秦巴硒菇提取物對人肝癌細胞株SMMC-7721增殖與凋亡的調節作用的實驗研究
2019-07-29 05:52:16袁作輝王倩鳳
衛生職業教育 2019年16期
關鍵詞:肝癌
李 娟,袁作輝,楊 帆,王倩鳳
(甘肅省人民醫院干部病房,甘肅 蘭州 730000)
肝癌是起源于肝臟的上皮或間葉組織的惡性腫瘤[1],近年來肝癌的發病率在全球范圍內均有增加的趨勢[2],據2018年全球癌癥數據統計,2018年全球癌癥死亡率最高的五大癌癥中就包括肝癌[3-4],肝癌新增人數達841 080例,占比4.7%,肝癌死亡人數781 631,占比8.2%,肝癌在我國尤為高發,是造成我國癌癥死亡的第四大原因[5-7],肝癌死亡率是僅次于胃癌和食管癌的第三大消化系統惡性腫瘤[8-9],并且其發病率呈逐年上升的趨勢。肝癌的早期發病隱匿,病程進展至中后期會迅速侵襲轉移,惡性程度較高,目前臨床上進行手術切除和肝移植以及術后聯合放化療是治療的主要手段[10-12],但僅有30%的肝癌患者適合手術治療,但因超過80%的患者就診時已是晚期,常常錯過最佳的手術治療時機,即使經過治療也不能較好地改善患者生活質量,并且術后復發率高,5年生存率僅有15%,預后差[13-14],嚴重威脅著人類的身體健康。因此,尋找新的肝癌治療藥物是延長肝癌患者生存時間、降低肝癌死亡率的關鍵。探尋以抑制肝臟腫瘤細胞增殖、誘導凋亡為切入點,探究一種新型、安全且有效的抗腫瘤藥物成為目前肝癌治療的新思路,也是提高臨床療效及輔助外科治療的新方法[15-18]。我國有豐富的中藥來源,一些植物提取物成為抗腫瘤藥物的重要來源,例如苦參堿、紫杉醇、秦巴硒菇提取物等。因此,近年來天然產物的抗癌活性受到廣泛關注[19]。其中,中藥秦巴硒菇(AgaricusblazeiMurrill-Qbsg)中富含β-葡聚糖,對自身免疫系統有著很強的刺激作用。在試管培養及動物模型中,秦巴硒菇中的β-葡聚糖都顯示出了抗腫瘤作用。從秦巴硒菇多糖中提取的酸性RNA蛋白復合物(FA-2-bβ)具有較強的抗腫瘤活性,具有抑制肝癌細胞SMMC-7721增殖的活性,FA-2-b-β有抗感染、調節機體免疫功能、抗過敏、修復肝損傷、促進腫瘤細胞凋亡的效果[20-21]。因此,本研究利用體外培養肝癌細胞SMMC-7721,并給予不同濃度的秦巴硒菇提取物處理,探究秦巴硒菇提取物對肝癌細胞增殖、凋亡能力的影響,并進一步探究其作用機制。
1 材料與方法
1.1 材料
1.1.1 細胞、藥物及試劑 人肝癌SMMC-7721細胞株購自中國醫學科學院基礎醫學研究所,秦巴硒菇、胰蛋白酶、甲基噻唑藍(MTT)、胎牛血清DMEM完全培養基購于美國Sigma公司,AnnexinV-FITC/PI細胞凋亡檢測試劑盒購自北京寶賽生物技術有限公司,反轉錄試劑盒及RT-qPCR試劑盒購于TAKARA公司,抗 P53、Caspase3、Caspase9、內參一抗甘油醛-3-磷酸脫氫酶(glyceraldehyde-3-phosphate dehydrogenase)、β-actin購自上海英駿生物技術有限公司,二抗購于北京索萊寶有限公司。
