拉關木小苗在白骨壤林內及林緣生長實驗研究
2019-09-21 02:28:16周亮王秀麗林嫻慧劉可鑫盧昌義
生態科學 2019年4期
周亮, 王秀麗, 林嫻慧, 劉可鑫, 盧昌義,*
拉關木小苗在白骨壤林內及林緣生長實驗研究
周亮1, 2, 王秀麗1, 林嫻慧1, 劉可鑫1, 盧昌義1, 2,*
1. 河口生態安全與環境健康福建省高校重點實驗室(廈門大學嘉庚學院), 漳州 363105 2. 廈門大學環境與生態學院, 廈門 361102
探討引進的紅樹植物拉關木()是否對本土紅樹植物白骨壤林()造成入侵風險。通過野外種植實驗, 將1年生拉關木苗種植在成熟白骨壤林的林內和林緣, 每月觀察記錄并測量其生長指標。結果表明: 白骨壤林內和林緣的1年生拉關木苗的月均苗高增量變化趨勢相似, 均表現為夏季高, 冬季低; 月均基徑增量表現為林內低于林緣, 且隨著種植時間的延長而降低。白骨壤林內和林緣的拉關木苗野外種植13個月后的苗高增量僅為14.1 cm和13.5 cm, 基徑增量僅為1.0 mm和1.5 mm, 葉片數增量均為負值, 分別為-0.4片和-0.7片, 可見外來種拉關木苗在廈門海滄灣的白骨壤林內及林緣生長速度緩慢, 不具有速生快長特性, 因此, 外來紅樹植物拉關木只有在其適宜生長的環境才能表現出速生的特性。種植13個月后, 白骨壤林內和林緣拉關木苗的存活率僅為4.7%和5.3%。表明由于光照強度、低溫等環境因子的限制, 引進種拉關木小苗很難通過自然擴散在廈門本土紅樹植物白骨壤林內自然更新生長。
拉關木苗; 白骨壤林; 光照強度; 生長指標; 外來種
0 前言
拉關木〔〕(L.) Gaertn. F., 屬于使君子科(Combretaceae), 是真紅樹植物的一種, 廣泛分布于南美、西印度群島、百慕大群島、西非以及佛羅里達沿岸[1]。拉關木屬于速生常綠喬木, 其高度通常小于15 m, 胸徑小于30 cm, 在特定環境下樹高會達到25 m, 胸徑達到70 cm[2]。由于經濟發展, 沿海地區過度開發, 導致毗鄰的紅樹林區域面積縮小, 中國現有的天然紅樹林面積目前僅為15000 hm2[3]。為提高我國紅樹林的種類多樣性和河口海岸帶紅樹林防護工程質量, 中國林業科學研究院熱帶林業研究所的研究人員于1999年從墨西哥拉巴斯市首次引入拉關木, 種植于海南東寨港紅樹林保護區的引種園內[4]。近年來, 在廣東、福建等地也陸續引進該種造林。由于拉關木的速生快長特性, 其是否會對本土紅樹植物造成入侵值得關注。影響紅樹林下層植被分布的因素主要是: 光照強度、潮汐、種子大小及植物本身的生長特性, 其中, 光照強度的影響受到學者們普遍關注[5]。陳玉鋒等[6]研究表明, 光照是限制植物紅毛五加()光合速率的主要影響因素。眾多的研究者對紅樹林的深入研究均表明: 光照強度對紅樹林的生長、發育、繁殖、種群變化和群落演替均有著重要的影響[7-12]。生態入侵的表現往往體現在對光照條件“生態位”的競爭方面。作為外來引進種, 拉關木與鄉土紅樹植物生態位的關系值得關注。目前尚未見拉關木苗在本土紅樹植物白骨壤林內及林緣的生長情況的研究報道。
