多巴胺系統基因與母親教養行為對青少年抑郁的影響:一項多基因研究*
2019-10-17 01:00:18曹衍淼張文新
心理學報 2019年10期
曹衍淼 張文新
多巴胺系統基因與母親教養行為對青少年抑郁的影響:一項多基因研究
曹衍淼 張文新
(山東師范大學心理學院, 濟南 250014)
近年來, 伴隨著對單基因研究局限性的認識及對“遺傳率缺失”的探索, 越來越多的研究強調考察抑郁的多基因遺傳機制的重要性。本研究對1052名漢族青少年(12.31 ± 0.37, 50.2%女生)進行一年的追蹤, 采用多基因累加得分研究范式考察多巴胺系統基因與母親教養行為對青少年抑郁的縱向影響及其作用模式。結果發現:(1)多基因累加得分和母親消極教養正向預測青少年抑郁風險; (2)控制早期抑郁后, 多基因累加得分與母親積極、消極教養交互影響青少年抑郁, 在低積極/高消極教養環境中, 相比多基因累加得分較低的青少年, 多基因累加得分較高的青少年抑郁水平更高; 但是在高積極/低消極教養環境中, 不同多基因累加得分的青少年抑郁水平無差異。該交互作用模式符合“素質?壓力”模型。研究結果為抑郁的多基因遺傳基礎提供了證據。
青少年抑郁; 母親教養行為; 多巴胺; 多基因累加得分; 基因×環境
1 問題提出
青少年抑郁障礙的影響因素和發生機制是發展心理病理學的重要熱點研究課題。隨著分子遺傳學的興起, 過去十幾年眾多研究者從單基因×環境相互作用(G × E)的角度, 考察了遺傳基因對青少年抑郁的影響(e.g., Xia & Yao, 2015)以及基因與環境因素對青少年抑郁的交互作用(e.g., Cao et al., 2018; Zhang et al., 2015; 曹衍淼, 王美萍, 曹叢, 紀林芹, 張文新, 2017)。盡管單基因研究為理解抑郁的發生機制與個體差異提供了重要啟示, 但是單基因對青少年抑郁的效應量通常不足2%, 而且研究結果的可重復性較低(e.g., Dick et al., 2015)。近年來越來越多的證據顯示, 抑郁具有復雜的多基因遺傳結構。譬如, 數量遺傳學研究以多基因(polygenic)和累加性(additive)假設為基礎進行遺傳率估計, 發現遺傳因素能夠解釋抑郁30% ~ 70%的變異(e.g., Nivard et al., 2015)。伴隨著對單基因研究局限性的認識及“遺傳率缺失”的探討, 分子遺傳學領域的研究者開始采用多基因累加得分及基因×基因交互的方法探索抑郁復雜的多基因遺傳機制和作用過程(e.g., Cao, Lin, Chen, Ji, & Zhang, 2018; Stocker et al., 2017)。本研究即采用多基因視角, 考察多巴胺(dopamine, DA)系統多基因累加得分與母親教養對青少年抑郁的影響, 并檢驗該多基因×環境交互模式符合何種理論假說(“素質?壓力”模型與“不同易感性”模型), 以期豐富抑郁的多基因遺傳機制研究。
1.1 多巴胺系統基因與抑郁:從單基因到多基因研究
依據單胺缺陷假說(monoamine deficiency hypothesis), 多巴胺系統功能失調是引發抑郁的重要原因(Belmaker & Agam, 2008)。多巴胺系統功能受到多巴胺代謝、轉運和傳導多個環節的共同影響(Opmeer, Kortekaas, & Aleman, 2010)。此外, 越來越多的研究顯示中腦?邊緣多巴胺通路(mesolimbicpathway)和中腦?皮質多巴胺通路(mesocortical pathway)功能缺陷與抑郁密切相關(Dunlop & Nemeroff, 2007)。基于此, 在眾多多巴胺系統基因中, 既參與調節多巴胺代謝、轉運和傳導, 又表達于中腦?邊緣、中腦?皮質多巴胺通路中的基因位點受到了研究者的廣泛關注。譬如, 兒茶酚胺氧位甲基轉移酶基因(catechol-O-methyltransferase,)、多巴胺轉運體基因(dopamine transporter,)及D2型多巴胺受體基因(dopamine receptor D2,) (Cao et al., 2018; Lin et al., 2017; Pinsonneault et al., 2011)。
基因編碼的COMT酶能夠降解去甲腎上腺素、腎上腺素、多巴胺等神經遞質, 降低突觸間隙中的多巴胺濃度進而導致抑郁。在眾多基因多態性中, 多數研究者關注rs4680(Val158Met)多態性與抑郁等心理病理問題的關聯, 但是研究顯示在亞洲樣本中, rs6267多態性與COMT酶活性的關聯強于rs4680多態性(Lee et al., 2005)。基于此, 本研究重點關注位于22號染色體q11.2區的rs6267多態性。在該多態性中, 與T等位基因相比, G等位基因與更高COMT酶活性有關(Lee et al., 2005), 即G等位基因具有較高的多巴胺代謝率進而會導致較低的多巴胺濃度。Lin等(2017)的研究顯示, rs6267多態性與帕金森病人的抑郁癥狀顯著相關。
基因編碼的多巴胺轉運體能夠將突觸間隙的多巴胺轉運再攝取到突觸前膜, 調節腦內多巴胺含量進而影響抑郁。現有研究主要考察了VNTR多態性, 但是在何種等位基因(9R或10R)與較高的DAT表達活性有關上仍存在分歧(e.g., Costa, Riedel, Müller, M?ller, & Ettinger, 2011; Heinz et al., 2000)。相比之下, 位于5號染色體p15.33區的rs27072多態性表達活性更為穩定, 在體內和體外實驗中一致發現T等位基因比CC基因型具有更高的表達水平(Pinsonneault et al., 2011)。有關雙相障礙的研究顯示, 在多個重復驗證樣本中, 相比其他基因位點, rs27072多態性與雙相障礙存在穩定關聯(Pinsonneault et al., 2011)。
