基于RAPD標記對71個柞蠶品種的聚類分析
2019-07-22 01:32:45劉丹梅李文利王丹丹
江蘇農業科學 2019年11期
劉丹梅 李文利 王丹丹


摘要:采用隨機引物擴增多態性DNA(RAPD)的方法,在DNA分子水平上對我國不同地區、不同化性的71個柞蠶品種進行聚類分析。從140條RAPD隨機引物中篩選出28條可穩定擴增條帶的多態性引物,共擴增出92條多態性條帶,占總擴增帶數的29%;采用非加權組平均法(unweighted pair-group method with arithmetic means,簡稱UPGMA)聚類法對試驗結果進行分析,構建了71個柞蠶品種的親緣關系,結果顯示,各個地區與不同化性的柞蠶品種之間親緣關系沒有明顯區別,遺傳類型相互混雜。
關鍵詞:RAPD標記;柞蠶;UPGMA;聚類分析;親緣關系
中圖分類號: S885.1? 文獻標志碼: A? 文章編號:1002-1302(2019)11-0061-05
柞蠶屬于鱗翅目(Lepidoptera)天蠶蛾科(Saturniidae)的絹絲類昆蟲,我國境內的柞蠶在分類學上屬于柞蠶(Antheraea pernyi),主要分布在10余個省份。目前保存著上百個地理種群或生態類型,如山東的客嶺莊種、河南的魯山種、遼寧的璦陽種、貴州的湄潭種以及用顏色命名的青皮蠶、黃皮蠶、青黃蠶、銀白蠶、藍靛蠶等[1]。這些品種都是經過長期的人工選擇培育以及自然突變逐漸形成的具有不同生理、遺傳特性和區域適應性的柞蠶品種,其經濟性狀各具特征[2]。
隨著分子遺傳學的發展,可以通過DNA大分子本身來分析遺傳差異,為物種的起源、親緣關系和進化研究開辟了一個新的途徑。這些研究所提供的大量客觀的分子水平上的信息,非常有力地推動了物種起源、親緣關系和進化的研究。20世紀90年代發明的分子標記技術,特別是隨機擴增多態性DNA(random amplified polymorphic DNA,簡稱RAPD)、選擇性擴增DNA片段(selective amplification DNA fragments,簡稱SADF)等技術[3-7],以其快速方便的特點,在核酸分子水平上為物種的多態性提供了豐富的信息,從而被廣泛應用于物種遺傳變異、進化和親緣關系等研究領域。這些技術的應用已經揭示了許多物種種內或種間的遺傳進化與親緣關系,如亞洲玉米螟地理種群分化的研究[8]、蚱屬種間親緣關系研究[9]、我國野桑蠶和家蠶的系統發育關系研究等[10]。目前已利用這些標記技術成功地構建了家蠶的連鎖圖[11-14]。在柞蠶上,此方面的研究也已經開展起來[15],劉彥群等利用RAPD標記分析4種體色柞蠶品種的遺傳關系,表明品種的遺傳聚類劃分與體色之間的相關性不大,柞蠶體色并不能真實反映品種間的遺傳關系[16];靳向東等作了廣適性大型多絲量柞蠶新品種選育報告,將柞蠶品種資源研究運用于新品種選育中,育成2個多絲量柞蠶新品種[17]。在技術上,RAPD技術已被應用在多種動植物上,如農志歡等在十大功勞屬6個種上[18]、李言等在海帶上[19]、石洪玥等在中華絨螯蟹上[20]、陳德西等在蝗蟲上[21]都利用RAPD技術開展了種質資源的研究。目前,國內的研究主要集中在家蠶、玉米、水稻等生物的種質資源方面,有關柞蠶種質資源遺傳進化、分類以及親緣關系的研究在深度和廣度上有待完善。因此,本研究采用RAPD技術對71個柞蠶品種的分類和親緣關系進行了聚類分析,以確立品種間的親緣關系,以期對我國這一特有生物資源開展進一步的分子生物學研究。
1 材料與方法
1.1 試驗材料
收集我國遼寧、河南、吉林、山東與內蒙古等地的不同地理種群和不同化性的柞蠶品種(表1)[22-26]。本研究在大連理工大學生命科學實驗室完成。
參照美國Operon公司設計的含有10個寡核苷酸隨機引物,由寶生物工程(大連)有限公司合成。RAPD反應系統購自北京鼎國生物工程有限公司。Protease K(蛋白酶K)、RNaseA購自美國Promega公司,瓊脂糖為西班牙產,其他常用試劑均為國產分析純。
