廬山不同海拔茶樹光合響應差異研究
2019-08-21 01:32:42程冬梅張麗韋紅飛江新鳳周賽霞張志勇彭焱松
茶葉科學 2019年4期
程冬梅,張麗,韋紅飛,江新鳳,周賽霞,張志勇,彭焱松*
廬山不同海拔茶樹光合響應差異研究
程冬梅1,張麗1,韋紅飛2,江新鳳3,周賽霞1,張志勇1,彭焱松1*
1. 中國科學院廬山植物園,江西省植物遷地保護與利用重點實驗室,江西 九江 332900;2. 九江市廬山茶葉科學研究所,江西 九江 332900;3. 江西省蠶桑茶葉研究所,江西 南昌 330202
為明確茶樹光合活性和光系統Ⅱ(PSⅡ)的光抑制對海拔梯度的響應規律,以茶樹品種廬山本地群體種為材料,觀測了廬山5個不同海拔高度茶樹葉片的光合氣體交換和葉綠素熒光參數。結果表明,葉片厚度、最大凈光合速率(Pnmax)、PSⅡ最大相對電子傳遞速率[rETR(Ⅱ)max]、光補償點(Ic)、光飽和點(Isat)、半飽和光強(Ik)、水分利用效率(WUE)等隨海拔升高呈增加趨勢,水汽壓虧缺(VPD)、氣孔導度、蒸騰速率、胞間CO2濃度、暗呼吸速率(Rd)等隨海拔升高而呈下降趨勢。PSⅡ潛在最大光化學量子產量(Fv/Fm)隨海拔升高而增加;PSⅡ非調節性能量耗散量子產量[Y(NO)]隨海拔升高而降低。PSⅡ實際光化學量子產量[Φ(Ⅱ)]、光化學淬滅(qP)、PSⅡ相對電子傳遞速率[rETR(Ⅱ)]均隨海拔升高而增加。綜上分析,茶樹在廬山低海拔處的光合作用和PSⅡ活性顯著低于高海拔處,這與低海拔處葉片PSⅡ受到光抑制密切相關。
廬山云霧茶;海拔梯度;光合作用;氣體交換;葉綠素熒光;光抑制
茶樹(L.)屬于山茶科山茶屬多年生小喬木或灌木,野生種常見于中國長江以南各省的山區,栽培廣泛,種類豐富[1]。廬山云霧茶屬中國十大名茶之一,大多產自廬山本地群體種,因茶樹生長于廬山的流泉飛瀑、行云走霧中,以其獨特的口感和品質而聞名。廬山主峰最高海拔1?474?m,茶樹在不同海拔高度均有分布。由于溫度、濕度、光照等多種環境因子均隨著海拔梯度而發生劇烈變化[2-3],因此茶樹的生長發育、生理代謝與品質在不同海拔也必然呈現出不同的響應特征。研究表明,廬山云霧茶酚氨比與海拔高度成反比,咖啡堿含量與海拔相關性不顯著,霧日數是影響茶葉品質的限制性因子[4]。
光合作用作為植物最重要的生理代謝活動,產生的糖類不僅作為碳源為其他生理活動提供必需的能量[5-6],而且作為信號分子參與類黃酮生物合成等代謝活動[7],因此光合作用直接影響茶樹的產量與品質。光合作用的光反應階段是在位于類囊體膜上的光合機構組成的光合電子傳遞鏈上進行的,光合機構在結構和功能上均具有復雜精細的調控過程,對溫度、光照、濕度等環境變化十分敏感[8-10]。光系統Ⅱ(PSⅡ)對光照最為敏感,較高的光強容易對PSⅡ產生光抑制[11-12],而干旱、高溫、低溫等其他脅迫則會加重光對PSⅡ的光抑制程度[13-15]。因此,研究茶樹的光合活性和PSⅡ光抑制程度隨海拔梯度的變化規律,對評價不同海拔茶樹生長發育及茶葉產量與品質、指導茶樹的科學種植有重要的實踐意義。研究表明,茶樹品種和海拔幅度的差異都影響到光合強度的變化[16-17],然而,目前廬山本地群體種茶樹光合活性尤其是PSⅡ光抑制程度隨海拔梯度變化的規律尚未見報道。
本試驗沿廬山海拔梯度選取5個茶園的本地群體種茶樹,于8月9日—24日分別觀測茶樹冠層葉片光合氣體交換和葉綠素熒光參數,以此揭示茶樹的光合作用活性和PSⅡ的光抑制程度隨海拔梯度的變化規律。
1 材料與方法
1.1 試驗材料
試驗在廬山5個沿海拔梯度分布的茶園中開展,5個茶園的海拔高度、地理位置、氣候等信息見表1。試驗供試茶樹品種均為廬山本地群體種,種植方式為單行條栽或雙行條栽。選擇樹齡均在30?a左右、高度50?cm左右、無遮陰的健康茶樹,于2018年8月9日、8月10日、8月21日、8月23日、8月24日晴朗無云的天氣上午10:30開始觀測光合作用氣體交換和葉綠素熒光參數。所有觀測的葉片均選當年生的夏梢頂芽以下第三和第四片葉。
