耐低pH酵母菌的分離及其對酸脅迫環境的適應性
2019-08-28 12:52:44柴麗娜傅徐陽馬婷婷付大偉
食品工業科技 2019年16期
關鍵詞:酵母菌
徐 偉,柴麗娜,傅徐陽,馬婷婷,付大偉,王 薇
(1.哈爾濱商業大學食品工程學院,黑龍江哈爾濱 150076;2.黑龍江省質量監督檢測研究院,黑龍江哈爾濱 150028)
果酒的品質與釀造果酒的微生物緊密相關,優良的菌種可以提高果酒的質量,是釀造果酒的關鍵[1]。果酒發酵的過程中,酵母菌的作用是不可忽略的,其中的釀酒酵母菌(Saccharomycescerevisiae)是產酒精的主要微生物;非釀酒酵母菌則是產香氣的微生物,主要產香氣的有葡萄汁有孢漢遜酵母(Hanseniasporauvarum)、季也蒙有孢漢遜酵母(Hanseniasporaguilliermondii)、東方伊薩酵母(Issatchenkiaorientalis)、巴斯德畢赤酵母(Pichiapastoris)等。與釀酒酵母相比,非釀酒酵母具有生產分泌多種酶(酯酶、葡萄糖苷酶、蛋白酶等)的能力,可以與原料中的風味前體化合物相互作用產生芳香活性化合物,所以在發酵產香過程中發揮重要作用[2]。Moreria等[3]對葡萄汁有孢漢遜酵母,季也蒙有孢漢遜酵母和釀酒酵母的純種發酵及混合發酵進行了研究,其中發現季也蒙有孢漢遜酵母具有最高的生長力,同時2-乙酸苯乙酯和2-苯基乙醇的產生量也很高,并且受其它菌的影響也很小,而這些物質可賦于果酒特有的果香與花香[4]。一般而言,工業化生產相比于傳統化生產的酵母菌多樣性明顯減少。為更好的體現果酒自身的典型性和風格,且保留特定生態條件下果酒香氣特征的物質,增加果酒中酵母菌種的多樣性是必不可少的[5-6]。
耐酸酵母菌是指可以在pH≤4的條件下生長或代謝的一類酵母菌,當發酵液的pH控制在3.5~4.5左右時,細菌及其他絕大多數雜菌均受到抑制,而果酒酵母則能正常活動[7]。據報道[8],酸脅迫過程會引起氧化脅迫,抗氧化酶的活性發生了改變,生物體內應對氧化脅迫最重要的兩類酶分別是超氧化物歧化酶(Superoxide dismutase,SOD)和過氧化氫酶(Hydrogen Peroxidase,CAT)。活性氧是需氧生物細胞代謝的正常產物,但環境脅迫易導致活性氧簇(ROS)過量產生,使得細胞發生氧化脅迫。為了消除不利影響,生物體需要依靠過氧化氫酶和超氧化物歧化酶等抗氧化酶共同作用將ROS降解為無害物質[9]。因此,本文在酸環境下分離篩選耐低pH酵母菌,并采用掃描電子顯微鏡(SEM)和透射電子顯微鏡(TEM)觀察菌體形態以及細胞結構的變化,對耐低pH酵母菌中抗氧化關鍵酶CAT和SOD的酶活力進行研究,為酵母菌的耐酸機理提供理論依據,為特色漿果果酒的開發提供新的菌株。