本文選取光照強度這個生態限制因子, 分析拉關木通過樹冠間隙(Canopy gap)與本地已成林紅樹植物白骨壤林的競爭可能性和動態過程。鐘才榮等[13]對紅樹植物拉關木的引種效果調查研究表明, 6個月生的拉關木苗在海南東寨港的苗高比廣東電白高出6 cm, 而基徑則比廣東電白的小0.2 cm。表明苗高和基徑可以反映不同省份地區拉關木苗的生長量情況。楊虎彪和劉國道[14]對不同光照強度對幼齡期鷓鴣茶生長的影響的研究結果表明, 不同光照強度對鷓鴣茶葉片數及株高均有影響, 光照強度73%的鷓鴣茶葉片數明顯高于光照強度28%處理。因此, 本研究通過1年生拉關木苗在鄉土紅樹植物白骨壤林的林內及林緣的苗高、基徑、葉片數的測定, 對1年生拉關木苗的生長過程和狀態進行定性、定量分析, 旨在了解拉關木在低光強環境中的生態特性及其在鄉土林內生境中的生態入侵風險評估, 為今后合理引進該樹種以及紅樹林生態系統的保護管理提供參考。
1 材料與方法
1.1 試驗地自然概況
研究區域位處福建省廈門市海滄區海滄灣沿岸的紅樹林區(118°03′E, 24°29′N)。該區屬南亞熱帶海洋性季風氣候, 年平均氣溫21.3 ℃, 1月份平均氣溫13.4 ℃, 7月份平均氣溫29.4 ℃, 極端最低氣溫4.3 ℃, 極端最高氣溫36.6 ℃, 年日照時間1971 h, 年降水量1084.5 mm[15]。試驗樣地為成熟的天然白骨壤林, 郁閉度約65.0%, 高1.8—2.0 m, 基徑5.0—7.0 cm。試驗樣地白骨壤林生長區為中潮帶。
1.2 試驗材料
2014年6月從廣東省湛江市徐聞縣紅樹林育苗苗圃選購1年生拉關木苗, 將拉關木苗放廈門大學漳州校區生化樓A棟1樓的廊道空間, 溫度、光照均較穩定, 將拉關木苗擺在育苗盆內, 定期給小苗澆水, 定植培養1個月。
1.3 試驗設計
所選的試驗樣地廈門市海滄區海滄灣沿岸的白骨壤林情況如圖1所示, 根據現場試驗樣地實際情況的考察, 該片白骨壤林附近光灘的底質和白骨壤林內、林緣的差別大, 白骨壤林試驗樣地附近光灘的底質主要為細石頭和砂質, 而白骨壤林內主要為黑色底泥。由于本試驗主要研究外來紅樹植物拉關木是否會對本土紅樹植物白骨壤林有入侵的風險, 考慮的主要環境因素是光因子, 而離白骨壤林樣地較遠的光灘也會因底質、高程、海浪等環境因子的不同, 不適合作為對照樣地。因此, 本研究根據試驗樣地的現場情況設置了白骨壤林的林內及林緣各設3個重復的平行樣地, 共6個實驗樣地, 選取高度一致、生長良好的1年生拉關木苗300株, 每個樣地種植50株。試驗樣地林內、林緣的底質、溫度、灘涂高程等相近。實驗前對每株實驗苗懸掛標志牌, 標注各實驗苗的編號。同時對實驗苗各生長指標進行測量, 測量其基徑、株高、葉片數, 作為日后實驗數據對比值。種植后, 以30 d為監測周期, 于每月底測量各樣地拉關木苗生長指標(苗高、基徑、葉片數)測定, 并統計各樣地拉關木苗的成活率。
1.4 野外光照強度監測
采用臺灣泰士TES—1332數位式照度計(單位), 分別測定各樣地林外、林緣、林內光照強度, 以種植的拉關木苗頂端的高度為光強測量高度, 選擇晴朗天氣, 于退潮期, 對每個樣地以每小時為頻度進行光照強度測定, 取其平均值作為該樣地的光照強度。
1.5 數據處理
用Excel進行統計分析和作圖, 采用SPSS 17.0軟件對數據進行單因素方差分析, p<0.05表示差異顯著。

Figure 1 The test plots offorest in Haicang Bay, Xiamen City
2 結果與分析
2.1 白骨壤林各樣地的光照強度情況
2015年2月, 選擇晴朗天氣, 樣地處于退潮期, 采用臺灣泰士TES—1332數位式照度計(單位Lux), 分別測定各樣地林外、林緣、林內的光照強度,測定結果如圖2所示, 樣地白骨壤林緣的光照強度為林內的2.7倍。
2.2 白骨壤林內及林緣拉關木苗生長動態分析
