阿爾茨海默病相關差異表達基因及其生物信息學分析
2019-09-10 07:22:44徐倩蘇湲淇譚毅楊元娟
中國藥房 2019年24期
徐倩 蘇湲淇 譚毅 楊元娟
摘 要 目的:為阿爾茨海默病(AD)發病機制的闡釋、早期預防與診斷以及治療靶點的篩選提供參考。方法:從美國國立生物技術信息中心公共數據平臺基因表達數據庫中下載基因芯片數據集GSE28146,使用GEO2R在線分析工具篩選出AD相關差異表達基因(DEGs);使用DAVID 6.8生物信息學資源數據庫進行基因本體(GO)分析和KEGG通路富集分析;使用STRING數據庫和Cytoscape 3.2.1軟件進行蛋白-蛋白相互作用(PPI)網絡分析。結果與結論:篩選出AD相關DEGs共1 478個,其中上調913個、下調565個。GO分析結果顯示,DEGs主要分布于細胞質、膜、細胞外隙中,主要通過轉錄的正/負調節、核因子κB活性的正調節、Rho蛋白信號轉導的調節、蛋白質磷酸化的調節等生物學過程以及蛋白質結合、DNA結合、轉錄因子活性(序列特異性DNA結合)等分子功能來誘導AD的發生。KEGG通路富集分析結果顯示,DEGs顯著富集于癌癥途徑、肺結核、破骨細胞分化、Janus激酶/信號傳導及轉錄激活因子信號通路、叉頭轉錄因子信號通路、EB病毒感染等信號通路上。DEGs編碼蛋白的PPI網絡含節點蛋白共1 205個、邊3 931條;其中的關鍵核心基因為SOCS3、NEDD4、CBLB,可能是AD發生發展的潛在靶點。
關鍵詞 阿爾茨海默病;差異表達基因;生物信息學;基因本體;KEGG通路富集;蛋白-蛋白相互作用
ABSTRACT? ?OBJECTIVE: To provide reference for interpretation of pathogenesis, early prevention and diagnosis, and selection of therapeutic targets of Alzheimer’s disease (AD). METHODS: The gene chip dataset GSE28146 was downloaded from the NCBI public data platform GEO, and the AD-related differentially expressed genes (DEGs) were identified by using GEO2R online analysis tool. GO analysis and KEGG enrichment pathway analysis were performed by using DAVID 6.8 bioinformatics resource database. The protein-protein interaction (PPI) network analysis was performed by using STRING database and Cytoscape 3.2.1 software. RESULTS & CONCLUSIONS: A total of 1 478 AD-related DEGs were identified, consisting of 913 up-regulated genes and 565 down-regulated genes. GO function enrichment analysis showed that DEGs mainly distributed in cytoplasm, membrane, extracellular space, and induced AD via biological processes such as positive/negative regulation of transcription, positive regulation of NF-κB activity, regulation of Rho protein signaling transduction, protein phosphorylation; via protein binding, DNA binding, transcription factor activity (sequence specific DNA binding) and other molecular functions. KEGG pathway enrichment analysis showed that DEGs was enriched in cancer pathway, pulmonary tuberculosis, osteoclast differentiation, JAK/STAT signaling pathway, FoxO signaling pathway, EB virus infection and other signaling pathways. There are 1 205 nodes and 3 931 edges in the PPI network of DEGs coding protein. Among them, the key genes are SOCS3, NEDD4 and CBLB, which may be the potential target of AD development.