1.2 環境參數及葉片厚度測量
海拔與地理位置數據由手持式GPS獲得,上午10:30茶樹冠層的光合有效輻射由LI-COR 1500觀測獲得,上午10:30茶樹冠層的氣溫、水汽壓虧缺由LI-COR 6800便攜式光合儀記錄。茶樹葉片厚度用游標卡尺測量,測量部位均選葉片最寬處并避開中脈。每個海拔茶園均隨機選取30片葉片進行測量,求平均值和方差。
1.3 光合氣體交換與葉綠素熒光檢測
葉片氣體交換和葉綠素熒光參數由LI-COR 6800便攜式光合儀根據儀器使用說明書進行檢測。使用提供人工光源的多相閃光熒光葉室,設置葉室溫度27℃,流速500?μmol·s-1,空氣相對濕度50%~75%,含CO2量400?μmol·mol-1,風扇轉速為10?000?r·min-1。上午10:30取自然光適應的葉片夾在葉室中,在1?800?μmol·m-2·s-1人工光源(紅光︰藍光為9︰1)下充分適應,待各參數穩定后開始記錄凈光合速率等氣體交換參數。根據測量數據計算水分利用效率=凈光合速率/蒸騰速率。然后取自然光適應的葉片在葉室1?800?μmol·m-2·s-1人工光源下充分適應后,分別在1?800、1?500、1?200、900、600、300、200、150、100、70、30、0?μmol·m-2·s-1的光強下進行光合速率的自動光響應曲線觀測,每個光強下持續120~180?s。提前用鋁箔紙完全包裹葉片暗適應60?min以上,觀測葉片的最小熒光F0和10?000?μmol·m-2·s-1飽和脈沖下最大熒光Fm,并計算PSⅡ潛在最大光化學量子產量Fv/Fm=(Fm-F0)/Fm。然后開啟500?μmol·m-2·s-1的光化光,待葉片在葉室中充分適應后檢測光適應時最小熒光F0'、最大熒光Fm'和穩態熒光Fs,分別計算PSⅡ實際光化學量子產量Φ(Ⅱ)=(Fm'-Fs)/Fm';PSⅡ相對電子傳遞速率rETR(Ⅱ)=光合有效輻射(PAR)×Φ(Ⅱ)×0.84×0.5;PSⅡ非調節性能量耗散量子產量Y(NO)=Fs/Fm;光化學淬滅qP=1-(Fs-F0')/(Fm'-F0')。葉綠素熒光參數的光響應曲線分別在0、25,50、100、150、300、500、1?000、1?500、2?000?μmol·m-2·s-1光強下進行自動觀測,每個光強下持續120~180?s。氣體交換和葉綠素熒光及其光響應曲線均至少觀測5個葉片,求平均數與方差。
1.4 數據統計分析
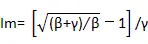
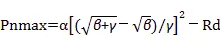
PSⅡ相對電子傳遞速率的光響應曲線用Platt等[19]提出的雙指數方程進行擬合:
式中,P為rETR(Ⅱ),Pm為rETR(Ⅱ)max,α為初始斜率,PAR為光合有效輻射,β為光抑制參數。
Ik=rETR(Ⅱ)max/α,表示葉片耐受強光的能力。
上述光響應曲線用Origin 8.5進行非線性擬合并通過坐標圖來表達。
2 結果與分析
2.1 不同海拔茶樹葉片光合活性變化
5個不同海拔高度茶樹葉片的光合氣體交換過程均在8月9日—8月24日晴朗無云的上午10:30開始觀測,此時日照輻射強度均已達到1?400~2?000?μmol·m-2·s-1,且隨著廬山海拔的升高,日照輻射強度逐漸上升,氣溫逐漸下降,水汽壓虧缺呈現下降趨勢,即空氣濕度隨海拔升高逐漸增加(表1)。圖1顯示隨著海拔的升高,茶樹葉片溫度下降,葉片厚度增加,且葉片溫度、葉片厚度與海拔分別呈顯著的負相關和正相關。凈光合速率隨海拔上升呈現增加的趨勢,蒸騰速率則隨海拔上升而呈下降趨勢,因此水分利用效率隨海拔升高而增加;隨著海拔的升高,氣孔導度略有下降,胞間CO2濃度呈降低趨勢,胞間CO2濃度與大氣CO2濃度的比值逐漸降低(圖1)。通過直角雙曲線修正模型模擬凈光合速率的光響應曲線(圖2),并計算出光響應曲線的初始斜率、最大凈光合速率、飽和光強、光補償點、暗呼吸速率等參數(表2)。結果表明,模擬的決定系數較高,模擬結果較好。初始斜率和最大凈光合速率在最低海拔處(240?m)數值最低,當海拔升高至460?m時,初始斜率和光合速率則顯著升高,且隨著海拔進一步升高而保持較穩定水平。飽和光強、光補償點和暗呼吸速率則隨著海拔升高而逐漸升高,在海拔1?100?m達到最高水平。