1.2 方法
1.2.1 柞蠶基因組DNA的提取 取液氮中冷凍的單個柞蠶蛹組織0.5 g研磨[27],加入2 mL提取緩沖液[10 mmol/L 三(羥甲基)氨基甲烷(Tris)-HCl pH值8.0,0.1 mmol/L 乙二胺四乙酸(EDTA),0.5% 十二烷基硫酸鈉(SDS),50 μg/mL 蛋白酶K],在50~60 ℃保溫3 h[28-31]。加入等體積的酚混勻,4 ℃、12 000 r/min離心10 min;取上層液加入等體積的酚/三氯甲烷抽提1次。最后用2/3體積的異丙醇沉淀,用75%乙醇洗沉淀1次。溶于400 μL TE中,加RNase于37 ℃消化30 min,-20 ℃保存備用。
1.2.2 RAPD反應體系及擴增程序 參照Williams等的方法[3]對隨機擴增具有影響的反應參數, 如鎂離子、引物、模板濃度、dNTP和Taq酶用量等進行優化[32-34]。25 μL反應液中,含2 U Taq酶、2.5 μL 10×緩沖液(1.5 mmol/L MgCl2+10 mmol/L Tris-HCl+50 mmol/L KCl+0.01%明膠)、dNTP(各200 umol/L)和Primer(10 pmol/L)各1.0 μL、DNA模板1.0 μL(100 ng)。反應程序如下:94 ℃預變性3 min;94 ℃變性30 s,37 ℃退火40 s,72 ℃延伸2 min,35個循環; 72 ℃延伸10 min。
1.2.3 RAPD多態性分析 每個柞蠶品種取6個個體,分別提取DNA后混合作為模板進行PCR反應。將RAPD擴增產物在1.4%瓊脂糖凝膠中恒壓電泳2 h,經溴化乙錠(EB)染色后在紫外分析儀下觀察拍照,選取重復性好且亮度較強的條帶進行統計分析[35]。在電泳圖譜上同一RAPD位點上有電泳帶者記為1,無電泳帶者記為0。根據Nei的公式:I=2NXY/(NX+NY)[36],計算出任意2個個體(群體或物種)間的遺傳距離D和遺傳相似系數I。式中:NXY為兩者共同擁有譜帶(RAPD標記)的數目;NX和NY分別為X和Y個體(或群體、物種)各自擁有的全部譜帶數。遺傳距離的計算和系統聚類采用NTSYS-PC 2.10e[37]。
2 結果與分析
2.1 引物篩選及PCR擴增
采用已經確定的反應體系和條件,以小黃皮、魯黃、寬青和抗病2號的DNA作為模板,對7組共140條隨機引物進行隨機擴增,結果共篩選出有效引物60條,再從中篩選出較好的引物28條(表2)。對71個柞蠶品種的DNA進行擴增,共得到92條清晰的具有多態性的譜帶,占總帶數的29%,片段大小介于500~3 000 bp間,平均每條引物可擴增3條具有多態性的譜帶。多態性譜帶最多的引物為OPM10(5條),最少的為OPG3(2條)。可見不同引物的擴增產物均有顯著差異,其中引物OPH19的擴增結果見圖1。此外,不同品種擴增的具有多態性的帶數亦不相同,其中多態性譜帶最多的品種為印度柞蠶,共有35條,最少的為青6號,只獲得16條。
2.2 柞蠶品種間的親緣關系分析
將每個DNA片段看作1個可區分的RAPD標記,根據Nei的公式[36]計算出71份柞蠶品種間的遺傳相似系數(I)和遺傳距離(D)。結果顯示:2個河南一化性品種731與云白的相似系數最高,達到0.886 7,可見兩者間的遺傳背景比較接近;二化品種黃安東與一化品種豫5號的相似系數為 0.867 1,表明兩者之間也有較近的親緣關系。一化性品種河41同白一化、豫早1號同201、豫早2號同魯松、魯黃同松黃等4對品種的相似系數都在0.785 7以上,表明這些品種間的親緣關系較近。而河41、白一化與其他河南一化性品種的相似系數均低于0.486 2,表明這2個品種與其他一化性品種的親緣關系較遠。
