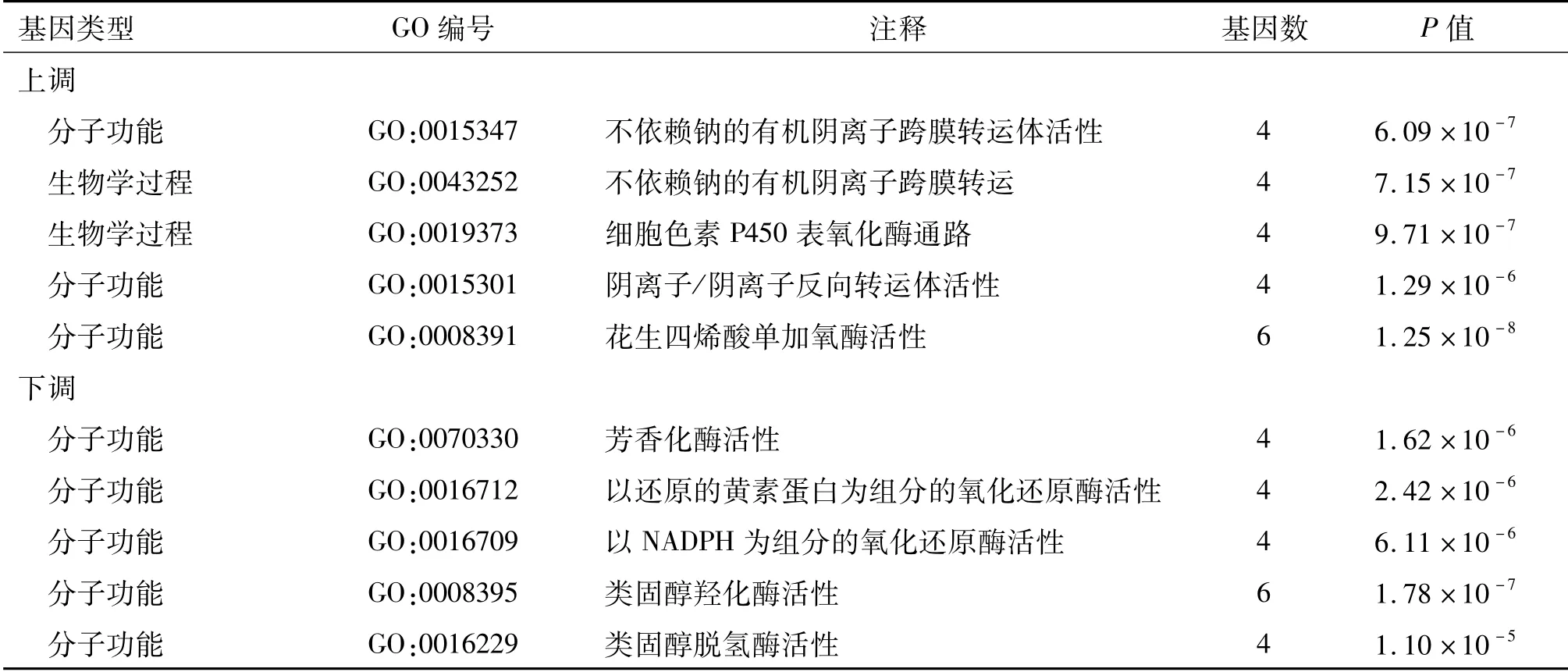
表1 差異表達基因的GO富集分析結果
2.3 差異表達基因的KEGG分析
對上調及下調基因進行KEGG通路富集分析后發現,上調基因有17條通路富集,而下調基因有9條通路富集。按富集程度由高到低排序,取前5位列于表2中。
2.4 差異表達基因的核心基因篩選
通過STRING及Cytoscape軟件對差異表達基因進行分析,獲得蛋白質相互作用網絡圖(圖1)。結果顯示蛋白質相互作用網絡中共有180個節點,最大連接度為35,最小為1,共有26個核心基因。圓形節點和連線分別代表基因編碼的蛋白質和蛋白質間的相互作用,相互作用蛋白的數目和相互作用的綜合評分則分別通過圓圈大小和連線粗細表示。以相互作用蛋白的數目排序,前10位基因為Hsd3b5、Cyp2b9、Abcg2、Cyp4a12a、Cyp2b13、Lepr、Acaca、Cyp17a1、Cyp7a1、Cyp2c38。
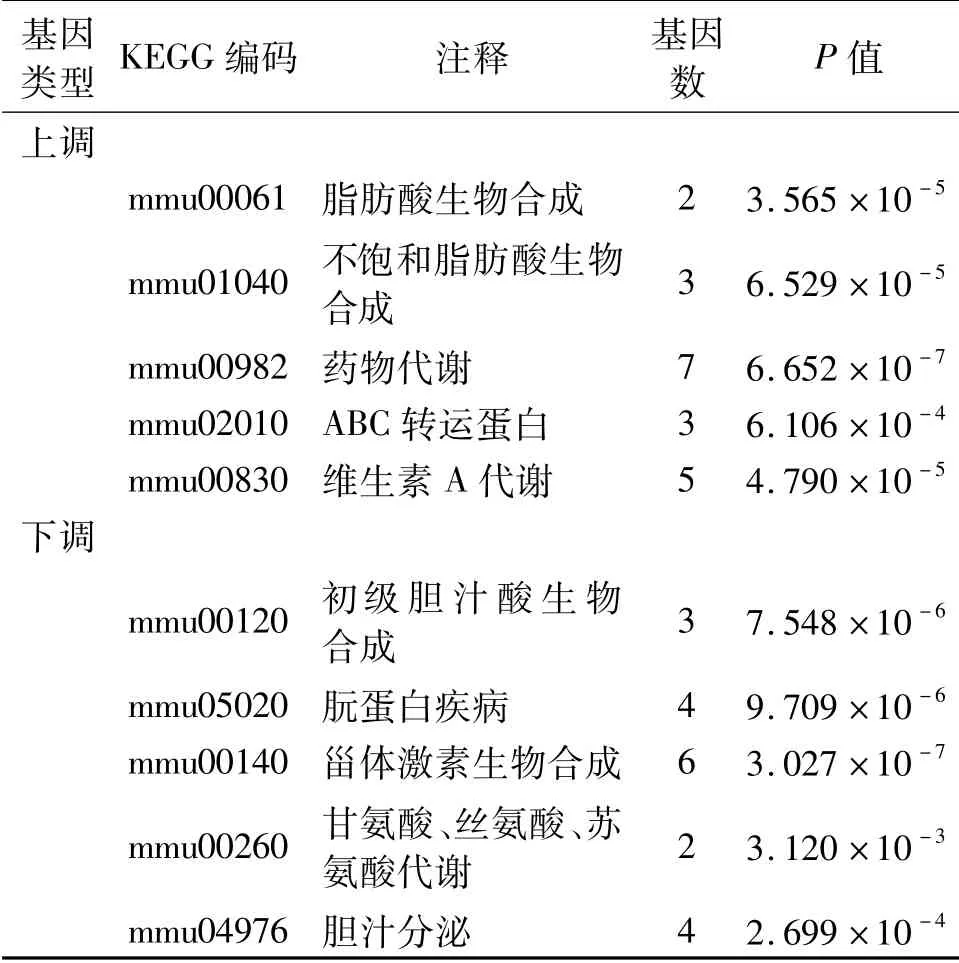
表2 差異表達基因的KEGG富集分析結果
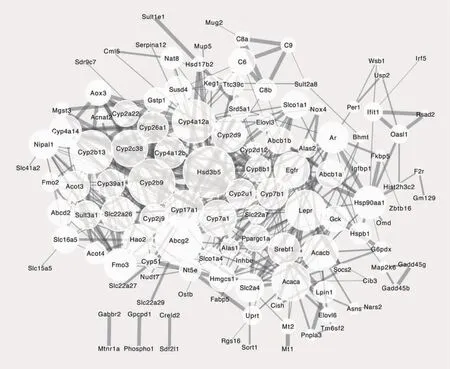
圖1 蛋白質相互作用網絡圖
3 討論
Wfs1基因敲除小鼠表現為血漿胰島素水平低下,葡萄糖耐量受損[8]。然而,只有雄性Wfs1敲除小鼠表現出明顯的高血糖以及體重減輕[9]。雜合子小鼠在體重和血糖水平方面與野生型之間并無顯著差異[5]。此外,Wolfram綜合征患者絕大多數都是純合突變或是復合雜合突變[4]。代謝組學分析發現,與野生型小鼠相比,Wfs1敲除小鼠胰腺和肝臟有更明顯的代謝改變[10]。肝臟是主要的代謝器官,發揮重要的生理及生化功能。因此,本研究對雄性Wfs1敲除小鼠的肝臟組織芯片數據進行挖掘,篩選出198個差異表達基因,其中有96個上調基因,102個下調基因。通過GO、KEGG富集分析,以及STRING、Cytoscape軟件對差異表達基因進行分析后發現,差異表達基因主要集中在脂肪酸生物合成和初級膽汁酸生物合成這兩個方面。