KEYWORDS? ?Alzheimer’s disease;Differentially expressed genes;Bioinformatics; Gene ontology; KEGG pathway enrichment; Protein-protein interaction
阿爾茨海默病(Alzheimer’s disease,AD)是一種慢性進行性的中樞神經系統退行性疾病,臨床上主要表現為記憶障礙、進行性認知功能損害、人格和行為改變等神經精神障礙[1]。AD的發病機制較為復雜,呈現多樣性和不確定性,以β-淀粉樣蛋白(Aβ)瀑布理論、Tau蛋白學說、氧化應激、炎癥機制、線粒體功能障礙和基因突變等假說較為常見[2]。AD是老年癡呆最常見的病因,據統計,60%~80%的癡呆由AD導致,且癡呆已經成為65歲以上人群的第五大死亡原因[3]。近年來,隨著人口老齡化進程加劇,我國AD患者的數量迅速增加,現已超過700萬人[4]。AD病程長、并發癥多樣且需要長期護理,給患者家庭和社會帶來沉重的經濟負擔,已成為不容忽視的社會問題[4]。隨著高通量測序技術及基因芯片技術的發展,從諸如基因組、轉錄組、蛋白質組和代謝組等不同層次研究疾病的發生、發展及預后已成為全世界學者共同關注的熱點之一[5-7]。本研究通過對美國國立生物技術信息中心(NCBI)公共數據平臺中的表達譜芯片原始數據進行挖掘,篩選出AD相關差異表達基因(DEGs),并對其進行基因本體(GO)分析、KEGG通路富集分析以及蛋白-蛋白相互作用(PPI)網絡分析,以期為AD發病機制的闡釋、早期預防與診斷以及治療靶點的篩選提供理論參考。
1 資料與方法
1.1 資料來源
從NCBI公共數據平臺基因表達數據庫(GEO,http://www.ncbi.nlm.nih.gov/geo)下載AD相關基因芯片數據集GSE28146(https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE28146)。該數據集基于GPL570平臺采用[HG-U133_Plus_2]Human Genome U133 Plus 2.0 Array陣列芯片(美國Affymetrix公司)檢測的30例人海馬組織獲得,其中對照組8例、AD病例組22例。
1.2 方法