用非加權組平均法(UPGMA)對不同化性和不同地理區域的71個柞蠶品種根據遺傳距離(D)進行聚類分析。由圖2可以看出,大部分的河南一化品種分布在不同的類群中,并且在各個類群中首先聚到一起。但是,也有不同地區與化性的品種能夠較早地聚在一起,如黃安東與豫5號,它們僅次于河南一化性品種731與云白,第2次就聚在一起。圖2還說明,各個地區與不同化性的柞蠶品種之間親緣關系沒有明顯區別,遺傳類型相互混雜。
2.3 不同地區柞蠶種群的DNA多態性聚類分析
為了進一步揭示不同地理區域間柞蠶品種的遺傳進化關系,根據每個柞蠶品種的最初來源,選擇具有代表性的、背景清楚的品種作為該地區的代表性品種(表3)。
將表2中來自同一個地區的所有代表性品種的標記作為該地區種群的RAPD標記,進行不同地區柞蠶種群的遺傳距
離(D)分析(表4)。
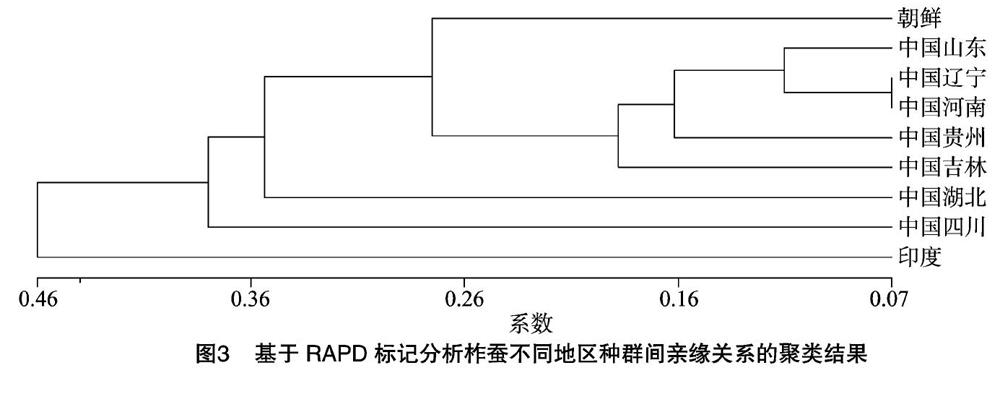
利用表3數據,用UPGMA法構建聚類樹狀圖。從圖3可以看出,來自河南地區的種群與遼寧地區的種群首先聚類到一起, 兩者之間的遺傳距離只有0.066 3。山東與遼寧、山東與河南種群的遺傳距離分別為0.104 2、0.126 1。除了四川與湖北、四川與印度的遺傳距離分別為0.693 1、0.665 0(>0.5)外,其他地區之間的遺傳距離都<0.5。
3 結論與討論
柞蠶為鱗翅目(Lepidoptera)天蠶蛾科(Saturniidae)的絹絲類昆蟲,已記載的共有35個種,其中31個種分布在印沃區,3個在古北區,1個在北美區。為人們所熟知并用于商業性生產的有中國柞蠶(Antheraea pernyi)、2種印度柞蠶(Antheraea assamensis和Antheraea mylitta)和日本天蠶(Antheraea yamamai)。
我國現存130多個品種都是中國柞蠶的變種和地理種或生態型。各個品種之間的血緣關系非常復雜,各個品種之間既有高度變異的一面,又有親緣很近的一面。劉彥群等通過對河41、四青、青黃1號和杏黃4個品種的研究發現,在DNA水平上的分類并沒有按體色進行[38],對現行的體色分類系統提出了質疑。
本研究表明,不同的品種,即使分布相距很遠,有的竟表現出驚人的相似性和緊密的親緣關系;而同一個品種的不同生態型,有的卻相差懸殊。如在對71個柞蠶品種進行RAPD多態性聚類分析時發現,整個系統聚類圖分為2支,每個分支中各地區的品種相互摻雜。但是在第一集團的40個品種中,一化性品種有15個,占37.5%;在第二集團31個品種中,一化性品種只有8個,占25.8%。從整個系統聚類圖上可以看出,大部分的一化品種分布在不同的類群中,并且在各個類群中首先分別聚到一起。河南的一化性品種731與云白的相似系數最高,達到了0.886 7,可見兩者間的遺傳背景非常接近;一化性品種河41同白一化、豫早1號同201、豫早2號同魯
松、魯黃同松黃4對品種的相似系數也都在0.785 7以上,表明這些品種間的親緣關系較近。但是,也有不同地區與化性的品種能夠較早地聚在一起,如黃安東與豫5號,它們僅次于2個河南一化性品種731與云白,相似系數為0.867 1。
有關柞蠶的分布與分化,印度學者喬利認為,生長在熱帶的印度柞蠶(A.assamensis)是柞蠶的原始類型,其他品種都起源于它,我國境內的柞蠶是由印度東北向我國南部傳播的[39]。本研究結果表明,遼寧地區的品種首先與河南地區的品種聚在一起,然后又與山東地區的品種聚在一起。以0.728 1為分界,除了貴州以外,包括朝鮮在內的長江以北的地區品種首先聚類在一起,這一集團中包括我國2個古代柞蠶傳播中心——山東和河南與現代柞蠶的傳播中心遼寧。印度柞蠶最早從整個系統中分離出來,然后是四川、湖北。雖然在古文獻中沒有找到有關這些地區柞蠶品種起源的記載,但是從系統聚類圖中可以推測,在遠古的時候柞蠶由印度傳到我國后,由四川經湖北向長江以北地區傳播,到達我國古代柞蠶傳播中心山東、河南與現代柞蠶的傳播中心遼寧。然后又從這些地區向吉林、貴州以及朝鮮等地傳播,所以發現吉林、貴州以及朝鮮的品種又從這些品種中分化出來。在最后剩下的遼寧、河南、山東的品種中,遼寧與河南分化得最晚;長江以北的地區品種的分化情況基本上與歷史資料的記載相吻合[1]。