1 材料與方法
1.1 材料與儀器
紅樹莓、藍莓、藍靛果 帽兒山果園采摘;對照菌1 葡萄汁有孢漢遜酵母(Hanseniasporauvarum)CICC 32337購買于中國工業微生物菌種保藏管理中心;對照菌2果酒酵母菌 市售菌粉;酵母浸粉、蛋白胨、瓊脂、麥芽浸粉等均為生物試劑 北京奧博星生物技術有限責任公司;鉬酸銨、磷酸二氫鉀(鈉)、磷酸氫二鉀(鈉) 天津博迪化工股份有限公司;30%過氧化氫 天津市天力化學試劑有限公司;EDTA、鄰苯三酚 天津市光復精細化工研究所。
JEM-2100F型日立透射電子顯微鏡、SU8010型日立冷場發射掃描電鏡 HITACHI公司;pHS-3C型雷磁pH計 廣州市深華生物技術有限公司;HWS24型電熱恒溫水槽 上海一恒科技有限公司;H2050R型離心機 湖南湘儀實驗室儀器開發有限公司;LG10-2.4A型高速離心機 北京德世科技有限公司;723N UV5100B型紫外可見分光光度計 上海元析儀器有限公司。
1.2 耐低pH酵母菌的分離篩選及鑒定
1.2.1 耐低pH酵母菌的分離篩選 吸取自然發酵7 d內的紅樹莓、藍靛果果汁以及藍莓果汁各1 mL,進行10-1~10-7梯度稀釋,選取濃度10-4、10-5、10-6在麥芽汁培養基中進行三區劃線初篩,并在YPD瓊脂培養基中劃線3次以達到純化,將所得純菌株進行劃線標號并保存。
將分離篩選到的菌株采用WL培養基進行初步鑒定,挑選菌落差異大的菌株接種到pH2.3、2.6、2.9、3.2、3.5、3.8、4.1、4.4 YPD液體培養基中,依據酒精度、產酯量、CO2生成量等發酵性能,篩選出的酵母菌再進行生理生化鑒定和分子生物學鑒定[10-11]。
1.2.2 生理生化鑒定 主要從脲酶試驗、產酯試驗、DBB試驗、產類淀粉試驗、碳源同化試驗、氮源同化試驗、糖發酵試驗及產酸試驗等對菌株進行生理生化鑒定,具體方法參照酵母菌的特征與鑒定手冊[12-13]。
將所得序列在GenBank數據庫中與已知同源序列進行比較(Blast)。下載相關菌種序列,用MEGA軟件進行相似性分析,并通過該軟件的鄰近法(neighbor joining)構建系統發育樹[14]。
1.3 耐低pH酵母菌對酸脅迫環境適應性的研究
