砷在海洋食物鏈中的生物放大潛力及發生機制探討
2019-06-10 02:07:30杜森張黎
生態毒理學報 2019年1期
關鍵詞:生物
杜森,張黎
1. 中國科學院南海海洋研究所,中國科學院熱帶海洋生物資源與生態重點實驗室,廣州 510301 2. 中國科學院大學,北京 100049
砷(As)是一種廣泛分布于自然界中的有毒類金屬元素,是危害最嚴重的環境污染物之一。砷位于元素周期表中第四周期第五主族元素,主要以+3和+5的亞砷酸鹽和砷酸鹽的形式存在于環境中。砷主要用于殺蟲劑、除草劑、殺真菌劑和土壤消毒劑,也用來除莠病和消毒種子;工業中也常用來有色金屬的冶煉、半導體元件制造業和凈化工業氣體等[1-2]。在綜合考慮毒性大小和污染發生頻率的基礎上,美國有毒物質與疾病登記署(Agency for Toxic Substances and Disease Registry)和環境保護署(Environmental Protection Agency)將砷排在2017年“優先污染物列表”的首位[3]。不同來源的砷通過陸地徑流、大氣沉降和廢水直接排放等途徑進入海洋生態系統,導致砷污染成為近海區域嚴重的環境問題之一[4]。海洋生物普遍具有較高的砷累積和代謝轉化能力,因而在體內含有高濃度的砷,并主要以有機砷形態存在。這導致海產品成為許多國家和地區人群砷攝入的主要來源(在美國和日本超過90%),一直受到密切關注[5]。
食物相暴露是海洋捕食者(像腹足類和魚類等)累積痕量元素的主要途徑[6],這導致痕量元素沿食物鏈進行傳遞。污染物是否具有生物放大(biomagnification)的能力,是評估其對環境造成危害的重要依據之一。具有生物放大能力的污染物在沿食物鏈/網傳遞的過程中,會隨營養級的增加在生物體內逐級富集,在高營養級生物中達到較高水平,對生物與人類健康產生潛在危害,因此其生態風險和人類健康風險就應當受到格外的關注。有機汞以及持久性有機污染物(如多氯聯苯)等就是典型的具有高生物放大能力的物質,幾十年來已被眾多研究所關注[7-8]。砷通常不被認為是具有生物放大能力的污染物,過去的研究發現,砷在淡水和陸地食物鏈/網中確實常被生物減小(biodiminution)[9-10]。然而,近些年不少研究發現,砷在某些海洋食物鏈/網中出現了生物放大的現象。迄今為止,關于砷沿食物鏈/網傳遞的研究卻非常匱乏,而且對相關機理的認識也不明確。因此,本文總結了近20多年來國內外有關砷在典型海洋生態系統中食物鏈/網傳遞作用的相關研究,初步探討砷在海洋食物鏈/網中是否具有生物放大潛力以及相關砷在不同食物鏈/網中傳遞作用的關鍵核心與影響因素,有助于加深對砷的生物有效性和生物地球化學行為的認識,對準確評估預測砷的生態風險和保障海洋生態安全有十分重要的意義。
1 砷在海洋生態系統中的濃度與形態(Arsenic concentration and speciation in marine ecosystems)
砷具有不同的化學形態(如圖1),不同形態砷的生物毒性具有很大差異。砷在海水和沉積物環境中主要以無機形態存在,而在海洋生物中已鑒定出20多種不同的無機和有機形態,其中無機砷主要包括高毒性的三價砷[As(III)]和五價砷[As(V)],有機砷包括低毒的甲基砷(Monomethylarsonate, MMA)、二甲基砷酸(Dimethylarsinate, DMA)、三甲基砷(Trimethylarsine, TMA),微毒的砷糖(Arsenosugars, AsS)、砷膽堿(Arsenocholine, AsC)和砷甜菜堿(Arsenobetaine, AsB)等[11-13]。與大部分重金屬通過絡合作用形成各種形態不同,這些有機砷形態為共價鍵形成的化合物,具有較為穩定的結構,例如砷甜菜堿即三甲基砷乙酸酯(CH3)3As+CH2COO-,由甜菜堿中氮原子被砷原子取代形成[13]。