1.1.2 主要儀器 多功能酶標儀(美國Thermo公司)、CO2培養箱(中國力康生物公司)、超凈工作臺(Thermo Forma)、倒置顯微鏡(OlympusBX51)、流式細胞儀(德國ParcGmbh,CyFlow Space公司)、低溫離心機HITACH(日本CF-16RX公司)、自動細胞計數儀(Invitrogen Countess)、BIO-RAD CFX96實時熒光定量PCR儀、高速冷凍離心機(美國Thermo scientific公司)、全波長分光光度計(上海UNICO公司)、Trizol及RIPA裂解液(美國MRCMolecular Research Center公司)、自動蒸汽消毒鍋(日本SANYO公司)、-80℃超低溫冰箱(美國Thermo公司)、凝膠成像儀、垂直電泳槽、濕式轉膜槽 (美國BIO-RAD公司)、3 mmWhatman濾紙(美國GE公司)
1.2 方法
1.2.1 細胞培養 人肝癌SMMC-7721細胞用含10%胎牛血清的DMEM完全培養基,置于37℃含5%CO2的飽和濕度培養箱內培養,每日觀察細胞生長情況,每隔1~2天換液一次,棄去鏡下觀察細胞生長情況欠佳及有污染的細胞株,剩余為實驗用。待細胞生長達80%左右時對細胞進行不同濃度秦巴硒菇提取物處理并進行后續實驗。
1.2.2 細胞增殖抑制率 取對數生長期SMMC-7721細胞,以1×105/mL細胞濃度種植96孔細胞培養板中,常規培養24 h,DMEM完全培養基稀釋秦巴硒菇提取物至濃度分別為0.5,1.0,2.0,4.0 μg/mL,陰性對照組加入 100 μL DMEM 完全培養基,每組設3個復孔。秦巴硒菇提取物處理48 h后,小心吸棄上層含藥物培養基。每孔加入10%MTT的完全培養基繼續培養4 h,棄掉上清液,加入DMSO,振蕩溶解10 min。在490 nm波長處測定吸光度值(OD值)。細胞增殖抑制率(%)=(1-實驗組細胞OD值/陰性對照組細胞OD值)×100%。
1.2.3 細胞凋亡檢測 人肝癌SMMC-7721細胞種植于細胞60 mm培養皿中,待細胞增長達對數生長期后,分別加入濃度分別為 0.5,1.0,2.0,4.0 μg/mL 的秦巴硒菇提取物,繼續培養 48h,以胰酶消化并收集細胞,用高速離心機1000 r/min離心5 min,PBS清洗兩次,以Annexin V-FITC在暗室中孵育30 min,然后加入2 μL的PI(100 μg/mL),用流式細胞分析檢測SMMC-7721細胞凋亡情況。
1.2.4 實時定量PCR 取對數生長期細胞以2.5×105/mL的濃度種植于60 mm培養皿中,培養24 h后分別加入不同濃度秦巴硒菇提取物,使終濃度為 0.5,1.0,2.0,4.0 μg/mL,繼續培養48 h后,加入1 mL Trizol裂解細胞并收集細胞于無酶EP管中,加入0.2 mL的三氯甲烷,劇烈震搖EP管25~30次,4℃離心15 min,取上清并加入等體積的異丙醇,輕搖EP管10次,放置-20℃冰箱,離心后吸棄去上清液,加入1 mL75%無水乙醇,重懸后離心,吸棄上清,室溫自行干燥,加入適量無RNA酶水,室溫放置10 min。NANODROP2000中檢測RNA濃度及純度。由于RNA保存不當時宜降解,影響其濃度及純度,遂取2 μgRNA按照TAKARA公司試劑盒反應體系說明書,立即將RNA反轉錄成cDNA。以cDNA為模板,按照TAKARA公司的qPCR試劑盒反應體系加樣,在實時熒光定量PCR儀進行PCR反應,反應條件為:95℃30 sec;95℃5 sec;60℃30 sec共 45個循環。β-actin作為管家基因進行校準。采用2-ΔΔCt相對定量法分析結果,ΔΔCt=實驗組(Ct目的基因-Ct管家基因)-對照組(Ct目的基因-Ct管家基因),相對表達=2-ΔΔCt。