1.3.1 耐低pH酵母菌的細胞形態與結構的電鏡觀察 J-23菌株和對照菌1、L-6菌株和對照菌2分別接種于pH2.3和4.8 YPD液體培養基各100 mL,置于28 ℃恒溫培養箱中培養2 d,以5000 r/min離心15 min,取菌泥至2 mL EP管中的0.1 mL處,加入固定液于4 ℃冰箱固定待用,再經過沖洗、脫水、置換、干燥、粘樣、鍍膜等一系列處理即可得到用于掃描電鏡檢測樣[15]。
透射電鏡的檢測樣經過固定、脫水、浸透、包埋、聚合、修塊、超薄切片機切片、醋酸鈾-枸櫞酸鉛雙染色、透射電鏡觀察,拍片[16]。
1.3.2 不同pH對酵母菌的抗氧化酶活力的影響
1.3.2.1 不同pH對酵母菌SOD的酶活力影響 SOD的提取:采用甲苯法提取SOD粗酶液,發酵液在5000 r/min,15 min條件下離心,取菌泥,每克菌泥加入0.924 mL甲苯,35 ℃,水浴45 min;加入3 mL pH7.8磷酸緩沖液(含0.1 mmol/L EDTA)提取5 h,使用高速離心機離心,取上清液(即粗酶液)。粗酶液50 ℃預熱30 min,離心取上清,加2 mol/L HCl調pH至5.0,4000 r/min離心15 min,取上清液,加入0.6倍的冷丙酮,攪拌,離心,取上清液,再加入0.6倍冷丙酮,離心,留沉淀,即SOD純產品[17]。
SOD酶活測定:以終止劑改進的鄰苯三酚自氧化法,紫外分光光度計420 nm法自氧化速率的測定。9 mL 50 mmol/L Tris-HCl(pH8.2,含1 mmol/L二乙撐三胺五乙酸),25 ℃水浴20 min,再加入40 μL在25 ℃預熱的45 mmol/L鄰苯三酚溶液(10 mmol/L HCl配制),混勻,每隔半分鐘測一次A0,反應3 min,對照組為Tris-HCl,自氧化速率應控制在0.06 A/min;加入一定量的SOD純產品和9 mL 50 mmol/L Tris-HCl(pH8.2,含1 mmol/L二乙撐三胺五乙酸),再按照相同后續步驟進行操作,每隔30 s測一次As[18-19]。SOD酶活力計算方法:
清除建筑垃圾的活,我們干得驚心動魄。那些新建成的樓房是多層,不是高層,沒有電梯,我們沿著樓梯爬行。樓梯都沒來得及裝護欄,站在頂層往下看,樓梯間的縫隙像刀斧劈出的無底深淵,我看得兩股顫顫,雙腿松軟。還好,樓梯只是我們進到每個樓層的通道,那些垃圾,不用我們運載,我們把它們裝入蛇皮袋,扛到西北角一個窗口處。窗外有一只吊籃。我們將垃圾搬到吊籃上。吊籃有專人操作,上上下下,起降匆忙。