基因編碼的D2型多巴胺受體通過影響中腦邊緣系統的多巴胺能活性從而影響罹患抑郁的風險。rs1799978多態性位于11號染色體q2.2-2.3區, 是基因啟動子區的一種功能性多態性位點, 在-241位置處腺嘌呤(A)置換鳥嘌呤(G)形成兩種等位基因(Doehring, Kirchhof, & L?tsch, 2009), 影響D2受體的表達(Zhang & Malhotra, 2011)。盡管尚未有關于rs1799978等位基因功能的直接證據, 但是相關研究提示A等位基因可能與更低的多巴胺受體活性(Doehring et al., 2009)及更高的抑郁風險有關(Cao et al., 2018)。具體而言, 有關利培酮(通過與D2受體結合產生拮抗作用以減少抑郁、精神分裂癥狀)的藥物研究顯示, 相比G等位基因, A等位基因或AA基因型攜帶者在治療后表現出更好的療效(Doehring et al., 2009)。這提示, A等位基因可能與較低的D2受體數量相關, 較低的D2受體數量可以使得利培酮與受體結合率更高, 從而起到更好的療效。此外, 相關研究顯示在亞裔樣本中, 相比其他多態性, rs1799978多態性與更好的利培酮療效有關(Xing et al., 2007), 并且與抑郁的關聯更強(Cao et al., 2018)。
盡管單基因研究為理解抑郁的發生機制與個體差異提供了重要啟示, 但是單基因研究的解釋率往往極低。造成微弱的遺傳解釋率的原因之一可能是研究忽視了抑郁的多基因遺傳結構(Belsky & Pluess, 2009)。尤其是, 研究顯示基因、基因和基因可能并非獨立作用于抑郁, 而是存在聯合效應。如早期的動物研究發現, 在基因敲除鼠中, COMT酶含量比未敲除鼠高400%, D2受體表達水平下降(Giros, Jaber, Jones, Wightman, & Caron, 1996)。這表明基因、基因和基因間可能存在聯合效應, 共同調節多巴胺系統的功能。
近期, 研究者通過累加個體攜帶的易感等位基因條數測量多基因聯合效應, 即多基因累加得分(multilocus genetic profile score, MGPS)。多基因累加得分方法為抑郁的遺傳機制研究提供了一種新的框架, 迄今已有研究顯示多基因效應比任何單一位點的效應更強(Pearson-Fuhrhop et al., 2014; Vrshek- Schallhorn et al., 2015)。譬如, Pearson-Fuhrhop等人(2014)依據單胺缺陷假說, 考察了影響多巴胺代謝、轉運和傳導功能的5種DA基因(、、、和)的多基因累加得分與抑郁癥的關聯, 結果顯示無論是在抑郁患者組還是健康對照組, 個體攜帶的風險等位基因(低活性多巴胺等位基因)數量越多, 其抑郁水平越高, 且相比單個基因位點, 多基因累加得分的解釋率更高。這提示多基因累加方法能夠提供更豐富的遺傳信息并且增加遺傳解釋力。此外, 在統計分析方面, 多基因累加得分可作為一種連續變量, 在檢驗基因×環境交互作用時更具有統計優勢(Dunn et al., 2011)。
1.2 多基因累加得分與環境因素的交互作用
理論和實證研究顯示, 多巴胺系統多基因不僅聯合影響個體罹患抑郁的風險, 并且可能調節個體對環境的敏感性。譬如, 生物社會發展模型(biosocial developmental model, BDM)指出, 多巴胺含量下降會引發青少年行為趨向系統功能缺陷, 增強個體在面對不利環境時的敏感性, 導致個體罹患內外化問題的風險提高(Beauchaine, Gatzke-Kopp, & Mead, 2007)。前額葉晚熟理論認為, 邊緣系統?前額葉連通性影響個體對環境的敏感性, 進而與抑郁情緒密切相關(Andersen & Teicher, 2008)。、和基因的主要表達腦區分別集中于大腦前額葉區域(Matsumoto et al., 2003)、中腦紋狀體及邊緣系統區域(Lewis, Melchitzky, Sesack, Whitehead, & Sampson, 2001; Noble, Gottschalk, Fallon, Ritchie, & Wu, 1997)。因此, 作用于邊緣系統?前額葉通路的三種基因可能聯合調節個體對環境的敏感性進而影響抑郁的發生發展。實證研究也為多巴胺系統多基因累加得分與環境的交互作用提供了初步的證據。如Davies, Cicchetti和Hentges (2015)的研究顯示, 在經歷母親消極教養行為時, 多基因累加得分較高的青少年比多基因累加得分較低的青少年問題行為更多。關于腹側紋狀體激活水平的研究顯示, 在面對獎賞信息反饋時, 個體攜帶的低活性多巴胺系統等位基因(基因、基因、基因和基因)數量越多, 則表現出更低的紋狀體反應性(Nikolova, Ferrell, Manuck, & Hariri, 2011)。
然而, 迄今少數幾項多巴胺系統多基因累加得分×環境研究主要關注不利的家庭環境因素, 如消極教養(Davies et al., 2015)、產前不利環境(Bischoff et al., 2017)、早期虐待(Coley, Sims, & Carrano, 2017)等, 尚未有研究關注積極環境的作用。在眾多的家庭環境因素中, 母親積極和消極教養行為是影響青少年抑郁的重要預測源。研究顯示, 母親的積極教養行為, 如溫情、支持性的教養行為會促進青少年積極的自我知覺和較好的社會情緒功能, 進而降低抑郁的風險(Olino et al., 2016; Wang et al., 2016)。相反, 消極教養行為, 如拒絕、嚴厲等會導致青少年形成較差的自我意識和自尊, 繼而導致較高的抑郁風險(Wang et al., 2016)。因此, 本研究的目的之一是綜合運用積極和消極教養指標, 考察基因、基因和基因多基因累加得分與環境的潛在交互作用。
1.3 “素質?壓力”模型與“不同易感性”模型
多基因累加得分與環境的交互作用模式可能被兩種競爭假設所解釋:“素質?壓力”模型與“不同易感性”模型。前者假設一些心理疾患或行為問題的產生是由于個人攜帶的風險基因和不利環境的綜合結果(Monroe & Simons, 1991); 而后者認為, 所謂的風險基因事實上是一種遺傳可塑性, 更具可塑性的個體對消極和積極環境均更加敏感(Belsky & Pluess, 2009)。基于這兩種競爭模型, 研究者對多基因累加得分本質上是一種“累加風險得分”還是“累加可塑性得分”亦存在紛爭。
然而問題是, 當前的多巴胺系統多基因累加得分研究中所測量的環境指標主要局限于消極環境, 尤其是當其取值范圍有限時, 采用傳統回歸方法可能無法反映交互作用的全貌, 其所驗證的“素質?壓力”模型可能反映了“不同易感性”模型的一部分(Roisman et al., 2012)。基于此, 研究者提出了更為準確的、新興的統計檢驗方法——再參數化回歸模型(re-parameterized regression model), 能夠從理論假設出發檢驗“基因×環境”交互作用的形式(Widaman et al., 2012)。因此, 本研究的第二個目的是采用新興的再參數化回歸模型驗證兩種競爭假設。