運用GEO2R在線分析工具(https://www.ncbi.nlm.nih.gov/geo/geo2r/?acc=GSE28146)對芯片原始數據進行DEGs的篩選,篩選條件為P<0.05,其中log2FC≥1.0為上調、log2FC<-1.0為下調(式中,“FC”表示AD病例組受試芯片熒光信號強度與對照組相比的差異倍數)[8]。采用GraphPad Prism 5在線軟件(https://www.graphpad.com/scientific-software/prism)繪制DEGs火山圖。
利用DAVID 6.8生物信息學資源數據庫(https://david.ncifcrf.gov/)對DEGs進行GO分析和KEGG通路富集分析(以Fisher確切概率法計算P值,P<0.05為“顯著富集”)[9]。將P值由小到大排序,分列出排序前10位的GO功能族和KEGG信號通路。
通過STRING數據庫(https://www.string-db.org/)對篩選所得DEGs進行PPI網絡分析,設置“可信度(Confidence)”為0.7;借助Cytoscape 3.2.1軟件的“CytoHubba”插件對DEGs編碼蛋白的相互作用進行可視化展示。其中,節點表示蛋白,邊表示蛋白之間的相互聯系,節點度值表示與某節點相連邊的數量(其值大小與對應節點在網絡中的重要程度成正比),將節點度值排序前3位的基因視為關鍵核心基因。
2 結果
2.1 DEGs的篩選結果
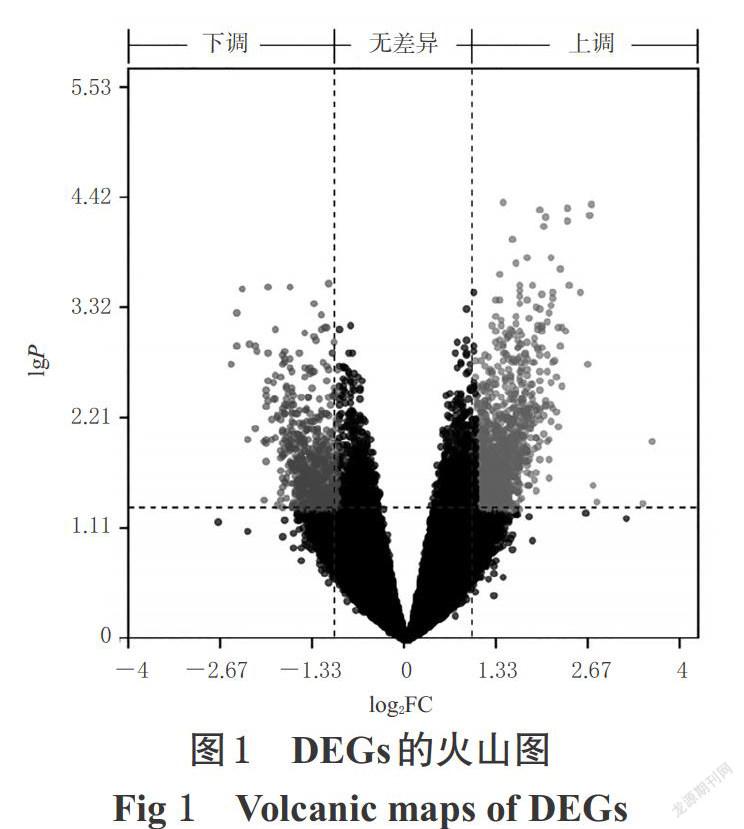
篩選出AD相關DEGs共1 478個,其中上調913個、下調565個。DEGs的火山圖見圖1。
2.2 DEGs的GO分析及KEGG通路富集分析結果
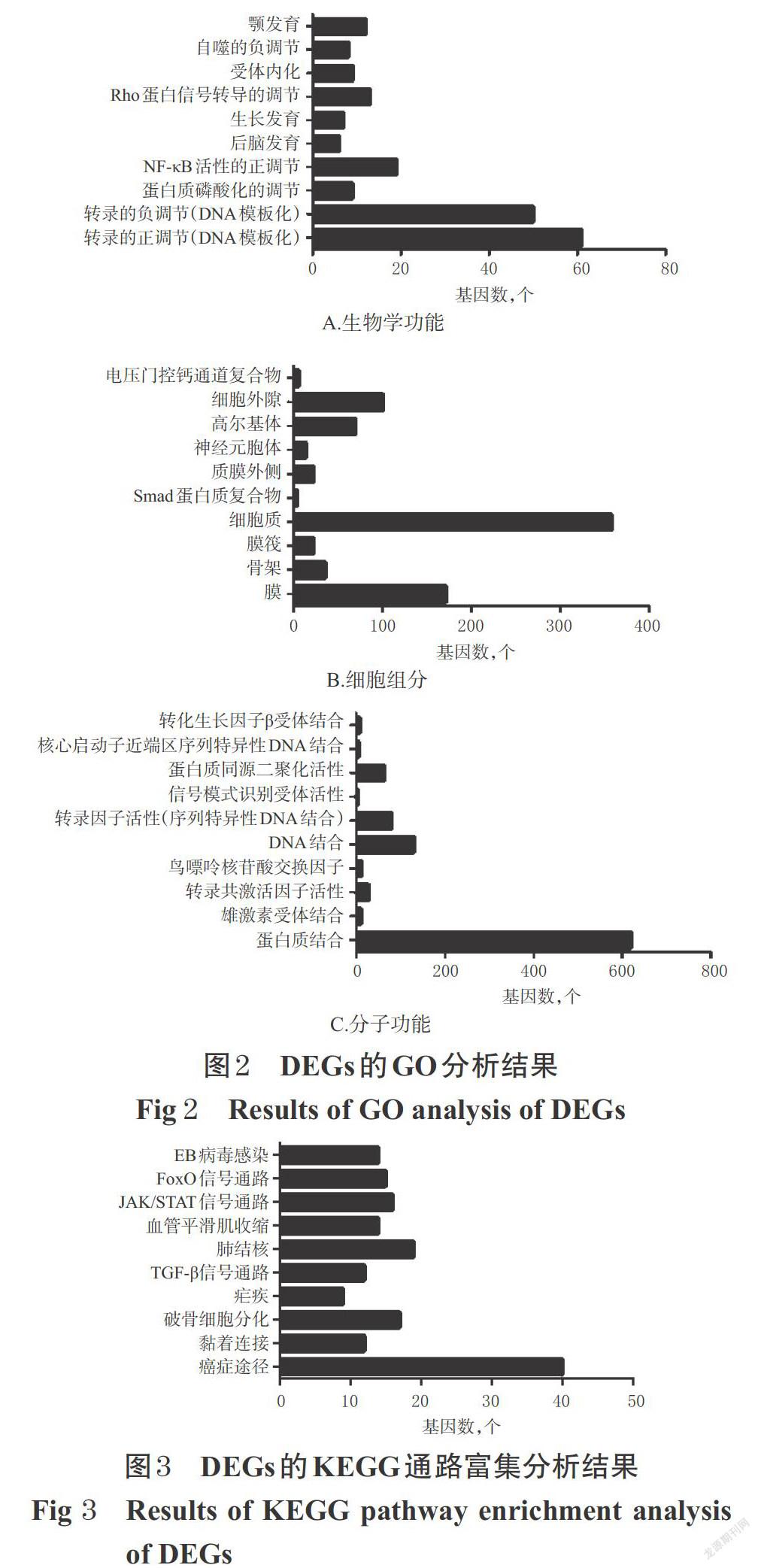
GO分析結果顯示,DEGs主要涉及轉錄的正/負調節、核因子κB(NF-κB)活性的正調節、Rho蛋白信號轉導的調節、顎發育、蛋白質磷酸化的調節、自噬的負調節等生物學功能;DEGs主要分布于細胞質、膜、細胞外隙、高爾基體等細胞組分;DEGs主要涉及蛋白質結合、DNA結合、轉錄因子活性(序列特異性DNA結合)、蛋白質同源二聚體活性等分子功能,詳見圖2。
KEGG通路富集分析結果顯示,DEGs在癌癥途徑、肺結核、破骨細胞分化、Janus激酶/信號傳導及轉錄激活因子(JAK/STAT)信號通路、叉頭轉錄因子(FoxO)信號通路、EB(Epstein-Barr)病毒感染、轉化生長因子β(TGF-β)等信號通路上顯著富集,詳見圖3。