式中:V表示反應液體積,mL;V1表示加樣體積,mL;N表示樣品稀釋倍數。
1.3.2.2 不同pH對酵母菌CAT的酶活影響 CAT的提取:吸取1 mL酵母菌液,加入9 mL 0.155 mol/L的KCl溶液于錐形瓶中,充分搖勻,用研缽在冰浴中研5 min,在4 ℃、8000 r/min下將勻漿液離心13 min,吸取上清液,最后用0.85%生理鹽水稀釋10倍,制得粗酶液。
CAT酶活測定:可見光光度法測定:取6支20 mL試管,如表1的加樣量于試管中,記錄J-23、L-6菌株及2株對照菌在405 nm處的吸光度,計算CAT酶活力[20]。酶活力計算方法:

表1 測定CAT酶活所需試劑的加樣量
式中:A測表示測定管中所加試劑在405 nm處的吸光度;A對表示對照管中所加試劑在405 nm處的吸光度;A標表示標準管中所加試劑在405 nm處的吸光度。
1.4 數據處理
酶活力的測定中每個樣品設3個平行,數據處理采用OriginLab Origin 9.0和SPSS Statistics 17.0軟件進行分析。
2 結果與分析
2.1 耐低pH酵母菌的分離篩選及鑒定結果
2.1.1 耐低pH酵母菌的分離篩選結果 從3種果酒中共得到75株酵母菌菌株,顯微鏡下觀察菌體形態有卵形、圓形、橢圓形等;在YPD瓊脂培養基上的菌落形態為奶白色、突起、表面光滑、濕潤、無褶皺。通過WL培養基初步鑒定篩選到8株菌落差異明顯的菌株,并在8個不同pH下測定發酵性能,篩選出生長狀況最好的J-23、L-6菌株為后續實驗菌株。
2.1.2 生理生化鑒定結果 J-23和L-6菌株的生理生化結果見表2。
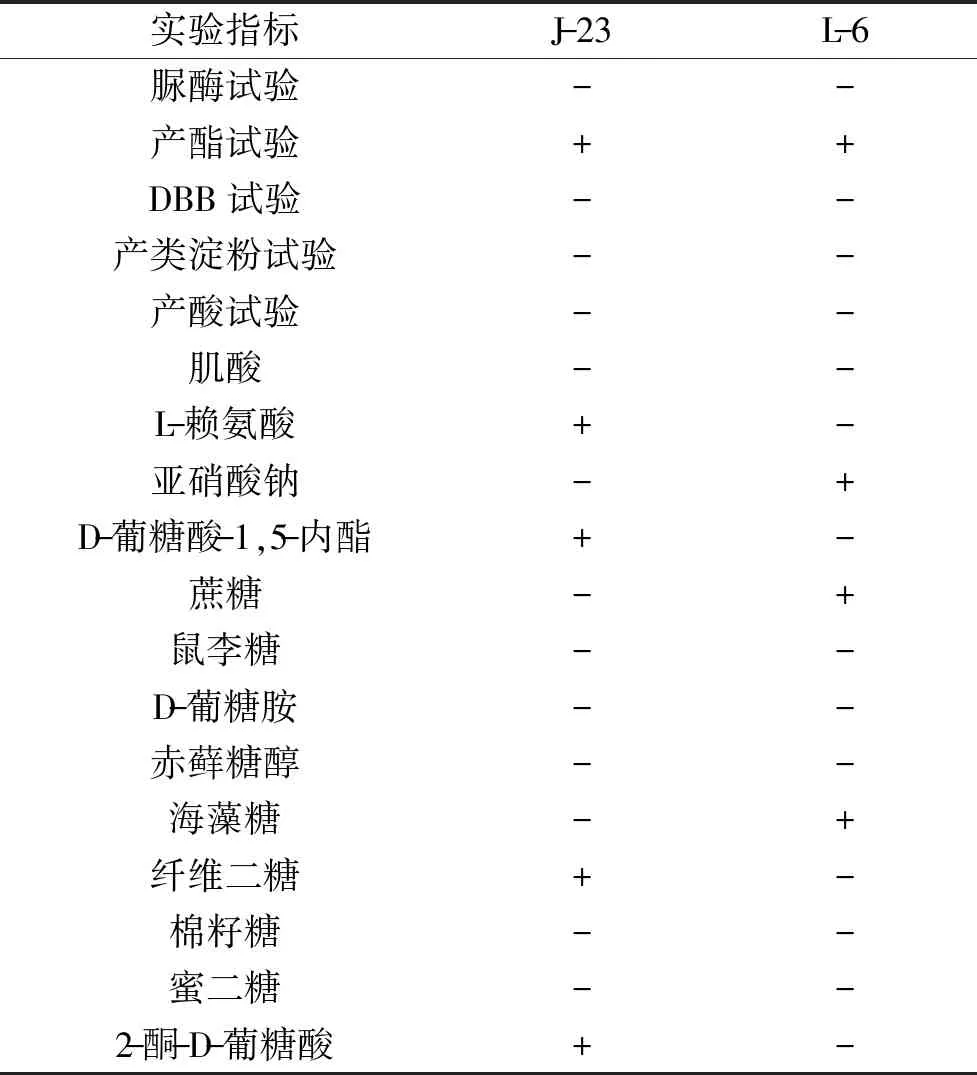
表2 2株菌株生理生化鑒定結果
由表2可知,通過WL培養基初步鑒定菌落特征:J-23菌株中間為淡綠色,邊緣帶白色;L-6菌株為藍綠色,邊緣帶白色。再結合生理生化特征,初步鑒定J-23菌株為季也蒙有孢漢遜酵母菌,L-6菌株為釀酒酵母菌。
2.1.3 分子生物學鑒定結果 通過在BLAST網站中的GenBank核酸序列數據庫中對獲得的序列進行同源序列搜索和相關信息檢索,將分離得到的菌株與模式菌株使用ClustalX 1.83排序后,并使用MEGA three 3.1計算序列相似性。通過Neighbor-Joining分析方法生成系統發育樹,并進了1000次bootstrap的統計檢驗[21],結果如圖1、圖2。