綜上, 本研究采用多基因研究視角, 考察多巴胺系統基因累加得分(、和)與母親教養行為對青少年抑郁的影響, 主要探討以下問題:(1)多基因累加得分和母親教養行為對青少年抑郁的交互作用; (2)對多基因累加得分與母親教養行為影響抑郁的交互作用形式(“素質?壓力”模型vs.“不同易感性”模型)進行檢驗。
2 研究方法
2.1 被試
被試來自國內一項大型追蹤項目。該項目最初在山東省濟南市的14所小學整群抽取三年級小學生2261名, 在每年第二學期進行一次追蹤測查。研究顯示, 在童年期, 抑郁的發生率和嚴重程度較低(Cole et al., 2002), 但是六、七年級(約12~13歲)是抑郁水平迅速上升的關鍵期, 而之后抑郁水平的增長速度減緩并趨近于成人水平(Cole et al., 2002; Ferro, Gorter, & Boyle, 2015; Natsuaki, Biehl, & Ge, 2009), 因此本研究選擇在青少年抑郁快速上升的年齡階段進行研究。鑒于工作量和研究經費問題, 以各學校班級為單位隨機抽取部分被試(1086人)進行遺傳數據收集。根據Duncan和Keller (2011)提出的遺傳研究統計檢驗力與樣本量需求, 當估計遺傳效應量在1%左右時, 為獲得80%以上的統計檢驗力, 樣本量要保證在600人以上, 本研究被試量滿足最低樣本量要求。具有遺傳數據的被試與原始樣本在基本研究變量(性別、年齡、抑郁、母親教養等)上均無顯著差異(性別: χ= 2.19,= 1,= 0.14; 母親受教育水平: χ= 2.83,= 2,= 0.24; 父親受教育水平: χ= 2.61,= 2,= 0.27; 家庭月收入: χ= 4.87,= 2,= 0.09; 年齡:(3185) = 0.12,= 0.91; T1抑郁:(3218) = 1.53,= 0.13; T2抑郁:(3077) = 0.80,= 0.43; 積極教養:(3206) = –0.91,= 0.36; 消極教養:(3206) = 0.76,= 0.45)。同時, 為避免人口分層偏差, 本研究僅在漢族被試(1052人)中進行分析, 漢族被試與少數民族被試在基本研究變量上亦無顯著差異(性別: χ= 0.00,= 1,= 0.98, 年齡:(1063) = 1.47,= 0.14; COMT: χ= 3.71,= 2,= 0.168; DAT1: χ= 0.55,= 2,= 0.76; DRD2: χ= 1.19,= 2,= 0.55; T1抑郁:(1072) = 0.06,= 0.96; T2抑郁:(1077) = 0.49,= 0.63; 積極教養:(1073) = 0.32,= 0.75; 消極教養:(1073) = 0.03,= 0.97)。最終, 本研究被試共1052人, 六年級時平均年齡為12.31 ± 0.37歲, 男生524人(49.8%), 女生528人(50.2%)。母親受教育水平在初中及以下者占11.7%, 高中或中專者占26.5%, 大專以上者占61.8%。父親受教育水平在初中及以下者占8.3%, 高中或中專者占20.6%, 大專以上者占71.0%。家庭月收入低于3000元的占23.2%, 3000 ~ 6000元的占47.9%, 高于6000元的占28.8%。
2.2 研究工具
2.2.1 母親教養行為
采用中文版兒童養育實踐問卷(Child-Rearing Practices Report) (Chen, Bian, Xin, Wang, & Silbereisen, 2010)測量母親教養行為, 該問卷由母親報告。采用溫情(4個項目, 如 “我以溫和、親切的態度和孩子說話”)、引導(4個項目, 如“當我的孩子遇到什么問題時, 我鼓勵他(她)把它說出來”)2個維度作為積極教養行為的指標, 采用拒絕(4個項目, 如“如果我的孩子不來煩我, 我就不會理睬她”)作為消極教養行為指標。問卷采用5點記分(0表示完全不符合, 4表示完全符合), 平均得分越高, 則該種教養行為越多。本研究中積極和消極教養行為的Cronbach’s α系數分別為0.83和0.54。驗證性因素分析顯示二因子模型擬合良好(χ= 223.58,= 53, RMSEA = 0.06, CFI = 0.92, TLI = 0.90)。
2.2.2 青少年抑郁
采用兒童抑郁量表(Children’s Depression Inventory, CDI; Kovacs, 1992)測量青少年抑郁癥狀。該量表共包含27個項目, 要求青少年報告近兩周內的抑郁癥狀, 分別采用0、1、2記分, 計算青少年的抑郁均分, 得分越高則表明青少年的抑郁癥狀越多。該量表在正常青少年群體中應用廣泛(Zhang et al., 2015), 具有良好的測量學特性。本研究中2個時間點青少年抑郁的Cronbach’s α系數均為0.88。驗證性因素分析顯示T1抑郁(χ= 699.34,= 324, RMSEA = 0.03, CFI = 0.95, TLI = 0.95)和T2抑郁(χ= 736.72,= 324, RMSEA = 0.04, CFI = 0.96, TLI = 0.96)的單因子模型均擬合良好。
2.3 研究程序
研究經過本單位倫理委員會審核批準。首先, 項目組將問卷測查、唾液樣品采集、DNA提取與分型流程等信息告知施測學校、青少年父母及青少年本人, 獲得三方的知情同意后進行了數據收集。其次, 所有施測程序均由經過嚴格培訓的研究生主試完成。
2.3.1 問卷施測流程
首先, 以班級為單位, 采用現場問卷調查的方式測評青少年抑郁癥狀, 每個施測班級由2名經過嚴格培訓并具有豐富施測經驗的研究生擔任主試, 施測結束后問卷當場收回。其次, 現場測評后, 由青少年將裝有母親教養問卷的信封帶回家交由母親填寫, 第二天帶回交給班主任老師統一回收。
2.3.2 唾液樣本采集、DNA提取與分型流程