脂肪酸生物合成通路表達增高主要體現在乙酰輔酶A羧化酶1基因Acaca(t=2.704,P=1.47×10-2,logFC=0.715)、乙酰輔酶A羧化酶2基因Acacb(t=2.201,P=4.13×10-2,logFC=0.633)表達的上升。乙酰輔酶A羧化酶1催化乙酰輔酶A生成丙二酰輔酶A,參與脂肪酸合成;而乙酰輔酶A羧化酶2催化乙酰輔酶A生成的丙二酰輔酶A發揮對肉堿棕櫚酰轉移酶1的抑制作用,從而抑制乙酰輔酶A進入線粒體進行β-氧化[11]。ACACA和ACACB基因的表達受到碳水化合物反應元件結合蛋白(carbohydrate response element-binding protein,ChREBP)、固醇調節元件結合蛋白-1(sterol regulatory binding protein-1,SREBP-1)等多種轉錄因子的調控[11]。高血糖可以增加ChREBP的N-乙酰葡萄糖胺修飾[12],從而提高ChREBP的蛋白穩定性和對下游靶基因的轉錄活性[13]。胰島素和胰島素樣生長因子-1(insulin-like growth factor-1,IGF-1)可以通過磷脂酰肌醇-3-激酶/蛋白激酶B通路,激活SREBP-1蛋白的轉錄活性[14]。Wfs1敲除小鼠胰島素分泌障礙[15],但IGF-1表達增高[16]。本研究發現ChREBP、SREBP-1的編碼基因Mlxipl(t=2.651,P=1.65×10-2,logFC=0.303)、Srebf1(t=3.748,P=1.52×10-3,logFC=0.680)在Wfs1敲除小鼠中均表達增高,提示在Wfs1敲除小鼠中,高血糖和IGF-1等代謝變化可能通過ChREBP、SREBP-1等轉錄因子促進脂肪酸合成通路表達增高。
初級膽汁酸由兩條途徑合成,經典的合成通路通過膽固醇7α-羥化酶(CYP7A1編碼蛋白)這一限速酶發起,旁路途徑由膽固醇 27α-羥化酶(CYP27A1編碼蛋白)發起,合成膽酸與鵝去氧膽酸這兩種初級膽汁酸[17]。本研究發現Wfs1敲除小鼠初級膽汁酸合成相關基因表現出下調。Cyp7a1(t=-2.546,P=2.05×10-2,logFC=-0.754)及其下游的固醇12α-羥化酶基因Cyp8b1(t=-4.836,P=1.41×10-4,logFC=-1.106)在Wfs1敲除小鼠中均明顯下調;而旁路途徑中,Cyp27a1在Wfs1敲除小鼠肝臟中無明顯變化(t=-0.319,P=7.54×10-1),但該通路下游的膽固醇25α7羥化酶基因Cyp7b1(t=-5.929,P=1.43×10-5,logFC=-2.330)表現出了明顯的下調[18]。