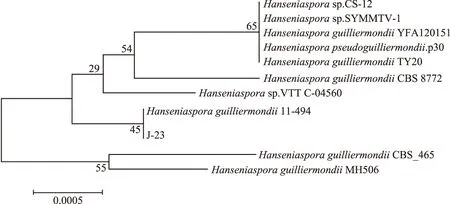
圖1 菌株J-23的系統發育樹關系圖

圖2 菌株L-6的系統發育樹關系圖
通過構建J-23菌株與10株模式菌株26S rDNA Dl/D2區域序列,發現J-23菌株與模式菌株Hanseniasporaguilliermondii11-494聚為一枝,同源性最近支持率為100%,結合兩者D1/D2序列一致性為100%,鑒定J-23菌株為季也蒙有孢漢遜酵母(Hanseniasporaguilliermondii);構建L-6菌株與11株模式菌株26S rDNA Dl/D2區域序列,與模式菌株SaccharomycescerevisiaeYLL20聚為一枝,同源性最近支持率為99%,結合兩者D1/D2序列一致性為99%,鑒定L-6菌株為釀酒酵母(Saccharomycescerevisiae)。
2.2 耐低pH酵母菌對酸脅迫環境適應性的研究結果
2.2.1 耐低pH酵母菌細胞形態與結構的變化結果
2.2.1.1 掃描電鏡對耐低pH酵母菌細胞形態的觀察結果 選擇購買的對照菌1、對照菌2與分離出來的J-23、L-6菌株分別在pH2.3和4.8的YPD培養基中培養,在SEM下觀察它們的細胞形態變化,結果如圖3所示。

圖3 不同pH下各菌株掃描電鏡圖(10k×)
由圖3可見,J-23菌株和對照菌1的菌體為橢圓形,L-6菌株和對照菌2的菌體為圓形。
在pH4.8時,J-23菌株相比于對照菌1的細胞表面光滑完整,菌體比較飽滿,都有明顯的芽痕;在pH2.3時,J-23的菌體外貌大致保持不變,但對照菌1出現明顯的凹陷、表面破裂和褶皺,細胞形態出現嚴重的變形。在pH4.8時,L-6菌株相比于對照菌2的細胞表面光滑完整,菌體都比較飽滿,芽痕較多;在pH2.3時,L-6菌株的菌體外貌差異性較小,對照菌2與其相比,菌體出現了明顯的褶皺和破皮現象。
2.2.1.2 透射電鏡對低pH酵母菌細胞結構的觀察結果 選擇購買的對照菌1、對照菌2與分離出來的J-23、L-6菌株分別在pH2.3和4.8的YPD培養基中培養,在TEM下觀察它們的細胞結構變化,結果如圖4所示。
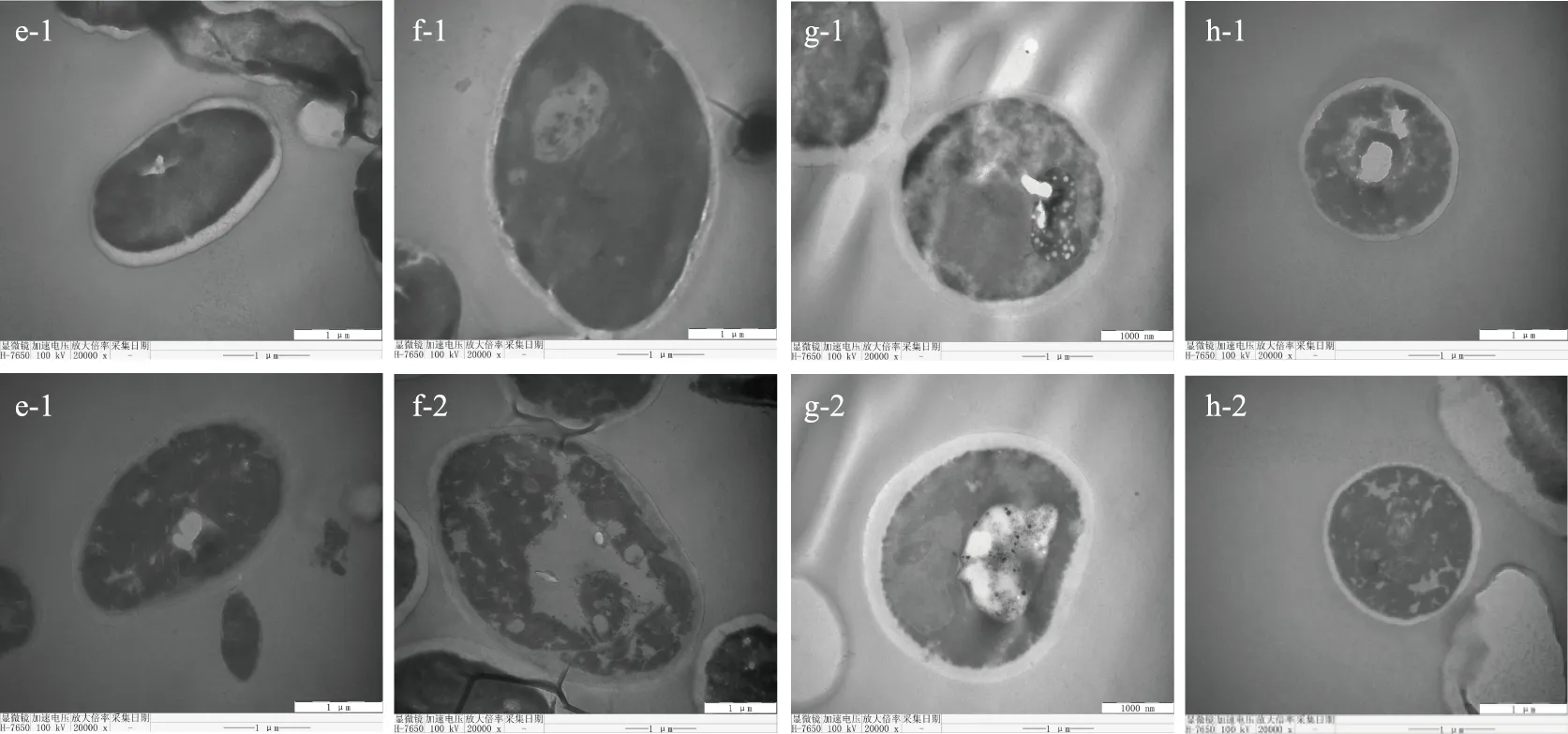
圖4 不同pH下各菌株透射電鏡圖(20k×)