首先, 以班級為單位采集青少年的唾液樣本, 保證每人采集2 ml以上。采集唾液樣品后, 利用Sequenom (San Diego, CA, USA)芯片基質輔助激光解吸/電離飛行時間(MALDI-TOF)質譜平臺對rs6267、rs27072、rs1799978多態性位點進行DNA提取與基因分型。PCR引物分別為:基因:forward 5’-ACGTTGGATGTAGGTGTCAATGGCC-TCCAG-3’, reverse 5’-ACGTTGGATGTCATGGGT-GACACCAAGGAG-3’;基因:forward 5’-AG-A----AC--ACAGTGCCCCTGGG-3’, reverse 5’-AAAAAC-----GTCTAACTTCATGCTGTCTG-3’;基因:forward 5’-AGGAGCTGGAGATGGAGATGCT-3’, reverse 5’-ATGCCCATTCTTCTCTGGTTTGGC-3’。PCR反應條件:94℃ 15 min; 94℃ 20 s, 56℃ 30 s, 72℃ 1 min, 共45個循環; 最終72℃ 3 min。隨后經歷單堿基延伸反應, 基因分型采用MassARRAY Typer 3.4軟件系統進行分析。本研究所使用的檢測平臺和技術具有較高的可靠性(基因分型有效率>97%)。
2.4 數據處理與分析
首先, 對3個基因多態性進行Hardy-Weinberg平衡檢驗并考察不同基因間的關聯。其次, 對3個基因的線性基因效應和等基因效應進行檢驗, 確定線性基因編碼方式是否合適以及單個基因是否存在主導效應(Stocker et al., 2017)。該部分統計分析共檢驗三個回歸模型:分解模型(disaggregated model)包括6個基因主效應(3個基因的顯性和隱性基因編碼)、6個基因×環境交互效應(即3個基因的顯性和隱性基因編碼與教養的交互作用)、教養和控制變量的主效應(性別和早期抑郁)共15個參數估計; 線性基因效應模型(linear gene effect model)用于檢驗線性基因編碼是否合適, 將同一個基因的顯性和隱性編碼回歸權重限定相等(6個限定, 包括基因主效應和基因×環境交互效應, 允許不同基因間的效應存在差異); 等基因效應模型(equal gene effect)將3個基因的效應限定相等(4個限定, 包括基因主效應和基因×環境交互效應)。線性基因效應和等基因效應模型的優勢在于可以獲得模型限定的估計:若線性基因效應相比分解模型有顯著的解釋率變化則表明至少一個基因存在非線性效應, 則該基因不適合0、1、2的線性編碼方式, 需要調整基因編碼方式; 若等基因效應模型比線性基因效應模型有明顯的解釋率變化則表明3個基因的預測強度存在明顯差異, 至少一個基因存在不同的預測強度或預測方向, 可能存在單個基因的主導效應或基因間效應相互抵消, 不適合進行累加基因得分研究。第三, 采用相關分析考察主要研究變量間的關系。第四, 采用分層線性回歸考察多基因累加得分與母親教養行為對青少年抑郁的影響, 為避免多重共線性, 多基因累加得分和母親教養行為均進行標準化處理, 若基因×環境交互作用顯著則進一步進行簡單斜率檢驗。同時, 將性別和早期抑郁癥狀作為控制變量, 控制T1抑郁本質上考察了基因與教養對T1到T2抑郁變化的影響。第五, 采用再參數化回歸(Widaman et al., 2012)檢驗交互作用符合何種理論模型(“素質?壓力”和“不同易感性”)。最后, 為驗證結果的可靠性進行敏感性分析。敏感性分析分5個方面進行:(1)將每個基因與母親教養行為對抑郁的交互作用進行檢測, 比較單基因和多基因結果的顯著性差異, (2)將多基因累加得分劃分為低、中、高三組, 進行分組回歸分析驗證多基因×環境交互結果。(3)對基因×基因交互效應進行檢驗, 排除基因間交互效應對多基因累加效應的干擾。(4)參照既有研究文獻(Cao et al., 2018), 將消極教養的項目全部進行反向計分, 隨后將積極和消極教養(反向計分后)的項目標準化后計算均值作為母親教養敏感性得分, 重復上述分析步驟, 用以驗證結果的可靠性。為控制I型錯誤, 采用基于FDR (false discovery rate)標準的B-H法對顯著性進行統計校正(Benjamini & Hochberg, 1995)。(5)控制早期抑郁可能造成低估基因和教養對抑郁的效應, 本研究補充考察了多基因累加得分與教養對T1抑郁和T2抑郁(不控制T1抑郁)的影響。采用SPSS 23.0和R 3.3.2進行數據處理和統計分析。
3 結果
3.1 三種基因標記的基因型分布
本研究考察的、、基因型分布如表1所示, 三個位點基因型分布符合Hardy- Weinberg平衡(χs < 1.74,= 1,s > 0.05)。三個基因多態性的次要等位基因分布頻率(minor allele frequency, MAF)均大于5%。此外, 三個基因之間不存在顯著關聯(χs < 2.84,= 4,s > 0.58)。
3.2 檢驗基因標記的不同效應
在計算多基因累加得分前, 需確定各基因采用線性編碼方式是否合適、是否存在單個基因效應主導多基因累加效應的情況, 以及多個基因的效應是否會因為編碼方式而相互抵消。本研究基于多巴胺活性進行顯性和隱性編碼, 將低活性多巴胺等位基因編碼為風險/易感等位基因。參照既有研究文獻(Stocker et al., 2017), 進行線性基因和等基因效應模型檢驗。結果見表2, 分解模型的解釋率最高, 不同模型間改變量并不顯著。本研究中三個候選基因沒有顯著偏離線性和等基因效應假設。因此,后續分析中, 按照個體攜帶的低活性多巴胺等位基因數量進行線性(0、1、2)編碼, 并進行多基因累加得分計算(多基因累加得分分布:1 = 7人, 2 = 43人, 3 = 259人, 4 = 487人, 5 = 217人, 6 = 39人)。
3.3 各變量相關分析結果
主要研究變量的平均數、標準差及相關系數見表3。多基因累加得分與積極教養、消極教養行為間均不存在顯著相關, 排除了基因?環境相關的可能性。多基因累加得分越高(即多巴胺活性越低)則個體罹患抑郁的風險越大, 母親積極教養行為顯著負向預測青少年抑郁, 母親消極教養行為顯著正向預測青少年抑郁。積極和消極教養行為間存在顯著負向關聯。青少年間隔一年的抑郁癥狀間呈中等程度正相關, 表明青少年抑郁具有一定的穩定性, 配對樣本檢驗顯示, T2抑郁顯著高于T1抑郁,(1033) = –5.29,0.001。此外, 與相關結果相一致, 獨立樣本檢驗表明, 母親積極教養與抑郁均不存在顯著的性別差異(積極教養:(1038) = 1.46,0.14; T1抑郁:(1038) = –1.86,0.06; T2抑郁(1043) = –1.13,0.26), 但是母親對男生的消極教養行為顯著多于女生((1040) = –2.54,0.01)。