近30年的研究表明,膽汁酸除了參與機體對食物的消化吸收外,其作為信號分子可以激活肝臟和胃腸道中特定的核受體,從而對膽汁酸、葡萄糖、脂肪酸、脂蛋白等的合成、代謝、轉運發揮調控作用[19]。2型糖尿病患者餐后血漿膽汁酸的濃度、組成與健康對照相比均發生明顯變化[20]。膽汁酸可作用于法尼酯衍生物X受體(farnesoid X receptor,FXR)和G蛋白偶聯膽汁酸受體1(G-protein coupled bile acid receptor 1,Gpbar1/TGR5)等多種受體發揮作用[19]。肝臟FXR受體可通過小分子異源二聚體伴侶抑制肝X受體介導的SREBP-1c的轉錄,或通過改變細胞內膽固醇含量間接調控SREBP-1c,從而抑制脂肪生成和脂肪酸合成[21]。膽汁酸在糖代謝中也發揮著重要作用。膽汁酸結合于腸道L細胞上的TGR5受體,從而促進胰高血糖素樣肽-1(glucagon-like peptide 1,GLP-1)的分泌;而結合FXR受體則對葡萄糖刺激產生的GLP-1分泌發揮抑制作用[22]。糖尿病大鼠攝入牛磺膽酸后,血漿胰島素及GLP-1水平升高,空腹血糖水平及葡萄糖耐量均有明顯改善[23]。有研究表明,GLP-1受體激動劑可以增加Wfs1敲除小鼠的胰島素分泌,改善血糖水平,并降低胰島中內質網應激及氧化應激相關標志物的水平[15]。
綜上所述,本研究通過生物信息學的方法,對Wfs1敲除小鼠肝臟組織的芯片數據進行分析,結果提示Wfs1敲除可能影響肝臟膽汁酸合成,從而進一步影響糖脂代謝。針對膽汁酸的相關研究,或許能為WFS1突變患者的治療提供新思路。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
電子制作(2018年18期)2018-11-14 01:48:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
山東工業技術(2016年15期)2016-12-01 05:31:22
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