2.2 不同海拔茶樹葉片光抑制變化
葉綠素熒光參數Fv/Fm和Y(NO)可以反映PSⅡ的光抑制程度。圖3顯示,在5個海拔梯度中,茶樹葉片的Fv/Fm在最低海拔處(240?m)為0.76,海拔上升至460?m時Fv/Fm增加至0.79,且隨著海拔的進一步升高,維持在0.80的水平。而Y(NO)則正好相反,海拔240?m處Y(NO)為0.43,海拔上升至460?m時Y(NO)下降至0.31,在最高海拔處(1?100?m)Y(NO)下降至0.27。海拔240?m處Fv/Fm的最低值和Y(NO)的最高值反映出茶樹葉片PSⅡ在低海拔受到一定程度的光抑制,且PSⅡ光抑制程度隨著海拔的升高而逐漸減小。
2.3 不同海拔茶樹葉片PSⅡ活性變化
為了進一步檢測不同海拔茶樹葉片PSⅡ的活性,用雙指數方程擬合PSⅡ相對電子傳遞速率的光響應曲線(圖4),并計算出曲線的初始斜率、最大電子傳遞速率、半飽和光強等參數(表3)。曲線初始斜率、最大相對電子傳遞速率和半飽和光強都在最低海拔處(240?m)達到最低值,隨著海拔升高而增加。曲線初始斜率反映了光合器官對光能的利用效率,半飽和光強反映出葉片對強光的耐受能力,因此高海拔地區茶樹葉片的光合器官活性和耐強光能力都高于低海拔葉片。此外,PSⅡ實際量子產量Φ(Ⅱ)和光化學淬滅qP的光響應曲線也表現出與相對電子傳遞速率的光響應曲線相似的變化,即Φ(Ⅱ)和qP在海拔240?m處位于最低水平,隨海拔的升高而逐漸增加,在海拔1?100?m處達到最高水平(圖5)。
南通之所以能成為長壽之鄉,主要不在于其所處的地理位置亞熱帶氣候四季分明、其經濟發展水平,如果是上述因素主要,很多地區和南通類似,但他們那里長壽群體(80歲以上者)占總人口比例并不高。更主要在于南通地區的文化傳統、社會意識、風俗習慣。
德氏乳桿菌(Lactobacillus delbrueckii)亞種保加利亞乳桿菌是使用D-乳酸脫氫酶(DLDH)將丙酮酸鹽主要轉化為D-乳酸的非均勻乳酸菌,其功能特性仍然不佳,進一步的研究表明,乳酸脫氫酶活性受氧下調,因此在厭氧條件下達到最高的D-乳酸滴度為1.94 g/L[29]。D-乳酸是德氏乳桿菌的主要乳酸產物,但不能被人體腸道代謝,Zhang J等[30]進一步研究了氨基酸水平上D-LDH和L-LDH基因的進化,發現德氏乳桿菌D-LDH基因是正向選擇的,可能是長期馴化的結果。Viana R[31]發現干酪乳桿菌有兩種特征基因編碼乳酸脫氫酶活性,它特別催化了L-乳酸的形成。
表1 不同海拔茶園地理位置及觀測時間點氣候信息 Table 1 Geographic and climatic information of five tea gardens of different altitudes 序號No.茶園名稱Names海拔/mAltitude經緯度N/ELatitude/Longitude坡向Slope氣溫Air temperature/℃日照強度PAR/μmol·m-2·s-1水汽壓虧缺VPD/kPa 1圓通茶園24029°31' 54"/115°53'36"南32.61±0.221?641±213.45±0.22 2馬尾水茶園46029°37' 40"/116°01'35"西北31.90±0.241?465±121.59±0.46 3王家坡茶園58029°35' 52"/116°01'33"東30.88±0.161?747±101.79±0.10 4小天池茶園90029°35' 03"/115°59'55"北30.83±0.211?894±801.17±0.08 5廬山植物園茶園1?10029°33' 03"/115°58'46"東南30.07±0.191?908±122.13±0.11