表1 多巴胺系統基因分布情況
注:MAF代表次要等位基因分布頻率, H-W代表Hardy-Weinberg平衡。

表2 線性和等基因效應模型比較
注:模型1包括6個基因主效應和6個基因×環境交互效應(3個基因的線性和隱性編碼)以及控制變量和教養的效應; 模型2將同一個基因的顯性和隱性編碼回歸權重限定相等(6個限定), 包括基因主效應和基因×環境交互效應;模型3將3個基因對抑郁的主效應和基因×環境交互效應設定為等同(4個限定)。
模型改變值均不顯著。
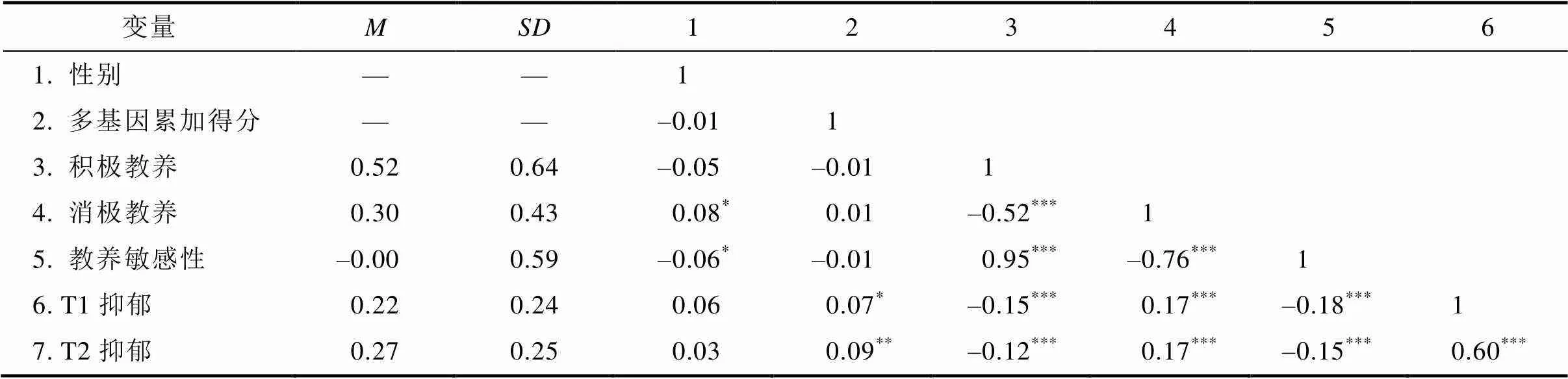
表3 描述統計與相關系數表
注:代表了平均值,代表標準差, 數字1~6分別代表了變量一列各個數字后面的變量。
< 0.05;< 0.01;< 0.001。
3.4 多基因累加得分與母親教養行為對青少年抑郁的交互作用
以T2抑郁為因變量, 控制T1抑郁和性別, 以多基因累加得分、母親教養行為以及基因×環境交互項為預測變量進行分層回歸分析。如表4所示, 多基因累加得分和母親消極教養顯著正向預測青少年抑郁, 但是母親積極教養對抑郁的預測作用不顯著; 控制T1抑郁和性別后, 多基因累加得分與母親積極教養、消極教養交互預測青少年抑郁, 且經過B-H統計校正后結果依然顯著。
進一步的簡單斜率檢驗顯示, 在控制T1抑郁和性別后, 在母親積極教養行為較低時, 具有較高多基因累加得分的青少年比具有較低多基因累加得分的青少年抑郁水平更高(= 0.03,= 2.89,= 0.004), 而在母親積極教養行為較高時, 不同多基因累加得分的青少年抑郁水平無顯著差異(= –0.002,= –0.21,= 0.84)。與此類似, 在母親消極教養行為較高時, 具有較高多基因累加得分的青少年比具有較低多基因累加得分的青少年抑郁水平更高(= 0.03,= 3.18,= 0.002), 而在母親消極教養行為較低時, 不同多基因累加得分的青少年抑郁水平無顯著差異(= –0.003,= –0.42,= 0.68) (詳見圖1)。
如表5所示, 本研究分別對積極教養和消極教養模型進行再參數化回歸, 檢驗上述交互作用符合何種理論模型(“素質?壓力”模型與“不同易感性”模型)。結果顯示, 無論是積極教養還是消極教養模型,多基因累加得分×教養的交互作用均符合“素質?壓力”模型, 詳細統計指標分析如下:(1)“素質?壓力”模型本身擬合良好(= 0.37,< 0.001); (2)在“不同易感性”模型中, 交叉點C的點估計值及其95%置信區間超出了教養行為的取值范圍(積極教養:–4.22 ~ 1.52; 消極教養:–1.31 ~ 3.76; 教養敏感性:–4.16 ~ 1.60), 這表明基因×環境交互模式不符合“不同易感性”假設。(3)檢驗顯示盡管“不同易感性”模型比“素質?壓力”模型增加一個參數(C)估計, 但“不同易感性”模型的解釋率并沒有顯著高于“素質?壓力”模型(Δs= 0.00,s > 0.05)。(4)“素質?壓力”模型的AIC和BIC指標均小于“不同易感性”模型。綜上, 各參數均表明, “素質?壓力”模型對數據的擬合優于“不同易感性”模型, 因此本研究中多巴胺系統多基因累加得分與母親教養行為對青少年抑郁的交互作用更加符合“素質?壓力”模型。
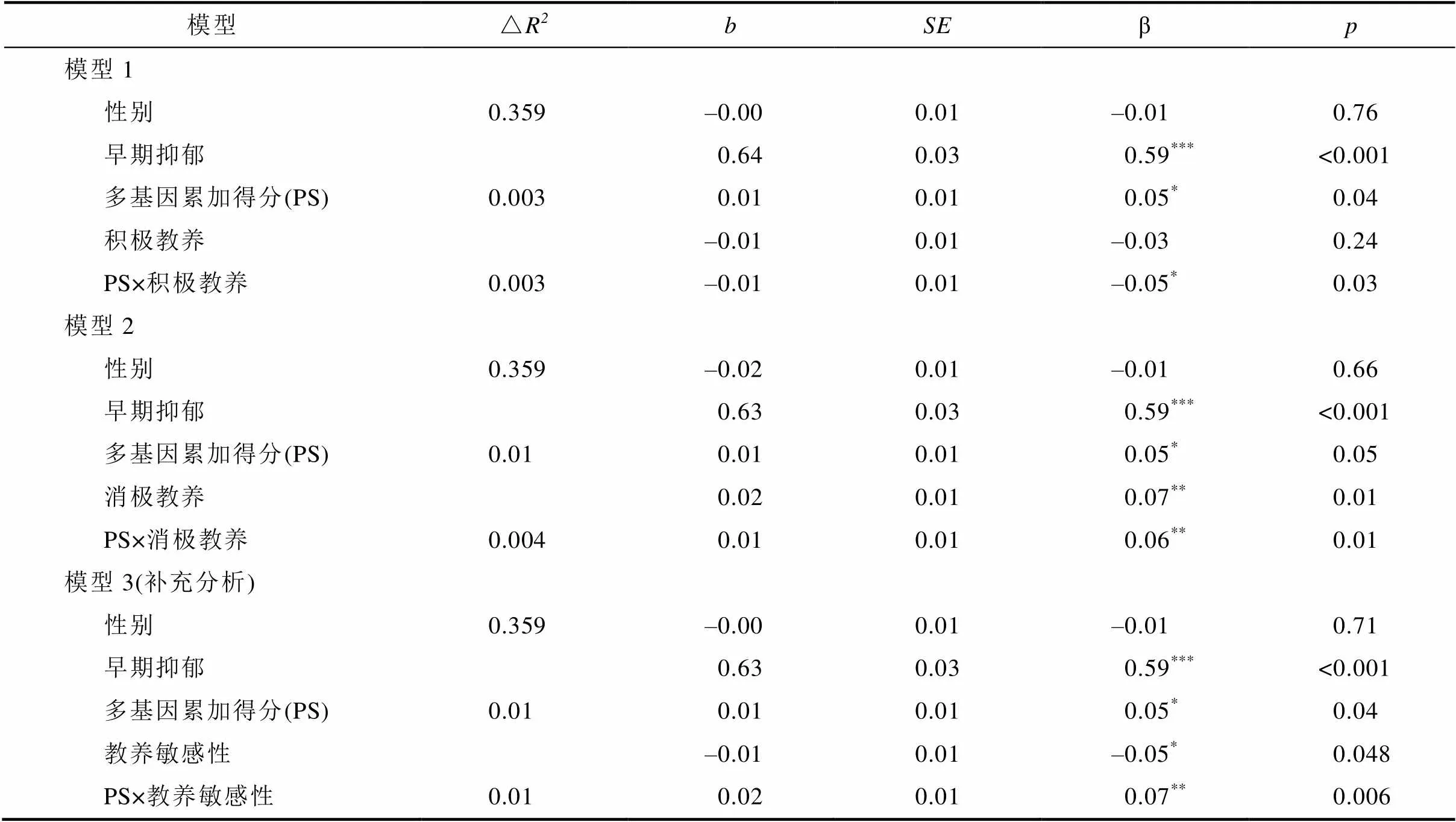
表4 多基因累加得分與母親教養行為對青少年抑郁的影響
注:< 0.05;< 0.01;< 0.001。

圖1 多基因累加得分與母親教養行為對青少年抑郁的影響
3.5 敏感性分析
為驗證上述研究結果的可靠性, 本研究進行了敏感性分析。首先, 對任何單基因與母親積極教養、消極教養行為對抑郁的交互作用進行回歸分析。結果顯示在B-H校正后, 除基因與積極教養行為的交互作用顯著外(= –0.05,= –2.78,= 0.006), 任何單基因與教養的交互作用均不顯著(|s < 0.04, ||s = –2.27,s > 0.05)。這也表明多基因累加得分研究在提高遺傳解釋率和顯著性方面比單基因研究更具優勢。
其次, 前述分析將多基因累加得分作為連續變量進行分析, 由于攜帶1、2和6個低多巴胺活性等位基因的人數較少, 可能影響上述交互作用結果的可靠性。為進一步驗證上述結果, 本研究對多基因累加得分重新編碼進行分組回歸檢驗。將多基因累加得分為1、2、3的青少年編碼為低分組, 將多基因累加得分為4的青少年編碼為中等得分組, 將多基因累加得分為5、6的青少年編碼為高分組。分組回歸分析結果與前述結果相一致, 相比低分組和中等得分組, 高分組的青少年對母親積極教養(= –0.04,= –2.33,= 0.02)和消極教養更加敏感(= 0.04,= 2.65,= 0.01) (詳見圖2)。
第三, 為了排除基因×基因交互效應的影響, 本研究對G × G × G × E效應進行了分析。需要指出的是, 采用線性基因編碼(0、1、2)的方式會導致每個基因型組合單元格人數限制無法滿足統計分析條件。本研究替代性的采用了二分編碼方式(: 0 = GG, 1 = T;: 0 = T, 1 = CC;: 0 = AA, 1 = G)進行分析, 結果顯示任何G × G, G × G × G, G × G × E, G × G × G × E交互效應均不顯著(|≤ 0.22, |≤ 0.90,≥ 0.37), 在一定程度上排除了潛在基因×基因效應的干擾。
第四, 參照既有研究文獻(Cao et al., 2018), 將消極教養和積極教養合并后的母親教養敏感性總分作為環境指標, 重復上述統計分析, 結果與單獨進行積極和消極教養行為的模式一致, 只是交互項的顯著性和效應量更大(參見表2、表4和表5的補充分析部分)。

表5 多基因累加分與父母教養行為對青少年抑郁交互作用的再參數化回歸模型檢驗
注:模型=+×性別 +×早期抑郁 +× (–) +× (–) × 多基因累加得分 +; CI代表置信區間;a vs. b代表“不同易感性”與“素質?壓力”模型之間的檢驗, 以分析減少一個估計參數時模型的Δ是否存在顯著變化; a代表該參數限定在指定數值上, 如= 1.52、–1.31、1.60表示將交叉點固定在積極教養的最大值1.52、消極教養的最小值–1.31處、母親教養敏感性(積極和消極教養合并分)的最大值1.60處。