2.3 DEGs編碼蛋白PPI網絡分析結果
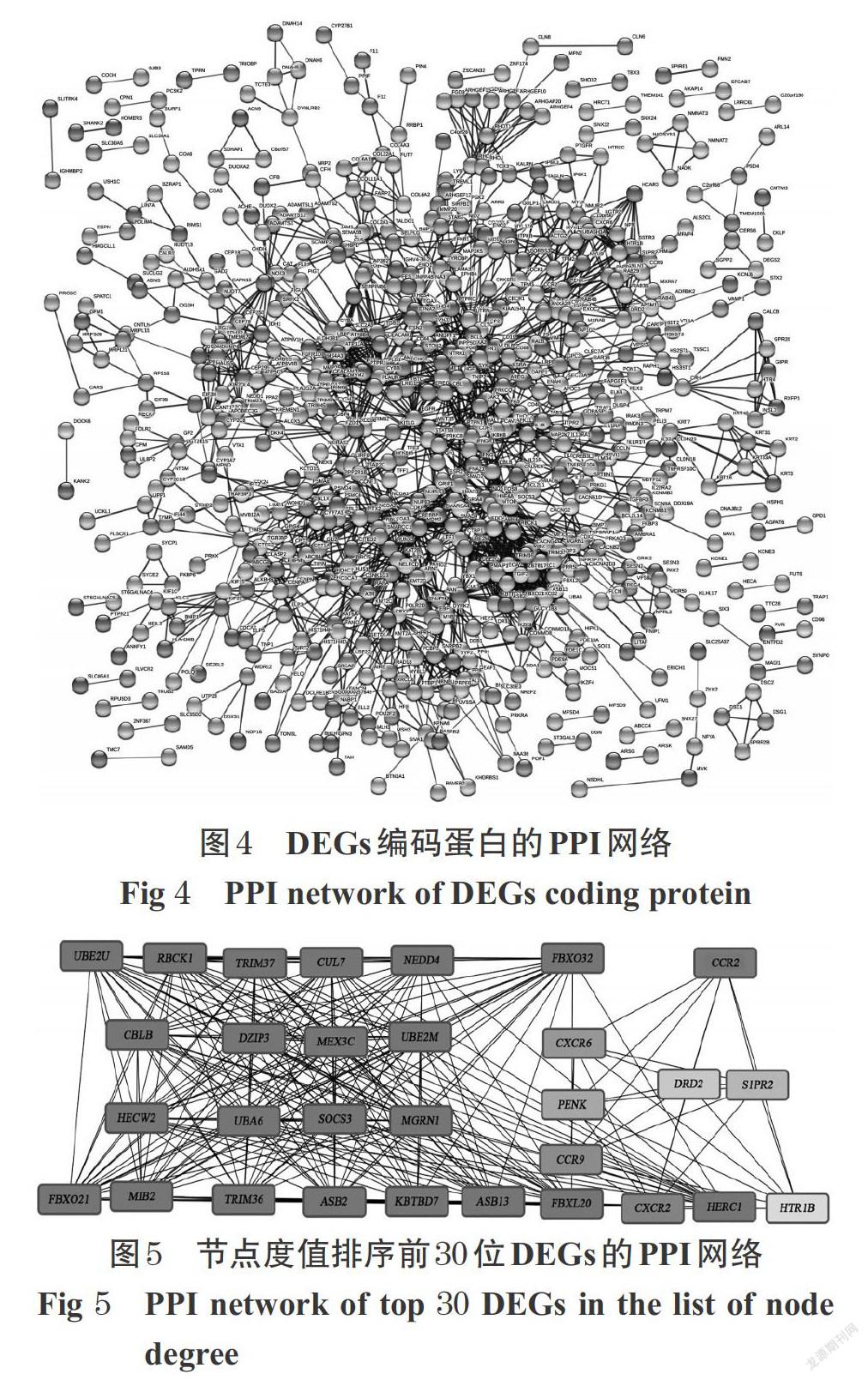
DEGs編碼蛋白的PPI網絡中,共包含節點蛋白1 205個、邊3 931條,見圖4。其中,節點度值排序前30位蛋白的PPI網絡見圖5(以基因表示),節點度值排序前3位的基因為SOCS3、NEDD4和CBLB,是網絡中的關鍵核心基因。
3 討論
近年來,隨著生物芯片、高通量測序等現代生物技術的高速發展和生物信息學分析的日益成熟,通過大數據分析,挖掘在疾病發生發展中起主導作用的相關基因,可為疾病發病機制和治療評價等研究提供新的思路。例如曹丹等[10]采用生物信息學方法分析肝癌相關DEGs,并構建編碼蛋白PPI網絡,發現TOP2A基因可能是肝癌相關的核心基因;馮曉飛等[11]利用生物信息學方法對骨肉瘤基因表達譜芯片進行分析,從分子水平上初步分析了骨肉瘤的潛在發病機制;許丁文等[12]應用生物信息學方法分析了絲甘蛋白聚糖對卵巢癌耐藥性的影響及作用機制。
AD作為癡呆的主要原因,是目前全球醫療保健領域最嚴峻的挑戰之一[13]。AD致病機制尚未明確,其發病是多基因、多途徑、多步驟、多階段相互作用和相互影響的復雜過程,尚缺乏有效的診療手段,對其早發現、早治療成為了臨床亟待解決的難題[1-4]。為此,本研究應用生物信息學方法,擬初步分析AD發生的潛在分子機制,以期為進一步揭示該癥病因提供新的線索,同時為AD治療新靶點的尋找提供理論依據。
本研究通過生物信息學方法從GEO數據庫中搜索得到芯片數據集GSE28146(人腦海馬組織,其中健康人腦海馬樣本8例,AD患者海馬樣本22例),并對其進行數據挖掘,包括GO分析、KEGG通路富集分析和PPI網絡分析。利用GEO2R在線分析工具對該芯片數據進行分析,共篩選出DEGs 1 478個,其中上調913個、下調565個。GO分析結果顯示,DEGs主要分布于細胞質、膜、細胞外隙中,主要通過轉錄的正/負調節、NF-κB活性的正調節、Rho蛋白信號轉導的調節、蛋白質磷酸化的調節等生物學功能以及蛋白質結合、DNA結合、轉錄因子活性(序列特異性DNA結合)等分子功能來誘導AD的發生。KEGG通路富集分析結果顯示,DEGs顯著富集于癌癥途徑、肺結核、破骨細胞分化、JAK/STAT信號通路、FoxO信號通路、EB病毒感染等信號通路上。