圖1 海洋生物體內常見砷化合物的結構Fig. 1 Structures of common arsenic species in marine organism
目前有關海洋生物中砷的研究主要集中在對不同海洋生物中砷的濃度和形態的野外調查[14-16],有少量研究通過室內砷暴露模擬實驗分析海洋生物對環境和食物中砷的累積和轉化機制[17-18]。野外調查顯示,通常海水中砷濃度較低(μg·L-1級),但海洋生物中普遍具有高砷水平,在未受污染的區域處于1~100 μg·g-1范圍內,在一些污染區域可超過1 000 μg·g-1[19]。表1總結了世界范圍內海洋生物整體或肌肉組織中總砷濃度范圍和已檢出的砷化合物形態。由于海洋生物組織中總砷濃度差異很大,這里用幾何平均值表征其典型濃度。
在不同類群海洋生物中,海藻和海草中總砷濃度較低[21]。海藻中的砷約1%~50%為無機形態,更多的是以砷糖和甲基砷為主的有機形態[20,27],這與淡水和陸地植物中以無機砷為主形成鮮明對比。浮游植物與浮游動物處于低等水生食物鏈,后者常以前者為食,所以在浮游動物中也可以檢測到浮游植物中出現的砷形態。浮游動物體內的砷形態也間接反映了其攝食食性,即雜食性種類浮游動物體內累積AsB,而食草性浮游動物體內累積砷糖,并將其轉化為微量AsB[28-29]。
海洋無脊椎動物累積的砷濃度普遍較高,而多毛類和腹足類等底棲無脊椎生物具有很高的砷含量(例如馬氏獨毛蟲Tharyxmarioni中超過2 000 μg·g-1和細角螺Hemifususternatanus中約300 μg·g-1)[21]。多毛類動物沙蠋(Arenicolamarina)體內的砷主要為三價砷(58%)和五價砷(16%),有機砷不到20%;對其進行水相As(V)暴露后發現該多毛類能將As(V)轉化成As(III)和DMA[30]。Chen等[31]對菲律賓蛤仔(Ruditapesphilippinarum)進行30 d無機砷暴露后發現,鰓部累積的砷顯著高于消化腺,蛤體內砷的主要形態為AsB和DMA;亞細胞分布顯示,砷主要分布在金屬硫因蛋白(metallothionein-like proteins, MTLPs)和富金屬礦體(metal-rich granules, MRG)中。MTLPs能夠與金屬結合從而在細胞內金屬的轉運和隔離中發揮重要作用[32],并增加生物對砷的耐受能力。
海洋魚類中的砷濃度范圍也非常大,與無脊椎動物類似,底棲魚類比浮游魚類含有更高的砷濃度(例如孔鰕虎魚Trypauchenvagina中高達134 μg·g-1[22])。砷甜菜堿是絕大多數海洋動物組織中最主要的砷形態,通常占海洋甲殼類和軟體動物組織中總砷的50%~95%,在海洋魚類組織中占80~99%以上。相反,砷在淡水生物中主要以無機砷為主,雖然也有研究報道砷甜菜堿的存在,但它占總砷的比例遠低于海洋生物[24]。
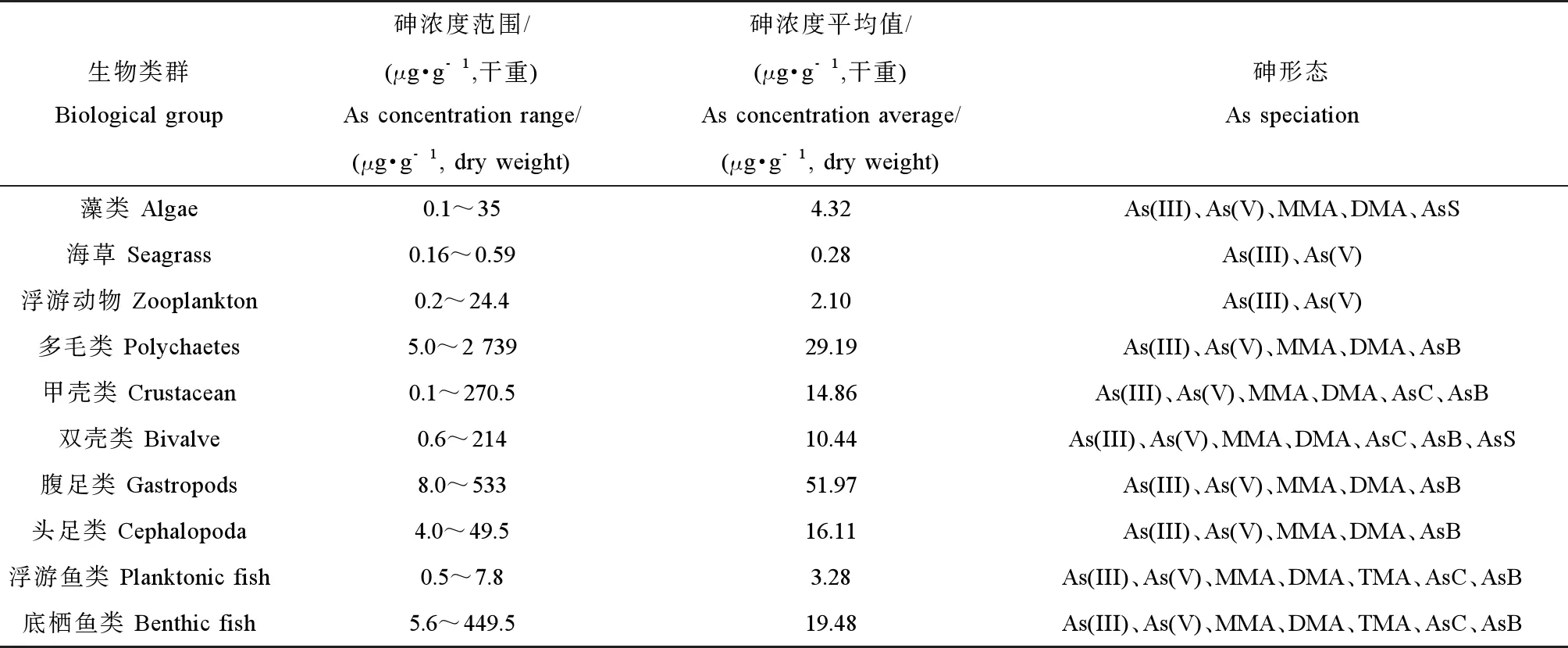
表1 海洋生物體內總砷的濃度和所檢出的砷化合物形態[20-26]Table 1 The total arsenic concentration and arsenic speciation of marine organisms[20-26]
海洋魚類累積砷的生物動力學模型可以描述為[33-34]:
dC/dt=ku×Cw+ AE×IR×Cf- (ke+g)×C
式中,dC/dt為累積速率,ku為水相吸收常數;Cw為水中砷濃度,AE(assimilation efficiency)為同化率;IR為攝食率,Cf為食物中砷濃度,ke為排出速率常數,g為生長速率,C為生物體內砷濃度。砷可以通過水相和食物相進入海洋生物體內,ku、AE和ke是影響海洋生物體內砷累積的3個重要參數,表2列舉了通過室內模擬實驗得出的常見海洋生物中的生物動力學參數。海洋魚類對水相砷的吸收速率遠小于小型底棲生物,尤其是在雙殼類中,砷的水相吸收速率和同化率較高,而排出速率相對較低,說明其對砷的累積能力較強,這也與調查中發現的小型底棲生物高砷含量一致。