由圖3可知, 白骨壤林內、林緣1年生拉關木苗種植初期(即2014年7月底), 其月均苗高增量較低, 僅為1.13 cm和0.99 cm; 野外種植2個月后(即2014年8月底), 白骨壤林內和林緣1年生拉關木苗的月均苗高增量分別達3.48 cm和2.75 cm。林內和林緣的月均苗高增量變化趨勢相似, 均表現為夏季高, 冬季低。由于2014年12月至2015年3月, 極端低溫的脅迫, 導致2015年2月至4月, 拉關木小苗的苗高增量減少, 次年3月, 林內和林緣的月均苗高增量達最小值, 僅為0.22 cm和0.28 cm。2015年5月后, 隨著氣溫的升高, 月均苗高增量有所增加。
由圖4可知, 月均基徑增量隨種植時間的變化趨勢與月均苗高增量相似, 均隨著種植時間的延長呈先增加后減少的變化趨勢。野外白骨壤林樣地種植第1個月, 由于野外環境的適應, 林內和林緣的拉關木小苗的月均基徑增量分別僅為0.13 mm和0.19 mm; 林內和林緣的月均基徑增量在2014年8月份達最大值, 分別達0.28 mm和0.50 mm; 到2014年11月(即野外種植第5個月)后, 林內和林緣樣地的拉關木苗的月均基徑增量均少于0.1 mm。
由圖5可知, 野外種植第1個月, 白骨壤林內和林緣拉關木苗的月均葉片數增量分別為–0.9片和–0.4片,剛種植一個月內出現落葉現象主要原因是: 1年生拉關木苗從室內定植培養到野外環境, 由于剛種植的苗根系有輕微受損, 且野外由于晝夜溫差大、海浪等環境因子的干擾, 需要一段時間的定植恢復期。但在2014年8月份(即野外種植第2個月), 拉關木苗的月均葉片數增量有所增加, 林內和林緣的月均葉片數增量分別為0.6片和4.0片。在2014年9月份至2015年4月份期間, 白骨壤林內和林緣的1年生拉關木苗的月均葉片數增量均為負值, 出現落葉現象; 2015年5月份, 林內和林緣樣地的拉關木苗的月均葉片數增量均有所增加, 分別達1.6片和2.4片。但在2015年6月份, 白骨壤林緣的1年生拉關木苗的月均葉片數增量減少, 到2015年7月份時出現葉片凋落現象; 而2015年6—7月份, 林內的1年生拉關木苗的葉片再次出現凋落現象, 海滄白骨壤林內及林緣1年生拉關木苗葉片數隨時間變化出現波動情況, 可能與樣地外圍商業船舶、漁船的出入, 導致對白骨壤林樣地造成較大的海浪沖擊有關。
Figure 2 Light intensity offorests in Haicang Bay in February 2015
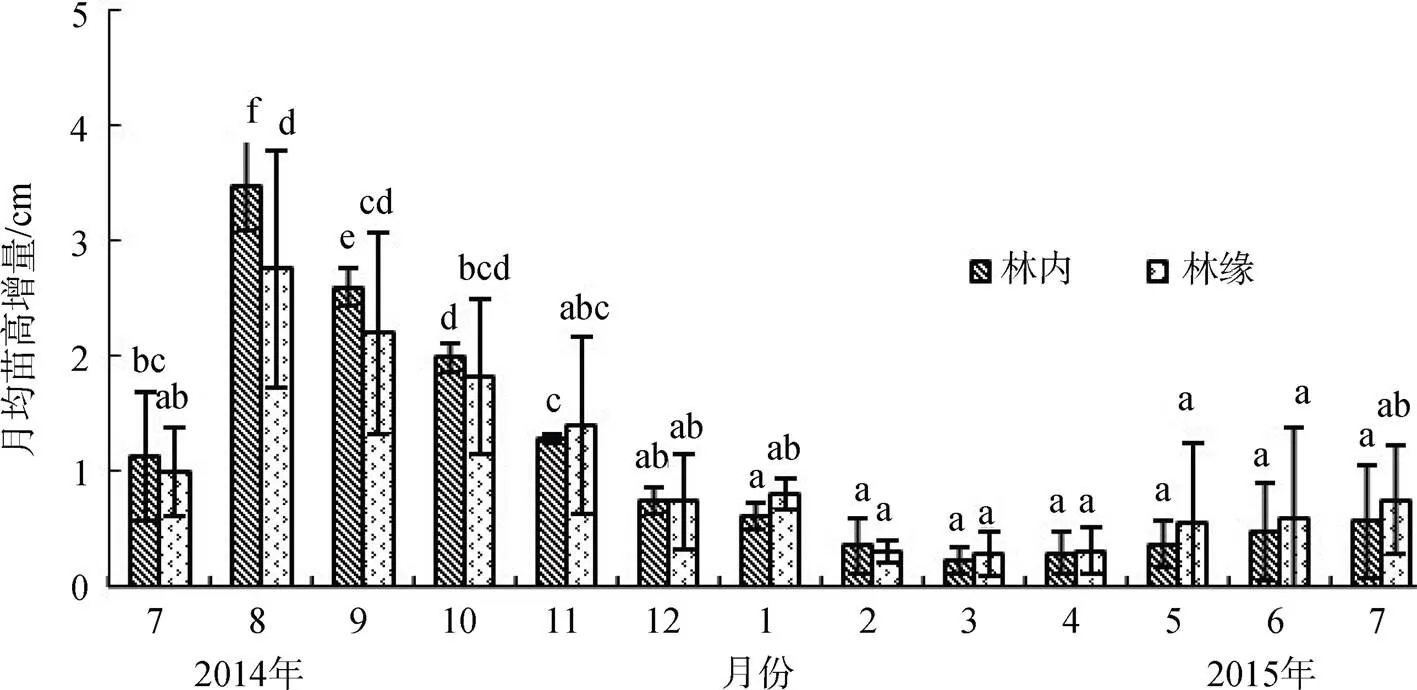
圖3 不同光照強度下白骨壤林的拉關木小苗月均苗高增量
Figure 3 Monthly average height increment ofseedlings under different light intensity inforests

圖4 不同光照強度下白骨壤林的拉關木小苗月均基徑增量
Figure 4 Monthly average base diameter increment ofseedlings under different light intensity inforests
由表1, 表2可見, 在2014年7月至2015年6月期間, 海滄白骨壤林內的1年生拉關木小苗的存活率均高于林緣。由于3個試驗樣地海滄白骨壤林內的郁閉度僅為60.1%—65.3% (如表1所示), 因此白骨壤林內的1年生拉關木苗能存活較長時間, 但到2015年7月, 白骨壤林內和林緣的拉關木苗的存活率僅為4.7%和5.3%。