1.2.5 Western blot檢測蛋白表達 不同濃度秦巴硒菇提取物作用于細胞后,收集細胞,用RIPA裂解液提取蛋白,用BCA試劑盒檢測蛋白濃度并調平。取等量的蛋白質用10%SDS-聚丙烯酰胺凝膠電泳跑膠進行分離,蛋白轉移至PVDF膜,5%脫脂奶粉封閉室溫搖床上1 h,TBST稀釋一抗4℃孵育過夜(一抗的稀釋比例下:P53:1∶1 200;Caspase3:1∶1 000;Caspase9:1∶1 000;β-actin:1∶2 000),復溫 1 h,TBST 洗膜 3 次,加入 TBST 稀釋的二抗(二抗稀釋比例1∶1 000),室溫孵育1 h,TBST洗膜3次。將PVDF膜浸在ECL超敏發光液中1 min,在ECL發光儀上進行曝光成像。凝膠成像系統獲取蛋白質條帶圖片,ImageJ對蛋白質條帶灰度值進行定量分析。
1.3 統計學處理
2 結果
2.1 秦巴硒菇提取物對肝癌SMMC-7721細胞增殖的影響
秦巴硒菇提取物對人肝癌SMMC-7721細胞的增殖有明顯的抑制作用,且隨秦巴硒菇提取物的濃度和時間的作用增加而呈現出抑制作用更加明顯,與對照組比較,差異有統計學意義(P<0.05)。結果見表 1。

表1 秦巴硒菇提取物對人肝癌SMMC-7721細胞不同濃度和時間的增殖抑制率
2.2 秦巴硒菇提取物對人肝癌SMMC-7721細胞凋亡的影響
不同濃度的秦巴硒菇提取物作用于人肝癌SMMC-7721細胞24 h后,采用流式細胞分析檢測細胞凋亡率的變化,結果顯示不同濃度的秦巴硒菇提取物 (0.5,1,2,4 μg/mL) 作用于SMMC-7721細胞后,凋亡率明顯高于對照組(0 μg/mL),且隨秦巴硒菇提取物濃度增加細胞凋亡率明顯增高,差異有統計學意義(P<0.05),結果見表 2。

表2 秦巴硒菇提取物對人肝癌SMMC-7721細胞凋亡的影響
2.3 Annexin V/PI凋亡檢測結果
應用Annexin V/PI、FCM法對秦巴硒菇提取物作用于SMMC-7721細胞作用后的凋亡現象分別進行兩次定性定量的測定。流式細胞散點圖分為4個象限,象限左下角LL代表正常細胞簇群,象限左上角UL代表機械損傷或壞死細胞簇群,象限右下角LR代表早期凋亡細胞簇群,象限右上角UR代表晚期凋亡或死亡細胞簇群。兩次對照組細胞與其他組細胞均存在顯著性差異(P<0.05)。見圖 1~2。
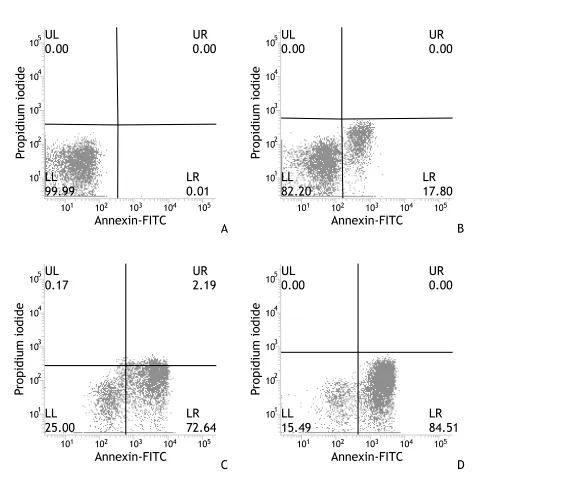
圖1 秦巴硒菇提取物對人肝癌SMMC-7721細胞凋亡的作用(第一組)
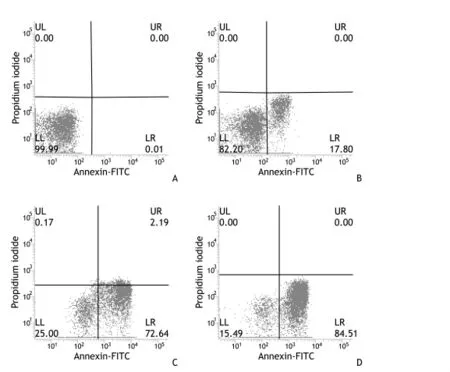
圖2 秦巴硒菇提取物對人肝癌SMMC-7721細胞凋亡的作用(第二組)
2.4 秦巴硒菇提取物對P53/Caspase信號通路的影響
從條帶可以看出,各組的條帶深淺基本一致,說明各組的細胞總量相同。本實驗結果提示不同濃度的秦巴硒菇提取物均能誘導SMMC-7721細胞發生凋亡。與對照組相比,以β-actin為內參蛋白。Western Blot實驗結果顯示,秦巴硒菇提取物誘導了P53、Caspase-3及Caspase-9蛋白的表達,最終誘導細胞凋亡。遞增濃度的秦巴硒菇提取物對人肝癌SMMC-7721細胞P53、Caspase3及Caspase9蛋白的影響,差異有統計學意義(P<0.05),見圖3,表明秦巴硒菇提取物可誘導肝癌細胞P53/Caspase信號通路的激活。

圖3 遞增濃度的秦巴硒菇提取物對人肝癌SMMC-7721細胞P53、Caspase3及Caspase9蛋白的影響