雖然目前有關海洋生物砷含量和形態的數據很多,但是由于這些研究通常僅針對一種或數種生物開展,而這些生物又受到各自所處不同環境條件的影響而具有很大的砷濃度范圍,因此很難根據這些數據分析不同營養級生物對于砷累積的差異性,只有針對特定生態系統中較完整的食物鏈進行調查研究,才能反映砷沿食物鏈傳遞的特征和規律。
2 砷在海洋食物鏈/網中的生物放大潛力(Arsenic biomagnification potential along food chains/web)
生物放大(biomagnification)一般是指化學物質在生物攝食作用下,導致了生物體內化學物質的濃度高于食物中的濃度的現象。也有一些學者將生物放大現象定義為濃度隨營養級的增加而升高,當生物放大系數(biomagnification factor, BMF)大于1時,這個物質就被生物放大。前者將濃度的升高限制在食物相暴露的因素下,而后者的影響因素包括通過水相暴露和食物相暴露,其中生物從水相吸收污染物的過程稱為生物濃縮(bioconcentration)[43]。由于海洋動物中的砷累積主要由食物相獲得[35-39],所以對于海洋食物鏈而言,這2種定義下的砷生物放大作用差異不大。
表3總結了砷在不同類型海洋生態系統中沿食物鏈傳遞的數據。這些研究較為綜合地分析了某一特定生態系統中環境(海水和沉積物)中砷的背景濃度和不同類群生物中的砷含量,同時部分研究通過氮穩定同位素(δ15N)確定了這些生物的營養級,可較有效反映砷在自然狀況下的食物鏈傳遞行為。從表中可以發現,不同類型的生態系統中砷的食物鏈/網的傳遞存在明顯的差異。總體上說,砷在水生生態系統中的食物鏈傳遞可以歸納為圖2中的4種模式,即生物減小、生物放大、無顯著趨勢和局部環節生物放大。
在海洋生態系統中,僅發現砷在意大利南部沿岸區域有顯著生物減小的現象[44],研究人員對7種大型海藻、2種海草、5種無脊椎動物、9種魚類和2種鳥類(羽毛)進行采樣分析后發現,砷在沉積物、大型藻類和海草中的濃度最高(分別高達11.8、35.4和17.4 μg·g-1),然后隨著營養水平的增加而減少,在三級消費者和頂級捕食者(魚類和鳥類)中的As濃度低于2 μg·g-1,這些不同生物中As濃度的對數值(Log10As)與δ15N之間呈現出顯著的負相關性,清晰地表明砷在該食物網中的生物減小作用。需要指出的是,這一研究區域嚴格上說并不是一個單一的食物鏈/網,而是一個開放區域的生物集合,因此其結果可能無法反映真實的砷食物鏈傳遞結果。相反,砷在其他的海洋生態系統中的食物鏈傳遞主要呈現出生物放大(圖2B)、無顯著趨勢(圖2C)和局部環節生物放大(圖2D)等多種現象。在日本駿河灣生態系統中發現生物(浮游動物、頭足類和魚類)體內砷含量(Log10As)與營養級(δ15N值)呈顯著正相關(P<0.05),有著明確的生物放大作用[45]。砷在中國大鹿島底棲食物鏈中也存在一定的生物放大趨勢[46]。蘇祿海底層魚類體內砷含量(Log10As)與營養級(δ15N值)之間同樣存在顯著的正相關(P<0.01),且BMF達1.70[47]。在珊瑚礁生態系統中,砷則出現了生物放大的趨勢[48]。在南極和北極的極地海洋性生態系統中,砷的濃度與營養級之間無顯著相關性[49-50]。
注:ku為水相吸收常數;AE為同化率;ke為排出速率常數。
Note:kustands for waterborne uptake rate constant; AE stands for assimilation efficiency;kestands for efflux rate constant.