< 0.05;< 0.01;< 0.001。
第五, 補充考察了多基因累加得分和教養對T1抑郁的同時效應, 以及對T2抑郁的縱向效應(不控制T1抑郁)。結果顯示, 多基因與教養的主效應和顯著性提高。但是, 除基因與消極教養對T2抑郁的交互作用顯著外, 其他基因×環境均不顯著(見表6和表7)。

圖2 多基因累加得分與母親教養行為對青少年抑郁影響的敏感性分析

表6 多基因累加得分與母親教養行為對青少年T1抑郁的影響
注:< 0.05;< 0.01;< 0.001。
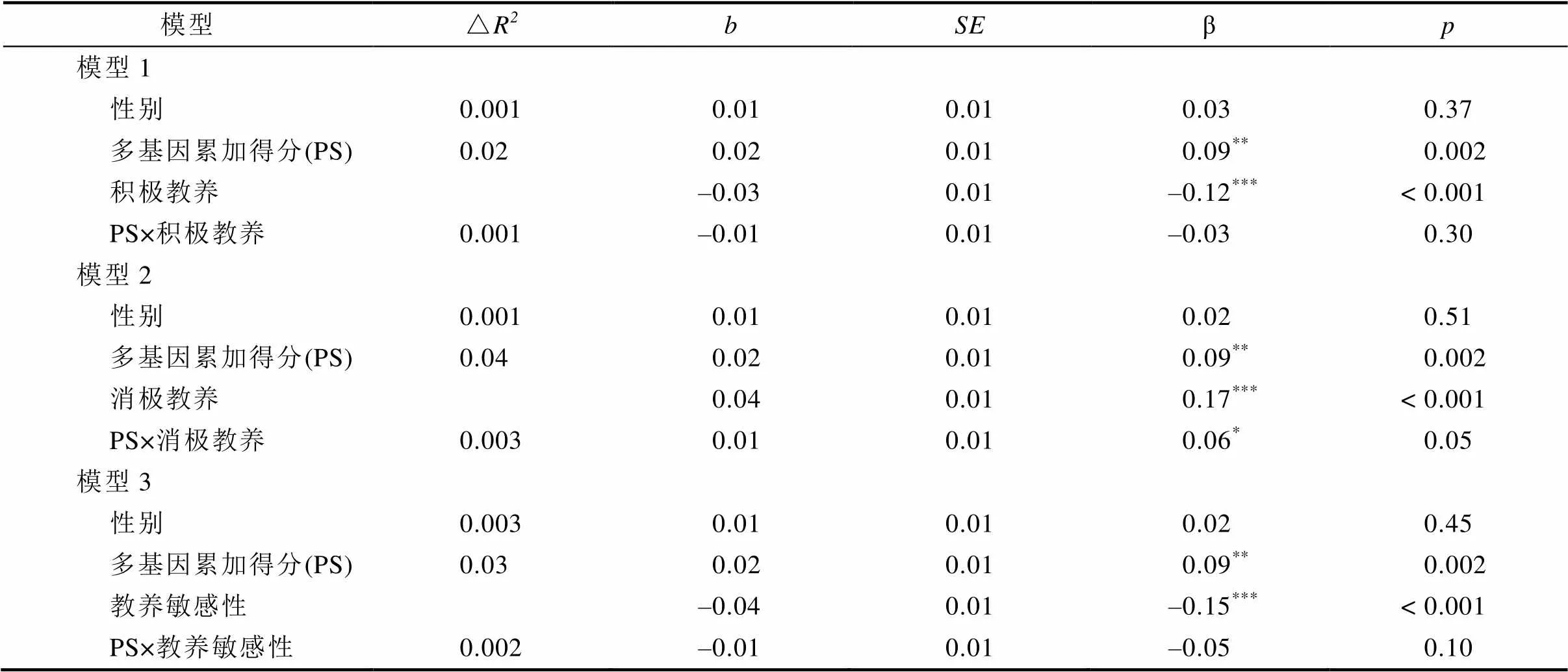
表7 多基因累加得分與母親教養行為對青少年T2抑郁的影響(未控制早期抑郁)
注:< 0.05;< 0.01;< 0.001。
4 討論
本研究采用多基因累加得分研究范式, 以三種多巴胺系統基因(基因、基因和基因)為遺傳指標, 考察了母親教養行為與多基因累加得分對青少年抑郁的影響。結果顯示控制早期抑郁后, 多基因累加得分顯著正向預測青少年抑郁; 多巴胺系統多基因累加得分與母親積極、消極教養均交互預測青少年抑郁, 其交互模式均符合“素質?壓力”模型。具體表現為, 在低積極或高消極教養環境中, 多基因累加得分較高的青少年比多基因累加得分較低的青少年抑郁水平更高, 但是在高積極或低消極教養環境中, 攜帶不同多基因累加得分的青少年抑郁水平不存在顯著差異。
與以往研究(e.g., Wang et al., 2016)相一致, 本研究發現母親消極教養行為是導致青少年抑郁的風險因素。但是, 母親積極教養對青少年抑郁的影響未達到顯著性水平。這可能是由于母親積極教養和消極教養行為對青少年抑郁的影響存在差異。一方面, 積極和消極教養行為對青少年抑郁的作用程度存在差異, 如Dallaire等(2006)的研究發現, 消極教養行為對兒童青少年抑郁的預測力更強。另一方面, Smokowski, Bacallao, Cotter和Evans (2015)研究進一步揭示, 不同類型的教養行為對青少年適應結果的預測作用存在領域特殊性, 積極教養可能與積極心理特征的關聯更為密切, 而對消極適應結果的影響較小, 而消極教養行為則是焦慮、抑郁等消極適應結果的關鍵風險因素。更重要的是, 積極和消極教養對青少年抑郁的作用路徑可能也存在差異, 如Luebbe和Bell (2014)的研究發現消極家庭氛圍(包括消極教養行為)和積極家庭氛圍(包括積極教養行為)分別通過消極情緒性和積極情緒性間接影響青少年抑郁。由此, 未來研究可以增加積極適應結果考察教養與適應結果間的領域特殊性問題。
本研究發現個體攜帶的多基因累加得分越高(多巴胺系統功能越低)則其抑郁水平越高。這與單胺缺陷假說相一致, 多巴胺系統功能的缺陷會增加個體罹患抑郁的風險。更重要的是, 本研究為抑郁的多基因遺傳結構提供了支持。但是, 本研究中多基因聯合效應的解釋率仍然為1%左右。這可能提示了抑郁具有復雜的產生和發展機制, 本研究所選取的多巴胺系統多基因累加得分也僅能反映其致病因素的冰山一角, 研究者應該更加慎重地看待這一結果。此外, 研究者應該深入探索多基因影響抑郁的作用機制, 如“多基因累加得分×環境”研究等。
從交互作用上來看, 在控制早期抑郁后, 母親教養行為與多巴胺系統多基因累加得分交互預測青少年后繼抑郁, 并且進一步的分析發現, 只有在消極環境下(包括低積極教養和高消極教養), 多基因累加得分較高的青少年比得分較低的個體具有更高的抑郁風險。再參數化回歸分析進一步顯示這一交互作用模式更符合“素質?壓力”模型。與此結果相一致, 前述關于腹側紋狀體活性的研究顯示, 個體攜帶的低活性多巴胺系統等位基因越多, 則在面對外界刺激時表現出更低的紋狀體反應性(Nikolova et al., 2011)。有關獎賞和懲罰敏感性的研究發現, 對獎賞刺激的敏感性與紋狀體激活水平呈正向關聯, 而對懲罰的敏感性與紋狀體激活程度呈負向關聯(Kim, Yoon, Kim, & Hamann, 2015)。這些研究結果提示, 攜帶低活性多巴胺等位基因數量越多的個體具有較低的紋狀體激活水平, 對社會獎賞刺激(如積極的教養環境)的敏感性較低從而無法在積極的環境中獲益, 但是其對消極環境刺激的敏感性較高從而增加其抑郁的風險。需要指出的是, 這一交互作用模式可能也僅僅描繪了青少年早期階段多基因×環境的交互作用, 當前研究者更傾向于采用動態的視角看待“素質?壓力”和“不同易感性”模型(王美萍, 2015)。譬如, 研究發現多基因累加得分(基因和基因)與家庭環境質量對抑郁的交互作用模式在青少年早期(15歲前)符合“不同易感性”假說, 而在青少年中晚期則符合“素質?壓力”模型(Dalton, Hammen, Najman, & Brennan, 2014)。由此, 未來研究應該補充其他年齡階段的被試或采用追蹤設計, 深入探索多基因×環境交互作用模式是否具有發展動態性。
雖然本研究中多基因累加得分及其與環境的交互作用解釋率仍然很低(1%), 但敏感性分析顯示, 與單基因相比, 多基因累加得分在提高遺傳解釋率和顯著性方面仍然具有一定的優勢。值得指出的是, 這并不意味著研究者可以為擴大遺傳效應而隨意增加基因位點數量來計算多基因累加得分。事實上, 即便累加了眾多SNP基因位點, 也無法解釋抑郁全部的遺傳機制, 甚至對遺傳率的提升甚微。如Peyrot等(2014)采用全基因組關聯掃描(GWAS)方法, 分別將150~32870個SNP位點納入多基因累加得分計算, 其多基因累加得分及其與童年期虐待的交互作用解釋率僅為0.09%~0.90%。但是, 較小的遺傳解釋率并不意味著它是可以忽視的(Evans, 1985)。因此, 研究者不能盲目追求解釋率的提升而濫用遺傳指標。由此, 如何選擇多基因累加指標成為研究者關注的重點, 伴隨著越來越多新功能性基因標記的發現, 這一問題變得尤為突出。對此, Vrshek-Schallhorn等(2015)指出為了避免數據驅動和濫用的風險, 在進行多基因累加效應研究時最好選擇同一神經遞質系統的基因而不是跨系統的基因, 這樣有助于探索多基因與內表型之間的關聯, 進而確定抑郁發生的內部作用機制。