注:表中氣候信息測定次數均至少3次
Note: Climatic parameters were measured at least three times
注:圖中*表示相關性顯著( P<0.05) Note: * represents significant correlation (P<0.05) 圖1 不同海拔茶樹葉片光合氣體交換參數 Fig. 1 Photosynthetic gas exchange parameters of tea leaves at five altitudes
圖2 不同海拔茶樹葉片光合速率的光響應曲線擬合 Fig. 2 Fitted light response curve of photosynthesis of tea leaves at five altitudes
表2 不同海拔茶樹葉片光合速率的光響應擬合數據 Table 2 Parameters of fitted photosynthesis of tea leaves at five altitudes 海拔Altitude/m初始斜率Initial slope α最大凈光合速率Pnmax/μmol·m-2·s-1光飽和點Isat/μmol·m-2·s-1光補償點Ic/μmol·m-2·s-1暗呼吸速率Rd/μmol·m-2·s-1決定系數R2Determination coefficient 2400.0698.101?899.2015.81-1.090.996?7 4600.07712.951?850.9019.11-1.470.998?5 5800.07816.321?809.5325.90-2.020.998?1 9000.0809.861?944.7117.50-1.400.997?0 1?1000.07711.892?905.6830.45-2.340.998?7
圖3 不同海拔茶樹葉片PSⅡ光抑制 Fig. 3 Photoinhibition of PSⅡ of tea leaves at five altitudes
3 討論
本試驗在廬山5個不同海拔的茶園中觀測了茶樹葉片光合氣體交換和葉綠素熒光,以揭示茶樹葉片光合作用生理活性和光合機構對海拔梯度的響應。隨著海拔的上升,氣溫降低、光照輻射增加、CO2分壓下降(圖1,表1),這些環境因素看似不利于光合作用,但茶樹葉片在高海拔地區的光合生理過程和光合機構活性整體上均高于低海拔地區,且胞間CO2濃度隨海拔升高呈下降趨勢,即CO2的利用率更高。基于植物生理生態學中的“最小成本假說”及其標準模型,高海拔地區葉片更低的胞間CO2與大氣CO2濃度之比和更高的羧化能力可以最低限度地減小碳同化的損耗,實現最大的碳同化能力[2,20]。值得注意的是,盡管光合活性總體趨勢沿海拔升高而增加,但并非線性增加(表2)。這表明除了海拔因素,坡向、生境等不同環境因素及其引起的光照、溫度、降雨等氣候條件的動態變化也會影響光合作用過程。此外,葉片的光補償點和光飽和點隨著海拔的升高而增加,體現了茶樹對高海拔地區強光輻射環境的適應和對光能較高的利用能力。這種規律在其他植物中也得到了廣泛報道[21-22]。