現代遺傳學的觀點認為物種的遺傳性狀是由基因決定的,各種外部形態及內部性狀的變異都是由于基因中DNA分子堿基序列發生改變。本研究在分子水平上對我國柞蠶品種資源的遺傳分化和親緣關系進行了探討,可能與已有的研究不完全相符,期望有助于對我國這一特有生物資源的保護和深入研究。
參考文獻:
[1]遼寧省蠶業科學研究所. 中國柞蠶品種志[M]. 沈陽:遼寧科學技術出版社,1994.
[2]郭奕生,陸 俊. 微衛星標記在雜交稻種子純度鑒定上的應用[J]. 廣東農業科學,2014,41(16):138-141.
[3]Williams J G,Kubelik A R,Livak K J,et al. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers[J]. Nucleic Acids Research,1990,18(22):6531-6535.
[4]Welsh J,McClelland M. Fingerprinting genomes using arbitraryprimers[J]. Nucleic Acids Research,1990,18(24):7213-7218.
[5]van der Voort J R,Wolters P,Folkertsma R,et al.Mapping of the cyst nematode resistance locus Gpa2 in potato using a strategy based on co migrating AFLP markers[J]. Theoretical and Applied Genetics,1997,95(5/6):874-880.
[6]Keim P,Schupp J M,Travis S E,et al. A high-density soybean genetic map based on AFLP markers[J]. Crop Science,1997,37(2):537-543.
[7]陳 洪,王振山,朱立煌. SRFA法構建水稻DNA指紋圖譜[J]. 生物工程學報,1996,12(3):266-269.
[8]孫 姍,徐茂磊,王戎疆,等. RAPD方法用于亞洲玉米螟地理種群分化的研究[J]. 昆蟲學報,2000,43(1):103-106.
[9]蔣國芳,陸 敢,黃 琨,等. 用RAPD標記研究蚱屬五個種間的親緣關系[J]. 昆蟲學報,2002,45(4):499-502.
[10]魯 成,余紅仕,向仲懷. 基于RAPD分析的中國野桑蠶和家蠶遺傳多樣性和系統發育關系研究[J]. 昆蟲學報,2002,45(2):198-203.
[11]夏慶友,周澤揚,魯 成,等. 家蠶不同地理品種分子系統學研究[J]. 昆蟲學報,1998,41(1):32.
[12]李 斌,魯 成,周澤揚,等. RAPD標記構建家蠶分子連鎖圖[J]. 遺傳學報,2000,27(2):127-132.
[13]何寧佳,魯 成,李 斌,等. 結合SADF與RAPD標記構建家蠶連鎖圖[J]. 昆蟲學報,2001,44(4):476-482.
[14]朱玉芳,譚遠德,萬春玲,等. 家蠶AFLP連鎖框架圖譜的構建[J]. 昆蟲學報,2001,44(4):483-493.
[15]桂慕燕,左正宏,王學民,等. RAPD分析在絹絲昆蟲親緣關系研究中的應用Ⅱ.柞蠶品種間的遺傳差異[J]. 遺傳,2001,23(5):452-454.