由圖4可見,在pH4.8時,J-23、L-6菌株及2株對照菌的細胞核、細胞膜、細胞壁、液泡等細胞結構都比較清晰,其中液泡最大且飽滿,可以貯藏和消化細胞內的一些代謝產物,維持細胞的形狀,起支持作用;在pH2.3時,J-23菌株的液泡變大,貯藏粒也增多,可以貯藏更多的酶類物質,調節滲透壓,協助維持細胞的形狀;而對照菌1的細胞壁在酸環境下被破損,導致細胞壁結構不清晰。L-6菌株的細胞結構整體變化不大,而液泡變大,自身可通過調節滲透壓來維持完整性;對照菌2的細胞形狀出現明顯的變形,細胞壁、細胞膜向內凹陷,可能是因為酸環境使某些胞內酶失活,滲透性降低,引起細胞內部脫水。
2.3 不同pH對酵母菌的抗氧化酶活力的影響結果


表3 不同pH下酵母菌中主要抗氧化酶活力的變化結果
2.3.2 不同pH對酵母菌CAT酶活力的影響結果 過氧化氫酶催化H2O2分解為H2O和O2,以清除體內過多的H2O2,從而抵制了氧化脅迫,進而減輕了弱有機酸的產生和酸環境引起的酸脅迫現象。由表3可知,在pH2.3與pH4.8時,J-23和L-6菌株中的CAT酶活力均顯著高于對照菌1和對照菌2。在低pH下,耐低pH酵母菌中CAT酶活力的增加,進一步體現了J-23和L-6菌株的耐酸性。
3 結論
本文從自然發酵的藍靛果與紅樹莓果汁中,分離篩選出2株耐低pH酵母菌,經鑒定L-6菌株為釀酒酵母菌;J-23菌株為季也蒙有孢漢遜酵母菌。SEM觀察pH2.3與pH4.8時,L-6和J-23菌體形態均呈現出:表面光滑完整、飽滿,未出現明顯的凹陷、褶皺等現象;在TEM下觀察結果,細胞結構完整清晰,為維持低pH環境細胞的形狀不變,可見胞內液泡增大。同時,L-6和J-23菌株抗氧化酶SOD和CAT酶活力均顯著(p<0.05)高于普通對照菌株,表明所分離的2株酵母菌,能夠很好地適應酸脅迫的環境,為高酸果汁發酵提供優良菌種。
猜你喜歡
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
中國民間療法(2021年8期)2021-07-22 05:53:28
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
當代水產(2019年3期)2019-05-14 05:43:24
天然產物研究與開發(2018年9期)2018-10-08 03:25:32
煙草科技(2015年8期)2015-12-20 08:27:04
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
食品工業科技(2014年9期)2014-03-11 18:15:31