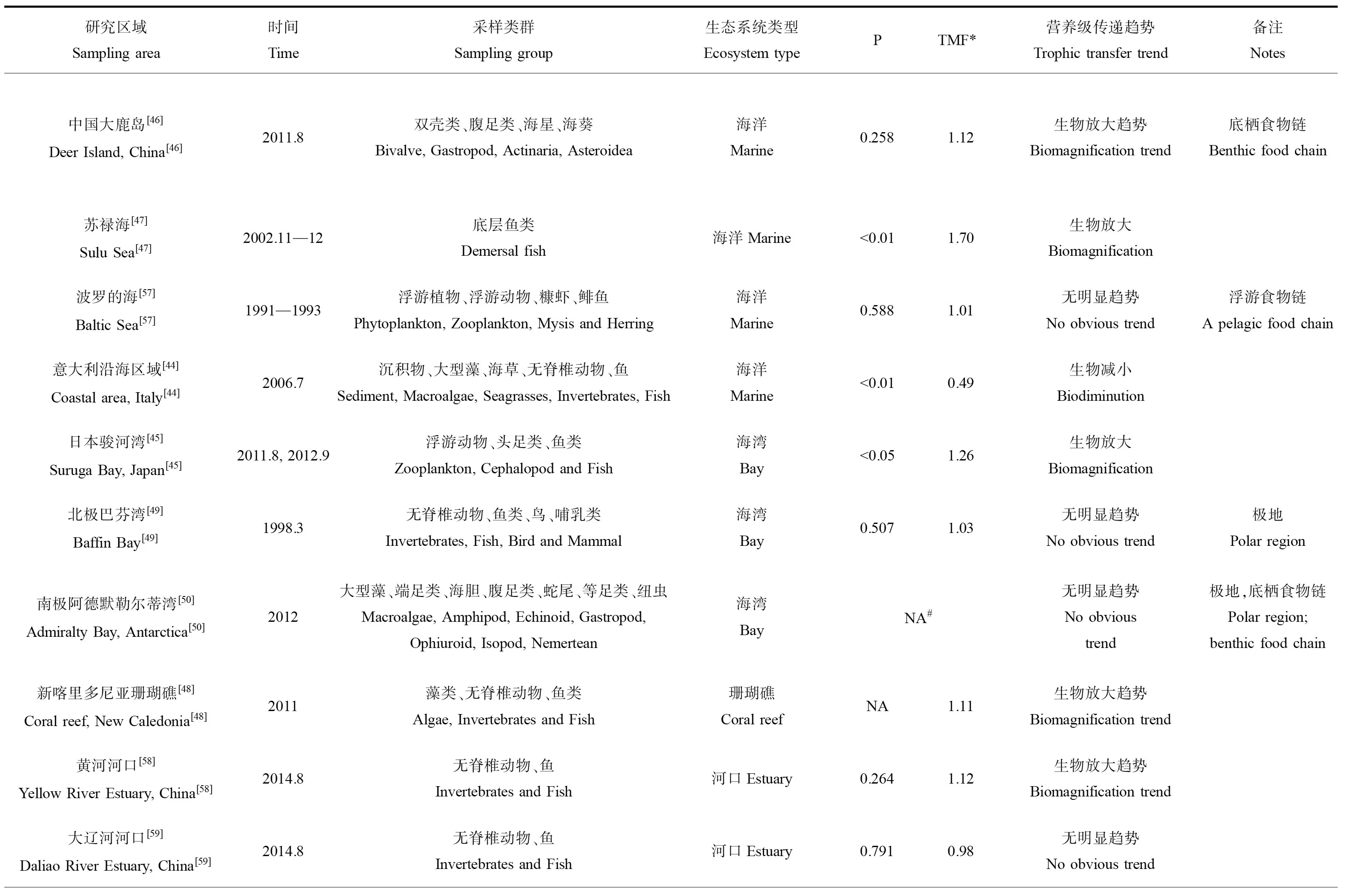
表3 砷沿不同水生生態系統食物鏈/網傳遞研究匯總Table 3 Summary of arsenic trophic transfer in different aquatic ecosystems
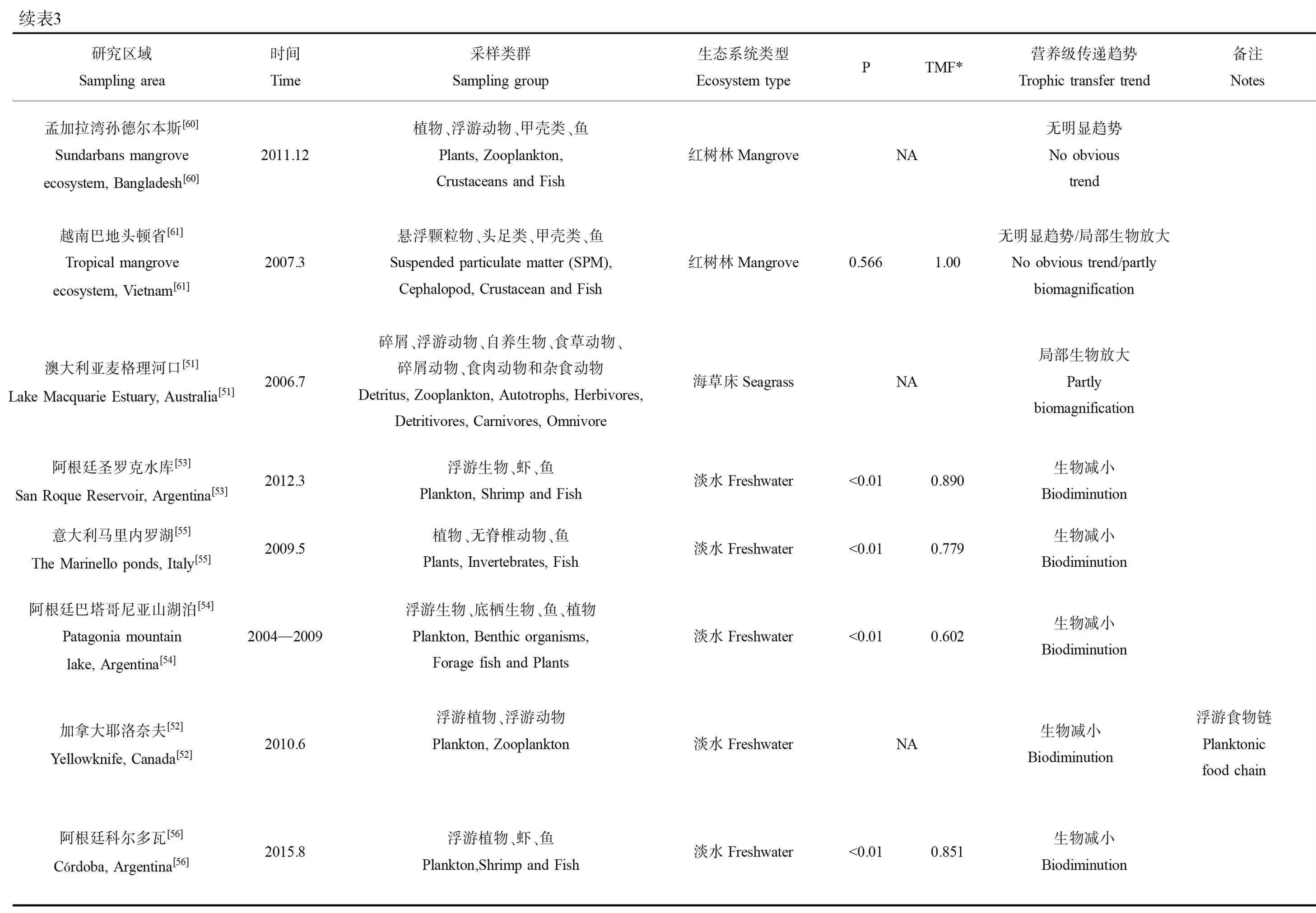
續表3研究區域Sampling area時間Time采樣類群Sampling group生態系統類型Ecosystem typePTMF*營養級傳遞趨勢Trophic transfer trend備注Notes意大利馬里內羅湖[55]The Marinello ponds, Italy[55]2009.5植物、無脊椎動物、魚Plants, Invertebrates, Fish淡水Freshwater<0.010.779生物減小Biodiminution阿根廷巴塔哥尼亞山湖泊[54]Patagonia mountain lake, Argentina[54]2004—2009浮游生物、底棲生物、魚、植物Plankton, Benthic organisms, Forage fish and Plants淡水Freshwater<0.010.602生物減小Biodiminution加拿大耶洛奈夫[52]Yellowknife, Canada[52]2010.6浮游植物、浮游動物Plankton, Zooplankton淡水FreshwaterNA生物減小Biodiminution浮游食物鏈Planktonic food chain阿根廷科爾多瓦[56]Córdoba, Argentina[56]2015.8浮游植物、蝦、魚Plankton,Shrimp and Fish淡水Freshwater<0.010.851生物減小Biodiminution
注:#NA表示無有效數據;TMF表示營養級放大系數。
Notes:#NA means no valid data; TMF stands for trophic magnification factor.