3 討論
光照是植物生長必不可少的要素, 光照強度的強弱直接影響到植物的光合作用, 最終影響植物的生長。本研究結果表明, 野外種植6個月的時間內, 成熟白骨壤林內的1年生拉關木苗的月均苗高增量普遍高于林緣, 2014年8月份, 白骨壤林內1年生拉關木苗的月均苗高增量比林緣高26.5%, 出現這種情況的原因可能是因試驗樣地的白骨壤林的郁閉度為60.1%—65.3%, 光線均能透進白骨壤林內, 弱光促進了1年生拉關木苗的苗高的增長。這與弱光對無瓣海桑幼苗的莖高有促進作用[16]的結果相似。有研究表明, 弱光對多數植物幼苗和秋茄()胎生幼苗的初期生長有促進作用的研究結果相似[8, 17]。這可能是低光照導致紅樹林植物的光合速率下降, 植物生長激素分泌增加, 從而致使植物個體的莖高增加[5]。白骨壤林緣和林內樣地的1年生拉關木苗的月均苗高增量在種植第2個月時均達到最大值, 分別達3.48 cm和2.75 cm。種植第3個月后, 林內和林緣的1年生拉關木苗的月均苗高增量、月均基徑增量、及月均葉片數增量均呈下降趨勢, 表明長時間的弱光條件不利于拉關木幼苗的生長; 這與刁俊明等[18]研究表明: 無瓣海桑在蔭蔽初期(2—4個月), 弱光環境對幼苗的莖高的增加有促進作用, 隨著蔭蔽時間的延長則產生抑制作用, 不利于幼苗的生長的結論一致。鐘才榮等[13](2011)研究表明, 拉關木在海南東寨港保護區6個月齡的苗高為86 cm, 在廣東電白紅樹林苗圃6個月齡的苗高為74 cm。而本研究中, 白骨壤林內和林緣的拉關木苗野外種植13個月的苗高增量僅為14.1 cm和13.5 cm, 基徑增量僅為1.0 mm和1.5 mm, 可見外來種拉關木苗在廈門海滄灣的白骨壤林內和林緣的生長速度極慢, 不具有速生快長特性, 因此, 外來紅樹植物拉關木只有在其適宜生長的環境下才能表現出速生快長的特性。

圖5 不同光照強度下白骨壤林的拉關木小苗月均葉片數增量
Figure 5 Monthly average number of leaves increment ofseedlings under different light intensity inforests

表1 不同光照強度下白骨壤林的拉關木小苗2014年的存活率

表2 不同光照強度下白骨壤林的拉關木小苗2015年的存活率
冬季至次年的初春季, 白骨壤林內和林緣的1年生拉關木苗的月均苗高增量、月均基徑增量均非常低, 在次年3月份1年生拉關木苗的月均苗高增量達最小值, 分別為0.22 cm和0.28 cm, 葉片出現凋落, 這可能是因為低溫導致1年生拉關木小苗的生長緩慢甚至出現落葉現象。因本土紅樹植物白骨壤是自然分布最北的抗低溫廣布種紅樹[19], 表明不耐低溫的引進種拉關木小苗難于在福建較北的地方占據本土紅樹植物白骨壤林的生態位。
本研究結果表明, 1年生拉關木苗在白骨壤林內和林緣種植一個月后, 存活率為86.0%和74.0%, 隨著種植時間的延長, 其存活率不斷減少, 且林內1年生拉關木苗的存活率普遍低于林緣, 這可能是因為試驗樣地白骨壤林為海滄灣的入海口處, 該試驗片區的海浪沖擊大, 白骨壤林緣的1年生拉關木苗由于海浪沖擊大, 而白骨壤林內海浪沖擊小, 導致拉關木苗種植1年時間里, 林內的存活率均高于林緣。種植6個月后, 林內和林緣拉關木苗的存活率為45.3%和32.0%, 到次年7月份林內和林緣的1年生拉關木苗的存活率僅為4.7%和5.3%。因此, 由于光照強度、低溫等環境因子的限制, 引進種拉關木小苗很難通過自然擴散在廈門本土紅樹植物白骨壤林內自然更新生長。