3 討論
3.1 秦巴硒菇提取物具有抑制肝癌細胞增殖、誘導凋亡的作用
細胞增殖不受控制是癌癥發生的主要機制之一。誘導癌細胞凋亡是大多數抗癌藥物抑制癌癥發展的作用機制。近年來大量研究發現,多類天然產物提取物也能通過誘導癌細胞凋亡發揮抗癌作用[22]。秦巴硒菇提取物為天然產物提取物,可抑制多種癌癥細胞的增殖,誘導癌癥細胞發生細胞周期阻滯,抑制細胞增殖相關通路激活,從而減緩腫瘤的生長,發揮抗癌作用[23]。本文研究也發現,高濃度的秦巴硒菇提取物能降低肝癌SMMC-7721細胞的活性。在無明顯細胞毒性的劑量下,秦巴硒菇提取物作用于肝癌細胞的時間長短與濃度大小與肝癌細胞增殖倍數呈負相關,顯著升高肝癌細胞的凋亡率,表現為劑量依賴性與時間依賴性,進一步提示秦巴硒菇提取物具有抑制肝癌細胞增殖、誘導凋亡的作用[24-25]。
3.2 秦巴硒菇提取物可誘導P53/Caspase信號通路的激活
細胞凋亡是指細胞發生程序性死亡,由多條信號通路共同調控,這些信號通路主要通過改變線粒體通透性,誘導線粒體釋放凋亡誘導因子而啟動細胞凋亡過程[26-27]。Caspase家族是細胞凋亡的最終執行者,Caspase家族蛋白的激活可誘導DNA降解、介導DNA損傷,最終導致細胞凋亡[28]。Caspase3是凋亡的執行分子,是細胞凋亡過程中最主要的終末剪切酶。它是由CASP3基因編碼的,外部死亡受體途徑和內在線粒體途徑啟動后共同的效應酶,在凋亡肝癌細胞中明顯表達[29]。而Caspase9由CASP9基因編碼的凋亡啟動蛋白酶,通過磷酸化進行調控激活,引發線粒體內部活化凋亡途徑。誘導細胞凋亡是癌癥治療中細胞死亡的首選模式,是癌癥治療關鍵。P53是Caspase家族蛋白的上游調控因子,在氧化應激、紫外照射等多種應激條件誘導DNA損傷過程中均呈高表達狀態[30]。P53作用于線粒體可誘導細胞色素C表達,細胞色素C可直接誘導Caspase9活化,活化的Caspase9可進一步促進Caspase3激活,從而導致細胞凋亡[31-32]。P53介導的P53/Caspase細胞凋亡信號通路是多類化療藥物治療癌癥的重要機制之一[33]。既往有研究表明,黑種草種子提取物可通過激活P53/Caspase信號通路誘導乳腺癌細胞凋亡,紫草素可通過誘導P53表達升高Caspase-9表達水平,從而誘導細胞凋亡。本研究采用Western blot法檢測了秦巴硒菇提取物處理肝癌細胞后,細胞P53、Caspase3和Caspase9的蛋白表達水平。結果顯示,秦巴硒菇提取物在誘導P53表達的同時也能升高肝癌細胞Caspase3和Caspase9的表達水平,提示其能夠誘導P53/Caspase信號通路的激活,可能是秦巴硒菇提取物誘導肝癌SMMC-7721細胞凋亡的作用機制之一。
綜上所述,秦巴硒菇提取物可抑制肝癌細胞增殖、誘導肝癌細胞凋亡,并能促進凋亡蛋白Caspase3和Caspase9的表達,其作用機制可能與誘導P53/Caspase信號通路的激活有關。本研究僅初步探討了秦巴硒菇提取物抗肝癌的作用機制,之后將通過體內實驗驗證并進一步探究其具體的分子作用機制,為臨床開發新的肝癌治療藥物奠定實驗基礎。
猜你喜歡
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學學報(醫學版)(2015年1期)2015-12-17 07:47:28
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