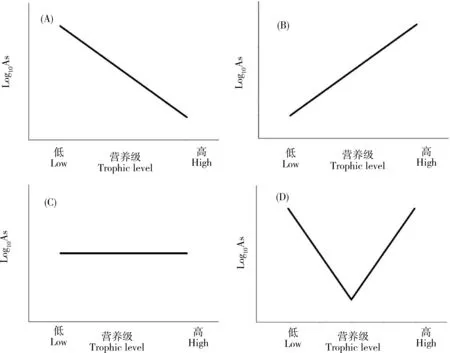
圖2 砷沿食物鏈/網傳遞模式圖注:(A)生物減小、(B)生物放大、(C)無顯著趨勢、(D)局部環節生物放大。Fig. 2 The mode pattern of arsenic transfer along food chains/websNote: (A), biodiminution; (B), biomagnification; (C), no significant trend; (D), biomagnification in part of food chains.
在沿海的淺海生態系統(包括河口、紅樹林和鹽沼海草床等),砷同樣表現出了生物放大的潛能。Barwick等[51]對澳大利亞溫帶河口海草生態系統中食物網的研究發現,在碎屑、浮游動物、自養生物、食草動物、碎屑動物、食肉動物和雜食動物間的35條經驗食物鏈中有19條出現砷濃度從被捕食者到捕食者增加的現象;在浮游動物(2.1 μg·g-1)→浮游雙殼類動物(4.8 μg·g-1±0.9 μg·g-1)→肉食性魚(6.2 μg·g-1±0.4 μg·g-1)和碎屑(6 μg·g-1±3 μg·g-1)→雜食性蝦(8 μg·g-1±1 μg·g-1)→雜食性魚(11 μg·g-1±2 μg·g-1)2條食物鏈中觀察到As的生物放大,而這2條食物鏈都屬于底棲食物鏈。
砷在淡水生態系統中的食物鏈傳遞研究相對較少,但是都一致地表現出顯著的生物減小作用(圖2A),僅在BMF值上有所差異[52-56]。例如,在阿根廷的一個湖泊調查中發現,砷在由浮游植物、底棲生物、魚和植物組成的食物網中出現顯著的生物減小現象(P<0.01)[54]。
從這些研究結果中不難發現,砷沿海洋食物鏈/網傳遞具有一定的復雜性,但是普遍具有隨營養級升高維持高生物累積或生物放大的潛力,與淡水食物鏈/網中砷的生物減小形成顯著的對比。然而,這些研究均存在一定的局限性(例如采樣沒有覆蓋主要生物類群,沒有用δ15N確定生物營養級,缺少對同一生態系統的重復性調查等),造成研究結果的諸多不確定性,使得砷在海洋食物鏈/網中是否生物放大仍然存在疑問。更重要的是這些研究基本都沒有對不同砷形態進行分析,而砷形態的區別恰恰是海洋生物和淡水生物最大的差異,很可能是解析砷在海洋生態系統中生物放大潛力的關鍵因素。
3 砷在海洋食物鏈/網中生物放大潛力的可能機制(Possible mechanisms of arsenic biomagnification potential along food chains/web)
目前已有研究從生物動力學角度來分析無機砷的食物鏈傳遞,在幾種海洋魚類和無脊椎動物中測定了控制食物鏈傳遞的2個參數(同化率和排出率),通過模型計算表明,無機砷難以在食物鏈中被生物放大[18,35,37]。Zhang等[62]研究了蛤和多毛類對沉積物中砷的生物動力學和生物可利用性,發現生物對砷的累積與沉積物中砷的形態有關,砷甜菜堿是有機砷中同化效率最高的形態,而As(III)的同化效率較低且排出速率較快。海洋魚類對痕量元素的同化率受被捕食者亞細胞痕量元素的分布以及魚類的攝食過程所影響[63],而不同的食物對海洋魚類As的生物有效性和生物累積存在顯著的影響作用[64]。痕量元素能夠在底棲食物鏈中生物放大的主要原因可能是腹足類能夠有效吸收食物中的痕量元素,而浮游生物食物網生物減小的發生則可能是其對痕量元素較低的同化率和較高的排出率[65]。