已有研究證實,蛋白質磷酸化[2]、轉錄調節[14]、NF-κB活性[15]、Rho蛋白信號轉導[16]、線粒體自噬[17]等在AD的發生發展中具有重要作用,且TGF-β信號通路[18]、JAK/STAT信號通路[19]、FoxO信號通路[20]及EB病毒感染[21]等信號通路均參與了AD的發生發展,與本文結果基本一致。
DEGs編碼蛋白PPI網絡共包含節點蛋白1 205個、邊3 931條,其中的關鍵核心基因為SOCS3、NEDD4、CBLB。
SOCS3基因編碼細胞因子信號轉導抑制因子3(SOCS3),該蛋白在不同組織、細胞中均廣泛表達,是JAK/STAT信號通路的主要負調控因子家族成員之一[22-23]。SOCS3基因位于人染色體17q25.3,由675個核苷酸組成,為抑癌基因之一[24]。Deng J等[25]研究表明,胃癌組織中SOCS3蛋白及其mRNA的表達均較癌旁正常組織明顯下降,其表達下調及淋巴結轉移是胃癌患者預后的獨立危險因素;Jiang BG等[26]研究結果顯示,肝癌組織中SOCS3基因DNA甲基化陽性率較高,可導致該基因表達下調,與肝癌細胞的分化、侵襲和轉移密切相關,但目前尚未見該基因與AD發生的相關報道。本研究發現,SOCS3基因主要富集于調節蛋白質磷酸化功能族(GO0001932)。既往Tau蛋白學說指出,Tau蛋白的磷酸化在AD發生發展中具有重要意義[2]。由此推斷,該基因可能與AD的Tau蛋白學說有關,但有待基礎研究予以驗證。
發育抑制蛋白4(NEDD4)是一種重要的HECT型泛素連接酶,由NEDD4基因編碼,在多種神經退行性疾病相關組織中呈高表達[27]。NEDD4基因在機體發育和疾病進展過程中具有重要的生物學功能,并可結合多種底物、調節多種信號通路。有研究發現,NEDD4酶可通過對成纖維細胞生長因子受體1(FGFR1)的泛素化來調控神經和胚胎發育[28]。此外,在細胞增殖、分化、凋亡中起重要作用的第10號染色體缺失的磷酸酶及張力蛋白同源物基因(PTEN)為NEDD4酶的底物分子,后者可通過結合生長因子受體結合蛋白10(Grb10)來負調控胰島素樣生長因子1(IGF1)通路,最終發揮對細胞增殖、分化、遷移的調節作用[29-30]。同時有研究還發現,NEDD4泛素化降解絲裂原誘導因子6(Mig6)與腫瘤細胞遷移有關[31]。