[1] NELLIS D. Seashore plants of South Florida and the Caribbean[M]. Sarasota: Pineapple Press, 1994: 164.
[2] JIMENEZ J, MARTINEZ R, ENCARNOCION L. Massive tree mortality in a Puerto Rican mangroves forests[J]. Caribbian Journal of Science, 1985, 21:75–78.
[3] PENG Y, ZHOU Y, Chen G Z. The restoration of mangrove wetland: a review[J]. Acta Ecologica Sinica, 2008, 28: 786–797.
[4] 廖寶文, 鄭松發, 陳玉軍, 等. 海南東寨港幾種國外紅樹植物引種初報[J]. 中南林學院學報, 2006, 26(3): 63–67.
[5] 刁俊明, 彭逸生, 鄭明軒, 等. 紅樹植物對低光照水平的生長及生理生態響應研究進展[J]. 嘉應學院學報, 2009, 27(3): 69–74.
[6] 陳玉鋒, 黃旭峰, 古銳, 等. 不同光照強度下紅毛五加光合及生理特性研究[J]. 中國中藥雜志, 2018, 43(5): 926–933.
[7] SMITH T. Effects of seed predators and light level on the distribution of(Forsk) Vierh in topical, tidal forests[J]. Estuarine, Coastal and Shelf Science, 1987, 25(1): 43–51.
[8] TAMAI S, IANPA P. Establishment and growth of mangrove seedlings in mangrove forests of southen Thaliland[J]. Ecological Research, 1988, 3(3): 227–238.
[9] ALLEN J, KRAUSS K, HAUFF R. Factors limiting the intertidal distribution of the mangrove species[J]. Oecologia, 2003, 135(1): 110–121.
[10] PROFFITT C, DEVLIN D. Long-term growth and succession in restored and natural mangrove forests in southwestem Florida[J]. Wetlands Ecology and Mana-gement, 2005, 13(5): 531–551.
[11] IMAI N, TAKYU M, NAKAMURA Y, et al. Gap formation and regeneration of tropical mangrove forests in Ranong Thailand[J]. Plant Ecology, 2006, 186(1): 37–46.