因此,砷的生物放大作用很可能是由有機砷的食物鏈傳遞作用所引起的。痕量元素的有機形態通常比無機形態具有更高的生物可利用性和食物鏈傳遞能力,其中了解最多的為汞。有機汞(以甲基汞為主)具有很強的親脂性,可以直接通過細胞膜結構進入細胞并與生物分子結合,因此具有比無機汞更高的生物可利用性。而甲基汞的生物放大作用也遠超無機汞,是痕量元素生物放大的最典型案例[7]。由此,有機砷可能比無機砷更容易被生物放大,這在一定程度上解釋了砷在海洋和淡水生態系統中食物鏈傳遞的差異現象。在越南南部沿海的紅樹林生態系統中通過對懸浮顆粒物、頭足類、甲殼類和魚類的調查后發現,盡管總砷濃度(Log10As)與營養級(δ15N值)之間沒有顯著的相關性(P=0.556),但生物體內的砷甜菜堿出現了明顯的生物放大現象(TMF=1.30,P<0.01)[61]。
海洋魚類和貝類普遍具有將從環境中吸收的無機砷通過生物轉化合成砷甜菜堿的能力(如圖3),這種生物轉化過程包括As(V)還原為As(III),As(III)甲基化至一甲基砷和二甲基砷,以及經過后續未知的過程合成砷甜菜堿[66-68]。砷甜菜堿的合成,有助于海洋魚類將砷累積在體內,從而達到較高富集的結果。在這種生理作用下,當魚類長期暴露在高無機砷環境中時,導致的結果是較高砷甜菜堿的富集,這也是魚類的一項重要解毒機制[69]。在溫帶海洋鹽沼系統中,植物體內砷主要為無機砷,但攝食這些植物的動物體內砷的主要形態為砷甜菜堿和砷糖為主的有機砷,說明動物將吸收的無機砷生物轉化成了有機砷[70]。Zhang等[37]通過構建了2條底棲食物鏈(沉積物—沙蠶Nereissuccinea—諸氏鯔鰕虎魚和沉積物—蛤Gafrariumtumidum—鰕虎魚)研究了砷沿底棲食物鏈的傳遞和生物轉化作用,發現蛤對砷的吸收效率分別為35%~65%,沙蠶對砷的吸收效率分別為52%~73%,從而造成沙蠶比蛤具備更高的砷累積能力;同時,在沉積物中砷的形態以無機砷為主,到初級消費者體內砷甜菜堿成為主要形態,再到魚類體內砷甜菜堿可以占到95%以上,說明無機砷在底棲食物鏈中被高效轉化,并主要發生在初級消費者環節,該結果也印證了As在底棲食物鏈中存在更大生物放大潛能。
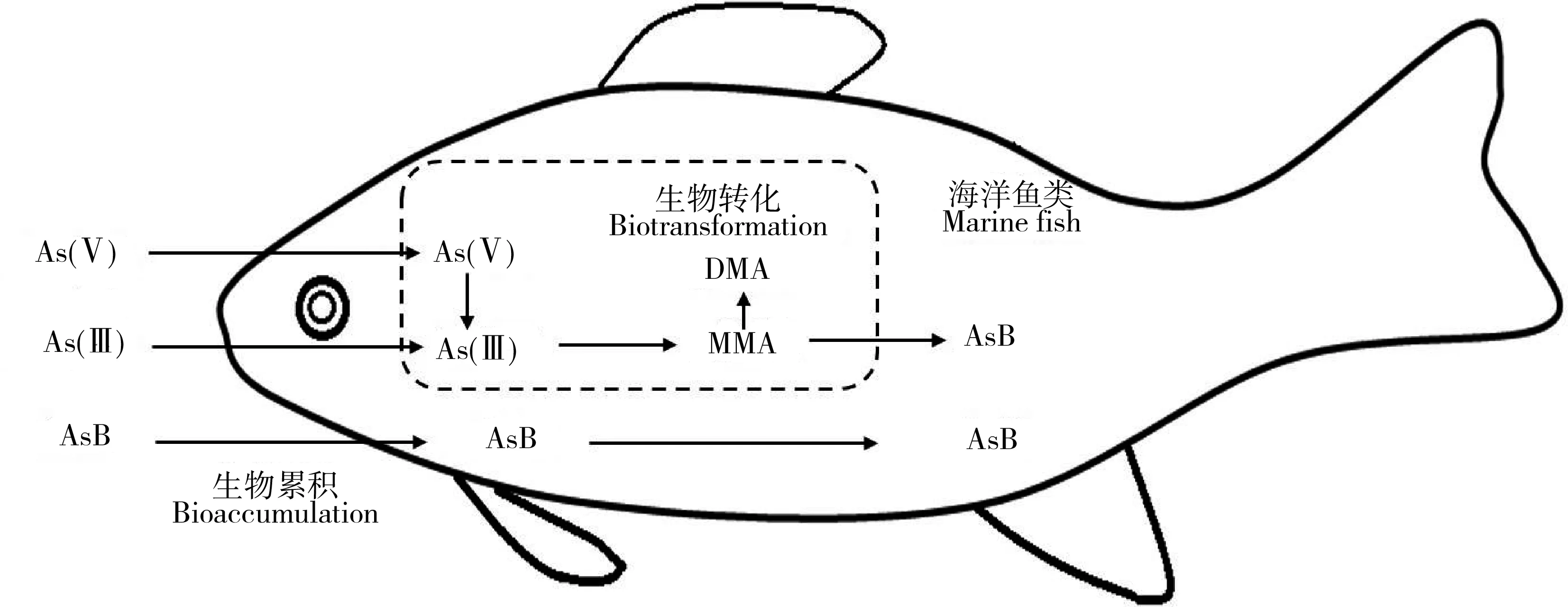
圖3 海洋魚類對砷的生物轉化[67]Fig. 3 Arsenic biotransformation of marine fish[67]
當砷從不同的餌料生物傳遞到海洋魚類時,餌料中的有機砷比例與總砷的營養級放大系數(TMF)呈現顯著的正相關,表明海洋生物中有機砷比無機砷具有更高的食物鏈傳遞能力,可以導致海洋魚類富集更高濃度的砷[64]。Zhang等[67]研究了海洋魚類(草食性籃子魚Siganusfuscescens和肉食性鱸魚Lateolabraxjaponicus)對砷的營養轉化和生物可利用性,發現食物中的無機砷較難被海洋魚類吸收,而是在體內被生物轉化成了有機砷,而食物中的砷甜菜堿可直接通過消化器官的上皮細胞被魚類吸收,成為魚體內砷的最終存儲形態。對澳大利亞紅樹林的調查顯示,紅樹林生態系統中的動物主要從食物中吸收有機砷(砷甜菜堿和砷核糖等)[71]。長期砷甜菜堿的暴露會導致大西洋鮭魚(Salmosalar)和大西洋鱈魚(Gadusmorhua)肌肉中砷濃度顯著增加[72]。目前關于有機砷食物鏈傳遞的生物動力學研究較少,因此需要對有機砷(特別是砷甜菜堿)的生物累積動力學參數進行量化解析,厘清其對總砷食物鏈/網傳遞的貢獻。