此外,NEDD4可調控獲得性免疫,可通過降解泛素連接酶E3來促進T細胞的激活[32];還可通過刺激B細胞中的腫瘤壞死因子受體相關因子3(TRAF3)發生賴氨酸63(K63)形式的泛素化來調節CD40介導的蛋白激酶B(Akt)通路的激活,從而參與免疫球蛋白的轉換[33]。上述研究表明,NEDD4酶可通過泛素化調控和獲得性免疫調節來參與多種神經退行性疾病的發生發展,可作為研究AD發病機制的潛在靶點之一。本研究結果顯示,NEDD4基因主要富集于對鈣的反應功能族(GO0031623),但該基因如何調節鈣反應功能族及此功能族在AD發生發展中的作用尚有待進一步研究。
CBLB基因編碼的泛素連接酶E3(CBLB)屬于環指(RING)型泛素連接酶,由氮端保守的酪氨酸激酶(TKB)結構域、RING結構域以及其他蛋白-蛋白結合模體組成。該酶中的TBK結構域可識別并結合底物蛋白酪氨酸激酶,RING結構域可募集泛素結合酶E2,從而啟動底物泛素化降解,參與細胞內信號轉導的負向調控[34]。同時,CBLB酶也可作為連接蛋白的招募信號分子,從而激活或抑制下游的信號通路分子,發揮促進細胞增殖、轉移和抗凋亡等作用[35-36]。本研究結果發現,CBLB基因主要富集于信號轉導功能族(GO0007165),在生物學功能上富集于蛋白質結合相關功能族。但筆者尚未發現該基因與AD發生的相關研究,其與AD的相關性有待確證。
綜上所述,本研究基于生物信息學方法篩選出了AD相關DEGs,并分析了其GO功能以KEGG富集通路,最終篩選出了SOCS3、NEDD4、CBLB等3個關鍵核心基因,可作為AD發病機制研究的潛在靶點,在一定程度上為深入揭示AD的分子機制和治療靶點提供了研究方向和理論依據。在后續研究中,本課題組將進一步針對我國AD人群進行分子生物學基礎研究,對上述結論進行確證。
參考文獻
[ 1 ] DOS SANTOS PICAN?O LC,OZELA PF,DE FáTIMA DE BRITO BRITO M,et al. Alzheimer’s disease:a review from the pathophysiology to diagnosis,new perspectives for pharmacological treatment[J]. Curr Med Chem,2018,25(26):3141-3159.