隨著廬山海拔的升高,茶樹葉片厚度呈增加趨勢。這種變化趨勢與云南大葉種茶樹的研究結果類似[17],體現了植物葉片對高海拔地區低溫環境的適應性[23-24]。光合同化速率受茶樹品種、海拔高度范圍、氣溫、水分等多種因素影響。福鼎大白茶在氣溫20℃左右時,隨著海拔從615?m升高到1?115?m,光合強度呈下降趨勢[16]。廬山本地群體種從海拔240?m上升到1?100?m時,氣溫從32℃下降到30℃左右,光合速率總體呈上升趨勢,且在900~1?100?m保持較高水平。這與云南大葉種茶樹在27℃左右、700~1200?m的海拔范圍內光合同化速率保持較高水平且差異不大的趨勢一致[17]。廬山本地群體種茶樹胞間CO2濃度和氣孔導度從240?m低海拔處到1?100?m的亞高山呈緩慢下降趨勢,而云南大葉茶隨著海拔進一步升高到1?500~2?000?m,光合速率逐漸下降,氣孔導度從最高值下降,胞間CO2濃度穩定[17]。這些結果說明,海拔幅度的變化導致的茶樹光合作用氣體交換過程的變化規律仍需要在更寬的海拔范圍尺度上進行研究。胞間CO2濃度與氣孔導度的共同變化可以反映光合速率的下降主要是受氣孔限制還是非氣孔因素限制[25]。由于隨海拔下降光合速率的下降伴隨著胞間CO2濃度的升高,因此限制廬山本地群體種茶樹葉片光合活性的主要限制因子是非氣孔因素,即葉肉細胞的活性。
圖4 不同海拔茶樹葉片相對電子傳遞速率的光響應曲線擬合 Fig. 4 Fitted light response curve of rETR(Ⅱ) of tea leaves at five altitudes
表3 不同海拔茶樹葉片相對電子傳遞速率的光響應擬合數據 Table 3Parameters of fitted rETR(Ⅱ) of tea leaves at five altitudes 海拔Altitude/m初始斜率Initial slope α最大相對電子傳遞速率rETR(Ⅱ)max半飽和光強Ik/μmol·m-2·s-1決定系數Determination coefficient R2 2400.32869.94213.510.999?6 4600.32585.01261.200.999?8 5800.34370.00203.940.999?3 9000.32982.37250.720.999?6 1?1000.34686.28249.270.999?6
圖5 不同海拔茶樹葉片Φ(Ⅱ)和qP光響應曲線 Fig. 5 Light response curve of Φ(Ⅱ) and qP of tea leaves at five altitudes
在觀測的5個海拔的茶園中,最低海拔(240?m)的茶樹葉片總體光合活性均最低,而海拔升至460?m及以上時,光合活性顯著升高,因此240?m及更低的海拔高度下茶樹的光合活性相對較低。與光合活性一致的是,240?m海拔處茶樹葉片PSⅡ光抑制程度相對最高(圖2)。溫度、光照、濕度等氣候環境與茶樹的生長發育和生理代謝息息相關[4,25]。比較低海拔與高海拔處茶園的氣候參數可見,240?m海拔處氣溫高達32.6℃,水汽壓虧缺最高,此處茶樹在觀測條件下相對其他高海拔茶園處于高溫、干旱的脅迫。茶樹對溫度、濕度條件非常敏感,高溫干旱脅迫影響茶樹的生長發育與生理代謝[26-27]。研究報道福鼎大白茶夏季光合適溫為30℃[28]。因此,夏季低海拔茶園高溫干旱條件可能對茶樹光合作用過程和光合機構產生一定的抑制。馬力等[4]研究證明,海拔1?165?m的廬山氣候觀測站觀測到的降雨量、霧日數均比山腳下觀測到的顯著增加,高海拔地區較高的降雨量和云霧量是“高山云霧出好茶”的重要條件。本試驗也證明高海拔云霧茶夏季因享有適宜的溫度和較高的濕度而保持較高的光合作用活性,光合機構避免受到光抑制而保持高效運轉。
隨著全球氣候變化的加劇和極端天氣的頻發[29-30],夏季普遍持續的高溫天氣對茶樹生長發育和生理代謝的影響日益凸顯。全球氣候變暖不僅直接通過高溫干旱脅迫影響低海拔植物光合活性和光合機構的正常運轉,而且對高海拔地區產生更高的變暖速率從而深刻影響高海拔地區植物的光合作用[31-32]。在茶樹的生產實踐中,夏季保持適宜的溫度和較高的濕度,是提高光合活性、保持光合機構正常運轉的重要前提,也是保證茶葉產量和品質的基礎。
參考文獻
[1] 張宏達, 任善湘. 中國植物志第49卷第3冊[M]. 北京: 科學出版社, 1998: 130.