[16]劉彥群,魯 成,向仲懷. 4種體色柞蠶品種遺傳關系的RAPD分析[J]. 蠶業科學,2006,32(1):20-24.
[17]靳向東,朱興友,任曉義,等. 廣適性大型多絲量柞蠶新品種選育報告[J]. 北方蠶業,2016,37(1):1-6.
[18]農志歡,張啟偉,楊 平,等. 十大功勞屬6個種的遺傳多樣性及親緣關系RAPD分析[J]. 南方農業學報,2016,47(7):1077-1082.
[19]李 言,劉延嶺,崔翠菊,等. 海帶種質資源遺傳多樣性的RAPD分析[J]. 生物技術通報,2016,32(4):151-158.
[20]石洪玥,劉 陽,王曉梅,等. 七里海中華絨螯蟹遺傳多樣性的RAPD和ISSR分析[J]. 水產科學,2016,35(3):252-260.
[21]陳德西,封傳紅,何忠全,等. 四川省甘孜州9個不同地理蝗蟲種群遺傳多樣性的RAPD分析[J]. 西南農業學報,2016,29(6):1336-1342.
[22]宋娜娜,柴志欣,何世明,等. 三江黃牛RAPD遺傳多樣性研究[J]. 畜牧與獸醫,2016,48(6):30-35.
[23]Imazio S,Labra M,Grassi F,et al. Chloroplast microsatellites to investigate the origin of grapevine[J]. Genetic Resources and Crop Evolution,2006,53(5):1003-1011.
[24]王夢亮,任曉琳,崔晉龍,等. 野生紅景天的RAPD和ISSR遺傳多樣性分析[J]. 中草藥,2016,47(3):469-473.
[25]宋憲軍,聶 磊,張 濤,等. 柞蠶部分品種及雜交種的RAPD分析[J]. 蠶業科學,2004,30(4):428-431.
[26]趙菲佚,焦成瑾,李志明,等. RAPD法鑒定紫花苜蓿品種的條件優化[J]. 湖南農業科學,2016(1):1-5.
[27]曹蘭娟,楊寶山,李 俊,等. 不同地區栗蠶(Dictyoploca japonica)的RAPD的分析[J]. 蠶業科學,2007,33(2):293-296.
[28]Wang X L,Wang X L,Chang C R. RAPD analysis of Rosa laevigata Michx. from different origins[J]. Agricultural Biotechnology,2015,4(5):33-36.
[29]王 卓,曹蘭娟,劉振林,等. 不同地區及不同季節的柞蠶寄生蠅RAPD分析[J]. 蠶業科學,2008,34(2):354-358.
[30]王小武,付 開,丁新華,等. 基于RAPD標記的新疆荒漠稻區稻水象甲遺傳多樣性分析[J]. 新疆農業科學,2016,53(11):2069-2076.
[31]王鳳成,劉丹梅,仝振祥,等. 用RAPD標記分析部分柞蠶二化性品種資源的遺傳多樣性[J]. 蠶業科學,2009,35(1):148-153.
[32]郭曉麗,白麗榮. 10個小黑麥品種(系)的遺傳多樣性分析[J]. 河北農業科學,2016,45(2):26-28.
[33]程道軍,魯 成,周澤揚,等. 幾種絹絲昆蟲遺傳多樣性的RAPD研究[J]. 蠶業科學,2002,28(4):277-282.
[34]Gui M Y,Zuo Z H,Wang X M,et al. Application of RAPD technique in genetic relationship of silk insect. Ⅱ.Genetic variance in Antheraea pernyi[J]. Heredity,2001,23(5):452-454.
[35]鐘伯雄,張金衛,丁 農,等. RAPD技術在家蠶雜交率檢驗中的應用[J]. 浙江大學學報(農業與生命科學版),2003,29(3):321-324.
[36]Nei M.Estimation of average heterozygosity and genetic distance from a small number of individuals[J]. Genetics,1978,89(3):583-590.
[37]陳瑩玉,袁思思,吳春蘭,等. 廣西金秀野生茶遺傳多樣性及分子指紋圖譜研究[J]. 廣東農業科學,2016,43(3):21-28.
[38]劉彥群,魯 成,向仲懷. 中國柞蠶DNA多態性的RAPD分析[J]. 蠶業科學,2002,28(4):283-288.
[39]喬利MS,林華生. 柞蠶屬種的分布與分化[J]. 國外農學-蠶業,1983(4):3-9.