[ 2 ] SERRANO-POZO A,FROSCH MP,MASLIAH E,et al. Neuropathological alterations in Alzheimer disease[J]. Cold Spring Harb Perspect Med,2011. DOI:10.1101/cshperspect.a006189.
[ 3 ] GBD 2016 Dementia Collaborators. Global,regional,and national burden of Alzheimer’s disease and other dementias,1990-2016:a systematic analysis for the global burden of disease study 2016[J]. Lancet Neurol,2019,18(1):88-106.
[ 4 ] JIA J,WEI C,CHEN S,et al. The cost of Alzheimer’s di- sease in China and re-estimation of costs worldwide[J].? Alzheimers Dement,2018,14(4):483-491.
[ 5 ] ZHANG L,NIE Q,SU Y,et al. MicroRNA profile analysis on duck feather follicle and skin with high-throughput sequencing technology[J]. Gene,2013,519(1):77-81.
[ 6 ] YONGFENG H,FAN Y,JIE D,et al. Direct pathogen detection from swab samples using a new high-through put sequencing technology[J]. Clin Microbiol Infect,2011,17(2):241-244.
[ 7 ] HE X,LI X,GUO Y,et al. Newborn screening of genetic mutations in common deafness genes with bloodspot based gene chip array[J]. Am J Audiol,2018,27(1):57-66.
[ 8 ] 張東亮,趙舒煊,向高,等.基于高通量芯片對強直性脊柱炎的生物信息學分析[J].第三軍醫大學學報,2019,41(1):63-70.
[ 9 ] 郭明飛,高家榮,姜輝,等. 2型糖尿病模型大鼠肝脂代謝相關基因的篩選及生物信息學分析[J].中國生物化學與分子生物學報,2017,33(7):726-735.
[10] 曹丹,余樂,王麗春.肝癌相關差異表達基因的生物信息學分析及蛋白互作網絡構建[J].四川大學學報(醫學版),2018,49(6):899-903.
[11] 馮曉飛,馬遙,趙舒煊,等.骨肉瘤基因表達譜芯片的生物信息學分析[J].華中科技大學學報(醫學版),2019,48(3):281-286.
[12] 許丁文,熊彥,嚴慧深,等.基于生物信息學分析絲甘蛋白聚糖對卵巢癌耐藥性的影響及作用機制[J].中國藥房,2019,30(1):46-51.
[13] SCHELTENS P,BLENNOW K,BRETELER MM,et al. Alzheimer’s disease[J]. Lancet,2016,388(10043):505- 517.
[14] ZOU C,WANG J,HUANG X,et al. Analysis of transcription factor- and ncRNA-mediated potential pathogenic gene modules in Alzheimer’s disease[J]. Aging:Albany NY,2019,11(16):6109-6119.
[15] LIAN H,YANG L,COLE A,et al. NFκB-activated astroglial release of complement C3 compromises neuronal morphology and function associated with Alzheimer’s di- sease[J]. Neuron,2015,85(1):101-115.
[16] GAO Y,LI XI,LIU XH,et al. Determining association of rho kinase 1 gene polymorphisms with risk of Alzheimer’s disease:a multicenter pilot study[J]. Ann Transl Med,2018. DOI:10.21037/atm.2018.05.51.
[17] MOLOUDIZARGARI M,ASGHARI MH,GHOBADI E,et al. Autophagy,its mechanisms and regulation:implications in neurodegenerative diseases[J]. Ageing Res Rev,2017. DOI:10.1016/j.arr.2017.09.005.
[18] DINIZ LP,MATIAS I,SIQUEIRA M,et al. Astrocytes and the TGF-β1 pathway in the healthy and diseased brain:a double-edged sword[J]. Mol Neurobiol,2019,56(7):4653-4679.