[2] Wang H, Prentice I C, Davis T W, et al. Photosynthetic responses to altitude: an explanation based on optimality principles [J]. New Phytologist, 2017, 213(3): 976-982.
[3] 羅旭. 不同高海拔對金冠蘋果光合特性和果實品質的影響[D]. 雅安: 四川農業大學, 2013: 5-6.
[4] 馬力, 黃紀剛, 吳昊. 廬山云霧茶氣候品質分析[J]. 中國高新區, 2017(23): 229-230.
[5] Rabinowitch E. Photosynthesis [J]. Annual Review of Physical Chemistry, 1951, 2(1): 361-382.
[6] Galston A W. Photosynthesis as a basis for life support on earth and in space [J]. Bioscience, 1992, 42(7): 490-494.
[7] 宛曉春, 夏濤. 茶樹次生代謝[M]. 北京: 科學出版社, 2015: 51-100.
[8] Umena Y, Kawakami K, Shen J R, et al. Crystal structure of oxygen-evolving photosystem Ⅱ at a resolution of 1.9 A? [J]. Nature, 2011, 473(7345): 55-60.
[9] Ja?rvi S, Suorsa M, Aro E M. Photosystem Ⅱ repair in plant chloroplasts-regulation, assisting proteins and shared components with photosystem Ⅱ biogenesis [J]. BBA-Bioenergetics, 2015, 1847(9): 900-909.
[10] Lu Y. Identification and roles of photosystem Ⅱ assembly, stability, and repair factors in Arabidopsis [J]. Frontiers in Plant Science, 2016, 7: 168. DOI: 10.3389/fpls.2016.00168.
[11] Aro E M, Virgin I, Andersson B. Photoinhibition of photosystem Ⅱ: inactivation, protein damage and turnover [J]. BBA-Bioenergetics, 1993, 1143(2): 113-134.
[12] 張子山. 低溫弱光脅迫下黃瓜葉片光系統Ⅰ與光系統Ⅱ的相互作用[D]. 泰安: 山東農業大學, 2013: 15-17.
[13] Huang W, Zhang S B, Cao K F. The different effects of chilling stress under moderate light intensity on photosystem Ⅱ compared with photosystem I and subsequent recovery in tropical tree species [J]. Photosynthesis Research, 2010, 103(3): 175-182.
[14] Takahashi S, Murata N. How do environmental stresses accelerate photoinhibition? [J] Trends in Plant Science, 2008, 13(4): 178-182.
[15] 程冬梅, 張志勇, 周賽霞, 等. 三種常綠闊葉樹光系統Ⅱ在低溫脅迫下的光抑制及恢復[J]. 廣西植物, 2018, DOI: 10.11931/guihaia.gxzw201808011.
[16] 吳道良. 紅河州不同海拔高度茶樹主要品質成分與生理學特性的變化[D]. 武漢: 華中農業大學, 2008.
[17] 施嘉璠, 謝序賓, 唐茜, 等. 茶葉光合作用強度的因素再探[J]. 四川農業大學學報, 1988, 1: 9-14.
[18] Ye Z P. A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa [J]. Photosynthetica, 2007, 45(4): 637-640.
[19] Platt T, Gallegos C L, Harrison W G. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton [J]. Journal of Marine Research, 1980, 38(4): 687-701.
[20] Togashi H F, Prentice I C, Atkin O K, et al. Thermal acclimation of leaf photosynthetic traits in an evergreen woodland, consistent with the coordination hypothesis [J]. Biogeosciences, 2018, 15(11): 3461-3474.
[21] 李存信, 林德輝. 不同海拔地區種植的水稻葉片光合作用特征的比較[J]. 云南植物研究, 1986, 8(4) :459-466.
[22] 韋玉, 李熙萌, 桑衛國, 等. 不同海拔高度矮嵩草的光合響應差異[J]. 生態科學, 2014, 33(6): 1160-1164.
[23] Woodward F. The differential temperature responses of the growth of certain plant species from different altitudes. Ⅱ. Analyses of the control and morphology of leaf extension and specific leaf area of Phleum bertolonii D. C. and P. alpinum L [J]. New Phytologist, 1979, 82(2): 397-405.