此外, 多基因累加得分解釋率的限制使得研究者對探索“缺失遺傳率”表現出極大的興趣。Manolio等(2009)指出遺傳率缺失的原因可能包括:第一, 大量具有極小效應的基因變體尚未被發現。因此, 采用GWAS和GWEIS (genome-wide by environment interaction studies)研究方法探索潛在的功能性基因及潛在的基因×環境交互作用仍然是未來研究的重點方向。第二, 研究者應該跳出“常見疾病, 常見變體(common disease, common variant)”的固有假設, 探索遺傳效應較大的罕見基因變體(在人群中的比例低于5%的基因變體)。第三, 現有研究很少測量DNA表達機制, 限制了其對表型的遺傳解釋率。甲基化、組蛋白修飾等表觀遺傳機制可以在不改變DNA結構的情況下影響基因?表型間的關聯。因此, 考察多基因累加得分與基因表達水平的變化, 可用于探尋其遺傳解釋率的缺失并更進一步剖析遺傳因素的潛在生物機制。第四, 多基因累加得分未能檢測基因×基因交互作用。在本研究中, 受單元格人數限制, 采用二分編碼方式對基因×基因交互效應進行了檢驗并在一定程度上證實了多基因累加的可靠性。但是, 在未來多基因累加研究中尚需對不同基因間的交互效應保持謹慎。正如Plomin, DeFries, McClearn和McGuffin (2001)指出多基因遺傳模型除了需要考慮不同等位基因效應間的累加效應外, 還需要考慮不同基因座的效應彼此間有相互作用的可能性。
本研究采用縱向設計控制早期抑郁, 考察多基因累加得分與母親教養對青少年抑郁的影響。采用這一設計的主要原因是青少年抑郁與母親教養間通常存在雙向關系(Hamza & Willoughby, 2011), 即母親教養不僅影響青少年抑郁, 而且也會受到青少年抑郁情緒的影響。由此, 如果不控制青少年早期抑郁, 教養行為對青少年后繼抑郁的影響本質上可能也混雜了早期抑郁對后繼抑郁的影響。因此, 控制早期抑郁有助于降低母親教養行為是抑郁癥狀的結果而不是原因的可能性(Chen, Li, & McGue, 2013)。雖然控制早期抑郁會帶來一些優勢, 但也可能會造成基因和環境的效應量和顯著性降低。此外, 綜合考慮控制早期抑郁與不控制早期抑郁的結果, 在理解本研究結果時需要注意的是, 多基因累加得分與母親教養的交互作用不能預測T1和T2時間點上絕對水平的抑郁癥狀, 本質上是預測了T1到T2抑郁的變化。在Petersen等(2012)的研究中也發現了類似的結果模式, 即G × E不能預測基線水平的焦慮抑郁癥狀, 但能夠顯著預測焦慮抑郁的發展變化, 尤其是能夠解釋抑郁隨年齡的變化趨勢。由此, 即使未能發現遺傳與環境交互效應對抑郁水平的顯著預測作用, 也不意味著G × E與抑郁發展無關。未來研究應該更深入的考察遺傳與環境對抑郁及其發展變化的影響以更深入地揭示此問題。
需要注意的是, 本研究也具有一些局限性。第一, 本研究缺乏重復驗證性樣本。近年來, 伴隨著基因×環境交互作用研究分歧的增多, 越來越多的研究者對遺傳研究的可重復性問題提出了質疑, 并且指出應該采用外部驗證樣本進行重復檢驗(Christ, Schwartz, Stoltenberg, Brauer, & Savolainen, 2018; Dick et al., 2015)。但是本研究未能具有相同測量工具及相似被試特征的外部驗證樣本, 其結果的可重復性有待未來研究進行驗證。第二, 多巴胺系統基因眾多, 本研究僅采用了三個基因多態性考察了多基因遺傳基礎, 未來研究尚需要考察其他候選基因多態性以豐富該領域的研究。第三, 本研究中消極教養的信度指標較低, 這可能與消極教養的測量題目(4個)較少有關。雖然本研究的驗證性分析也保證了其結構效度, 但仍需要謹慎的看待本研究結果。此外, 本研究僅考察了母親積極和消極教養行為, 未對父親教養行為進行測量。長期以來, 在分子遺傳研究領域, 研究者往往忽視了父親、母親教養行為的差異性及相互影響。據我們所知, 目前僅有一項研究比較了多基因累加得分與父親、母親教養行為交互作用的不同但未發現顯著差異(Stocker et al., 2017)。但是, 也有研究顯示父親和母親教養行為交互影響青少年抑郁(Quach, Epstein, Riley, Falconier, & Fang, 2015)。由此, 未來研究應該考察父親、母親教養行為與遺傳因素交互作用的差異, 并且關注父親和母親教養行為間的相互影響。最后, 本研究被試來自城市常態群體青少年, 該研究結果能否適用于臨床樣本或重性抑郁以及社會經濟地位較低的青少年群體尚需未來研究的驗證。
Aliev, F., Latendresse, S. J., Bacanu, S. A., Neale, M. C., & Dick, D. M. (2014). Testing for measured gene-environment interaction: Problems with the use of cross-product terms and a regression model reparameterization solution.(2), 165–181.
Andersen, S. L., & Teicher, M. H. (2008). Stress, sensitive periods and maturational events in adolescent depression.(4), 183–191.
Beauchaine, T. P., Gatzke-Kopp, L., & Mead, H. K. (2007). Polyvagal theory and developmental psychopathology: Emotion dysregulation and conduct problems from preschool to adolescence.(2), 174–184.
Belmaker, R. H., & Agam, G. (2008). Major depressive disorder.(1), 55–68.
Belsky, J., & Pluess, M. (2009). Beyond diathesis stress: Differential susceptibility to environmental influences.(6), 885–908.
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery rate: A practical and powerful approach to multiple testing.,(1), 289–300.