[19] LI W,LIU H,YU M,et al. Folic acid alters methylation profile of JAK-STAT and long-term depression signaling pathways in Alzheimer’s disease models[J]. Mol Neuroibol,2016,53(9):6548-6556.
[20] FERNANDEZ AM,HERVAS R,DOMINGUEZ-FRAILE M,et al. Blockade of the interaction of calcineurin with FOXO in astrocytes protects against amyloid-β-induced neuronal death[J]. J Alzheimers Dis,2016,52(4):1471- 1478.
[21] SHIM SM,CHEON HS,JO C,et al. Elevated Epstein- Barr virus antibody level is associated with cognitive decline in the Korean elderly[J]. J Alzheimers Dis,2017,55(1):293-301.
[22] SOROKINA LN,MINEEV VN,LIM VV. Role of negative regulators of SOCS1,SOCS3,and SOCS5 gene transcription in the negative cell signaling regulation system in asthma[J]. Ter Arkh,2017,89(3):43-47.
[23] GUI T,HE BS,GAN Q,et al. Enhanced SOCS3 in osteoarthiritis may limit both proliferation and inflammation[J]. Biotech Histochem,2017,92(2):107-114.
[24] CHIKUMA S,KANAMORI M,MISE-OMATA S,et al. Suppressors of cytokine signaling:potential immune checkpoint molecules for cancer immunotherapy[J]. Cancer Sci,2017,108(4):574-580.
[25] DENG J,JIAO X,LIU H,et al. Lymph node metastasis is mediated by suppressor of cytokine signaling-3 in gastric cancer[J]. Tumour Biol,2013,34(6):3627-3636.
[26] JIANG BG,WANG N,HUANG J,et al. Tumor SOCS3 methylation status predicts the treatment response to TACE and prognosis in HCC patients [J]. Oncotarget,2017,8(17):28621-28627.
[27] KWAK YD,WANG B,LI JJ,et al. Upregulation of the E3 ligase NEDD4-1 by oxidative stress degrades IGF-1 receptor protein in neurodegeneration[J]. J Neurosci,2012,32(32):10971-10981.
[28] PERSAUD A,ALBERTS P,HAYES M,et al. NEDD4-1 binds and ubiquitylates activated FGFR1 to control its endocytosis and function[J]. EMBO J,2011,30(16):3259- 3273.
[29] WANG X,TROTMAN LC,KOPPE T,et al. NEDD4-1 is a proto-oncogenic ubiquitin ligase for PTEN[J]. Cell,2007,128(1):129-139.
[30] VECCHIONE A,MARCHESE A,HENRY P,et al. The Grb10/NEDD4 complex regulates ligand-induced ubiquitination and stability of the insulin-like growth factor Ⅰ receptor[J]. Mol Cell Biol,2003,23(9):3363-3372.
[31] SUN M,CAI J,ANDERSON RA,et al. Type Ⅰγ phosphatidylinositol phosphate 5-kinase Ⅰ5 controls the ubiquitination and degradation of the tumor suppressor mitogen-inducible gene 6[J]. J Biol Chem,2016,291(41):21461- 21473.
[32] LIU Q,ZHOU H,LANGDON WY,et al. E3 ubiquitin ligase CBLB in innate and adaptive immunity[J]. Cell Cycle,2014,13(12):1875-1884.
[33] FANG DF,HE K,WANG N,et al. NEDD4 ubiquitinates TRAF3 to promote CD40-mediated Akt activation[J]. Nat Commun,2014. DOI:10.1038/ncomms5513.
[34] LIYASOVA MS,MA K,LIPKOWITZ S. Molecular pathways:CBL proteins in tumorigenesis and antitumor immunity-opportunities for cancer treatment[J]. Clin Cancer Res,2015,21(8):1789-1794.
[35] TSYGANKOV AY,TECKCHANDANI AM,FESHCHENKO EA,et al. Beyond the RING:CBL proteins as multivalent adapters[J]. Oncogene,2001,20(44):6382-6402.
[36] THIEN CB,LANGDON WY. CBL:many adaptations to regulate protein tyrosine kinases[J]. Nat Rev Mol Cell? ? Biol,2001,2(4):294-307.
(收稿日期:2019-03-10 修回日期:2019-09-12)
(編輯:張元媛)