[24] Hovenden M J, Vander Schoor J K. The response of leaf morphology to irradiance depends on altitude of origin in Nothofagus cunninghamii [J]. New Phytologist, 2006, 169(2): 291-297.
[25] 張蘭, 魏吉鵬, 沈晨, 等. 秋茶光合作用與品質成分變化的分析[J]. 茶葉科學, 2018, 38(3): 271-280.
[26] 李鑫, 張麗平, 張蘭, 等. 茶園高溫干旱災害防控技術[J]. 中國茶葉, 2018(7): 38-41.
[27] 孫君, 朱留剛, 林志坤, 等. 茶樹光合作用研究進展[J]. 福建農業學報, 2015, 30(12): 1231-1237.
[28] 陶漢之. 茶樹光合生理的研究[J]. 茶葉科學, 1991, 11(2): 169-170.
[29] 陳曉晨, 徐影, 姚遙. 不同升溫閾值下中國地區極端氣候事件變化預估[J]. 大氣科學, 2015, 39(6): 1123-1135.
[30] Reich P B, Sendall K M, Stefanski A, et al. Effects of climate warming on photosynthesis in boreal tree species depend on soil moisture [J]. Nature, 2018, 562(7726): 263-267.
[31] Pepin N, Lundquist J. Temperature trends at high elevations: patterns across the globe [J]. Geophysical Research Letters, 2008, 35(14): L14701. DOI: 10.1029/2008GL034026.
[32] Rangwala I, Miller J R. Climate change in mountains: a review of elevation-dependent warming and its possible causes [J]. Climatic Change, 2012, 114(3/4): 527-547.
Different Photosynthetic Responses of Camellia sinensis to Lushan Altitude Gradient
CHENG Dongmei1, ZHANG Li1, WEI Hongfei2, JIANG Xinfeng3, ZHOU Saixia1, ZHANG Zhiyong1, PENG Yansong1*
1. Provincial Key Laboratory of Plant Ex Situ Conservation and Utilization, Lushan Botanical Garden, Jiangxi Province and Chinese Academy of Sciences, Jiujiang 332900, China; 2. Jiujiang Lushan Mount Tea Research Institute, Jiujiang 332900, China; 3. Jiangxi Sericulture and Tea Research Institute, Nanchang 330202, China
Abstract: To illustrate the photosynthetic response and photosystem Ⅱ (PSⅡ) photoinhibition of tea leaves to altitudinal gradient, native tea population of Camellia sinensis grown in the cloudy and misty Lushan Mount was measured in terms of photosynthetic gas exchange and chlorophyll fluorescence. Results show that the leaf thickness, maximum photosynthesis rate (Pnmax), maximum relative electron transport rate of PSⅡ[rETR(Ⅱ)]max, compensation irradiation (Ic), saturation irradiation (Isat), half saturation irradiation (Ik) and water usage efficiency (WUE) increased with the altitude increased. While vapor pressure deficit (VPD), stomatal conductance, transpiration rate, intercellular CO2 and dark respiration rate (Rd) displayed an opposite trend. The maximum potential photochemical efficiency of PSⅡ (Fv/Fm) increased while the quantum yield of non-regulated energy dissipation of PSⅡ[Y(NO)] decreased with the altitude increased. The effective photochemical quantum yield of PSⅡ[Φ(Ⅱ)], photochemical quenching (qP) and relative electron transport rate of PSⅡ[rETR(Ⅱ)] all increased at higher altitudes. Overall, tea trees at lower altitudes exhibited decrease of photosynthetic performance compared to those at higher altitudes, due to photoinhibition of PSⅡ.
Keywords:Lushan Yunwu tea, altitude gradient, photosynthesis, gas exchange, chlorophyll fluorescence, photoinhibition
中圖分類號:S571.1
文獻標識碼:A
文章編號:1000-369X(2019)04-447-08
收稿日期:2018-10-24
修訂日期:2019-01-06
基金項目:江西省重點研發計劃項目(20161BBF60054)、中國科學院重點部署項目(KFJ-3W-N01-143)、江西省火炬計劃(20111BBE51005)、江西省科研基礎條件和能力建設項目(2017ZDD01002,2018ZDD10002)
作者簡介:程冬梅,女,博士,助理研究員,主要從事光合作用生理生態方面的研究。*通信作者:454260156@qq.com