[12] SANTIAGO L, LAU T, MELCHER P, et al. Morphological and physiological responses of Hawaiian hibiscus tiliaceus populations to light and salinity[J]. International Journal of Plant Sciences, 2000, 161(1): 99–106.
[13] 鐘才榮, 李詩川, 楊宇晨, 等. 紅樹植物拉關木的引種效果調查研究[J]. 福建林業科技, 2011, 38(3): 96–99.
[14] 楊虎彪, 劉國道. 不同光照強度對幼齡期鷓鴣茶生長的影響[J]. 熱帶作物學報, 2017(11): 2056–2059.
[15] 廈門市統計局, 國家統計局廈門調查隊. 廈門經濟特區年鑒—2015 [M]. 北京: 中國統計出版社, 2015.
[16] 刁俊明, 陳桂珠. 光強對無瓣海桑幼苗的生長和光合特性的影響[J]. 林業科學研究, 2008, 21(4): 496–492.
[17] PROFFITT C E, DEVLIN D J. Long-term growth and succession in restored and natural mangrove forests in southwestern Florida[J]. Wetlands Ecology and Mana-gement, 2005, 13(1): 531–551.
[18] 刁俊明, 曾憲錄, 陳桂珠. 無瓣海桑幼苗對不同遮光度的生理生態響應[J]. 生態學雜志, 2010, 29(7): 1289– 1294.
[19] 林鵬. 紅樹林[M]. 北京: 海洋出版社, 1984.
Growth ofseedlings inside and at the edge offorests
ZHOU Liang1,2, WANG Xiuli1, LIN Xianhui1, LIU Kexin1, LU Changyi1,2,*
1. Key Laboratory of Estuarine Ecological Security and Environmental Health (Xiamen University Tan Kah Kee College), Fujian Province University, Zhangzhou 363105, China 2. College of the Environment & Ecology, Xiamen University, Xiamen 361102, China
In order to assess the invasion risk of fast-growing introduced mangrove species, field experiments of planting 1-year-oldseedlings were used to study the effects of environmental factors of light intensity on their growth inside and at the edge of native mangrove speciesforest. Results showed that the average monthly shoot height of 1-year-oldseedlings inside and at the edge of theforests was similar, both high in summer and low in winter; the average monthly shoot diameter of 1- year-oldseedlings inside theforests was lower than that inforest margins, and the average monthly shoot diameter increment ofseedlings decreased with the prolongation of planting time. After planting for 13 months, the seedling height ofinside and at the edge of native mangrove speciesforest was only 14.1 and 13.5 cm, respectively; and the base diameter increment was only 1.0 and 1.5 mm, respectively. The average monthly number leaves of 1-year-oldseedlings inside and at the edge of theforest decreased to -0.4 and -0.7, respectively. The results indicated that the growth rate of exotic mangrove specieswas slowly inside and at the edge offorestsin Haitang Bay, Xiamen city; and it did not show fast-growing characteristics. Therefore, the exotic mangrove speciescan grow fast only in its suitable environment. The survival rate ofseedlings inside theforests was only 4.7%, and that inforest margin was 5.3%. Due to the limitation of light intensity, low temperature, it is very difficult forseedlings to naturally renew and grow under natural conditions in the natural indigenousmangrove forest in Xiamen.
seedling;forests; light intensity; growth indexes; exotic spe
10.14108/j.cnki.1008-8873.2019.04.001
Q948.1
A
1008-8873(2019)04-001-06
2018-09-11;
2018-11-05
國家重點研發計劃“典型脆弱生態修復與保護研究”重點專項(2017YFC0506103); 國家自然科學基金(41376115), 福建省中青年教師教育科研項目(JA15618), 福建省大學生創新創業訓練計劃項目(201913469024)
周亮(1981—), 男, 福建廈門人, 講師, 主要從事紅樹林濕地生態及環境遙感方面的研究, E-mail: zhou812@xujc.com
盧昌義, 教授, 博士生導師, 主要從事紅樹林濕地海岸生態恢復工程方面研究, E-mail: lucy@xmu.edu.cn
周亮, 王秀麗, 林嫻慧, 等. 拉關木小苗在白骨壤林內及林緣生長實驗研究[J]. 生態科學, 2019, 38(4): 1-6.
ZHOU Liang, WANG Xiuli, LIN Xianhui, et al. Growth ofseedlings inside and at the edge offorests[J]. Ecological Science, 2019, 38(4): 1-6.