除了生物體中的砷形態(內因)可能直接作用于砷的食物鏈傳遞以外,海洋生態系統中的生物和環境因素(外因)也可能對其造成影響。環境因素如溫度、濁度、營養鹽和光照等可影響生物中的砷形態,間接作用于砷的食物鏈傳遞;也可能通過影響生物的生長速率和代謝率(特別是小個體生物),間接影響砷的攝入、代謝和排泄[73-74],這些作用可綜合體現在砷生物累積的季節變化上。鹽度能夠影響大亞灣幾種常見魚類和甲殼類體內砷的累積,尤其是砷甜菜堿的濃度;同時沉積物中高砷的濃度對海洋魚類體內砷的累積也有重要影響[75]。室內實驗也證明高鹽度會導致貽貝(Mytilusedulis)對砷甜菜堿的高累積[76]。有研究認為海藻通過細胞磷酸鹽轉運系統累積海水中的砷酸鹽,但是其體內的砷糖水平更主要是受到海水中氮鹽的影響[77]。食物網中生物組成的不同與所處環境的差異也會對砷的食物鏈傳遞產生影響。底棲無脊椎動物和魚類往往比游泳生物含有更高濃度的砷,除了物種差異的原因外,還包括不同生物類群的食性不同,食物中砷濃度和形態的差異也會對砷的累積產生差異,因此,當食物網中生物組成偏向于底棲食性時,可能會導致更大的生物放大潛力[37,62]。因此,當研究砷的食物鏈傳遞時,不能僅著眼于砷本身而忽視了其他環境因素的作用。
4 總結與展望(Summary and prospect)
海洋生物普遍具有高含量的砷,這些砷主要為低毒性的有機砷形態。砷在淡水生態系統中普遍生物減小,但是在海洋生態系統中出現了一定的生物放大潛力。海洋魚類和貝類普遍具有將從環境中吸收的無機砷通過生物轉化合成砷甜菜堿的能力,有機砷比無機砷具有更高的食物鏈傳遞能力,可以導致海洋魚類富集更高濃度的砷。因此,有機砷的存在可能有助于砷沿著海洋食物鏈/網富集,在某些情況下被生物放大。但是目前關于砷在海洋生態系統中生物放大潛力及機制的研究較為匱乏,今后應該從一下幾個方向開展:
(1)不同砷形態如何在海洋食物鏈/網中傳遞的研究。目前關于砷在生態系統中的傳遞主要是關于總砷的研究,缺乏對不同砷形態在傳遞的過程中累積、傳遞和轉化的研究工作。
(2)砷在海洋生態系統中生物放大潛力的影響因素。過去主要關注痕量元素在生態系統中傳遞的調查工作,應該系統地研究砷在不同海洋食物鏈/網中從初級生產者到高營養級生物中的行為,通過比較不同物種對砷累積的差異以及不同食物鏈之間的異同,厘清引起砷生物放大的關鍵因素。
(3)砷甜菜堿合成關鍵基因及其調控機制的研究。海洋魚類能累積較高含量的砷,其中的砷主要為砷甜菜堿,所以應該采用分子生物學等方法研究砷甜菜堿合成的關鍵基因和調控機制,闡明海洋生物高砷累積的調控機制。
(4)室內模擬實驗與野外調查相結合,通過不同砷形態食物鏈傳遞的室內實驗來驗證砷形態是否是導致海洋生態系統中砷放大的關鍵原因,同時驗證相應的環境影響因素,最終闡明不同砷形態的食物鏈傳遞作用及其對砷生物放大的影響,對于準確評估砷的生態風險,進一步闡明砷元素的生物地球化學過程有著重要作用。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