Bischoff, A. R., Pokhvisneva, I., Léger, é., Gaudreau, H., Steiner, M., Kennedy, J. L., ... Silveira, P. P. (2017). Dynamic interaction between fetal adversity and a genetic score reflecting dopamine function on developmental outcomes at 36 months.(5), e0177344.
Cao, C., Rijlaarsdam, J., van der Voort, A., Ji, L. Q., Zhang, W. X., & Bakermans-Kranenburg, M. J. (2018). Associations between dopamine D2 receptor (DRD2) gene, maternal positive parenting and trajectories of depressive symptoms from early to mid-adolescence.(2), 365–379.
Cao, Y. M., Lin, X. N., Chen, L., Ji, L. Q., & Zhang, W. X. (2018). The catechol-O-methyltransferase and dopamine transporter genes moderated the impact of peer relationships on adolescent depressive symptoms: A gene-gene- environment study.(11), 2468–2480.
Cao, Y. M., Wang, M. P., Cao, C., Ji, L. Q., & Zhang, W. X. (2017). The interaction between dopamine D2 receptor gene TaqIA polymorphim and peer victimization on early adolescent depression.(1), 28–39.
[曹衍淼, 王美萍, 曹叢, 紀林芹, 張文新. (2017). DRD2基因TaqIA多態性與同伴侵害對青少年早期抑郁的交互作用., 28–39.]
Chen, J., Li, X. Y., & McGue, M. (2013). The interacting effect of the BDNF Val66Met polymorphism and stressful life events on adolescent depression is not an artifact of gene–environment correlation: Evidence from a longitudinal twin study.(10), 1066–1073.
Chen, X. Y., Bian, Y. F., Xin, T., Wang, L., & Silbereisen, R. K. (2010). Perceived social change and childrearing attitudes in China.(4), 260–270.
Christ, C. C., Schwartz, J. A., Stoltenberg, S. F., Brauer, J. R., & Savolainen, J. (2018). The effect of MAOA and stress sensitivity on crime and delinquency: A replication study.(3), 336–353.
Cole, D. A., Tram, J. M., Martin, J. M., Hoffman, K. B., Ruiz, M. D., Jacquez, F. M., & Maschman, T. L. (2002). Individual differences in the emergence of depressive symptoms in children and adolescents: A longitudinal investigation of parent and child reports.(1), 156–165.
Coley, R. L., Sims, J., & Carrano, J. (2017). Environmental risks outweigh dopaminergic genetic risks for alcohol use and abuse from adolescence through early adulthood., 106–118.
Costa, A., Riedel, M., Müller, U., M?ller, H. J., & Ettinger, U. (2011). Relationship between SLC6A3 genotype and striatal dopamine transporter availability: A meta-analysis of human single photon emission computed tomography studies.(10), 998–1005.
Dallaire, D. H., Pineda, A. Q., Cole, D. A., Ciesla, J. A., Jacquez, F., LaGrange, B., & Bruce, A. E. (2006). Relation of positive and negative parenting to children's depressive symptoms.(2), 313–322.
Dalton, E. D., Hammen, C. L., Najman, J. M., & Brennan, P. (2014). Genetic susceptibility to family environment: BDNF Val66met and 5-HTTLPR influence depressive symptoms.(6), 947–956.
Davies, P., Cicchetti, D., & Hentges, R. F. (2015). Maternal unresponsiveness and child disruptive problems: The interplayof uninhibited temperament and dopamine transporter genes.(1), 63–79.
Dick, D. M., Agrawal, A., Keller, M. C., Adkins, A., Aliev, F., Monroe, S., ... Sher, K. J. (2015). Candidate gene– environment interaction research: Reflections and recommendations.(1), 37–59.
Doehring, A., Kirchhof, A., & L?tsch, J. (2009). Genetic diagnostics of functional variants of the human dopamine D2 receptor gene.(5), 259–268.
Duncan, L. E., & Keller, M. C. (2011). A critical review of the first 10 years of candidate gene-by-environment interaction research in psychiatry.(10), 1041–1049.
[4] Adam Entous, “In Asia, Tone Lightens On Sea Disputes,” Wall Street Journal, October 13, 2010.
Dunlop, B. W., & Nemeroff, C. B. (2007). The role of dopamine in the pathophysiology of depression.(3), 327–337.
Dunn, E. C., Uddin, M., Subramanian, S. V., Smoller, J. W., Galea, S., & Koenen, K. C. (2011). Research review: Gene-environment interaction research in youth depression – A systematic review with recommendations for future research.(12), 1223–1238.
Evans, M. G. (1985). A Monte Carlo study of the effects of correlated method variance in moderated multiple regression analysis.(3), 305–323.
Ferro, M. A., Gorter, J. W., & Boyle, M. H. (2015). Trajectories of depressive symptoms during the transition to young adulthood: The role of chronic illness., 594–601.
Giros, B., Jaber, M., Jones, S. R., Wightman, R. M., & Caron, M. G. (1996). Hyperlocomotion and indifference to cocaine and amphetamine in mice lacking the dopamine transporter., 606–612.
Hamza, C. A., & Willoughby, T. (2011). Perceived parental monitoring, adolescent disclosure, and adolescent depressive symptoms: A longitudinal examination.(7), 902–915.
Kim, S. H., Yoon, H., Kim, H., & Hamann, S. (2015). Individual differences in sensitivity to reward and punishment and neural activity during reward and avoidance learning.(9), 1219–1227.
Kovacs, M. (1992).. Toronto, Canada: Multi–Health Systems Inc.
Lee, S. G., Joo, Y., Kim, B., Chung, S., Kim, H. L., Lee, I., ... Song, K. (2005). Association of Ala72Ser polymorphism with COMT enzyme activity and the risk of schizophrenia in Koreans.(4), 319–328.
Lewis, D. A., Melchitzky, D. S., Sesack, S. R., Whitehead, R. E., & Sampson, A. (2001). Dopamine transporter immunoreactivity in monkey cerebral cortex: Regional, laminar, and ultrastructural localization.(1), 119–136.
Lin, C. H., Chaudhuri, K. R., Fan, J. Y., Ko, C. I., Rizos, A., Chang, C. W., ... Wu, Y. R. (2017). Depression and catechol-O-methyltransferase (COMT) genetic variants are associated with pain in Parkinson’s disease., 6306.
Luebbe, A. M., & Bell, D. J. (2014). Positive and negative family emotional climate differentially predict youth anxiety and depression via distinct affective pathways.(6), 897–911.
Manolio, T. A., Collins, F. S., Cox, N. J., Goldstein, D. B., Hindorff, L. A., Hunter, D. J., ... Cho, J. H. (2009). Finding the missing heritability of complex diseases.(7265), 747–753.
Matsumoto, M., Weickert, C. S., Beltaifa, S., Kolachana, B., Chen, J., Hyde, T. M., ... Kleinman, J. E. (2003). Catechol O-methyltransferase (COMT) mRNA expression in the dorsolateral prefrontal cortex of patients with schizophrenia.(8), 1521–1530.
Monroe, S. M., & Simons, A. D. (1991). Diathesis-stress theories in the context of life stress research: Implications for the depressive disorders.(3), 406–425.
Natsuaki, M. N., Biehl, M. C., & Ge, X. (2009). Trajectories of depressed mood from early adolescence to young adulthood: The effects of pubertal timing and adolescent dating.(1), 47–74.
Nikolova, Y. S., Ferrell, R. E., Manuck, S. B., & Hariri, A. R. (2011). Multilocus genetic profile for dopamine signaling predicts ventral striatum reactivity.(9), 1940–1947.
Nivard, M. G., Dolan, C. V., Kendler, K. S., Kan, K. J., Willemsen, G., van Beijsterveldt, C. E. M., ... Boomsma, D. I. (2015). Stability in symptoms of anxiety and depression as a function of genotype and environment: A longitudinal twin study from ages 3 to 63 years.(5), 1039–1049.
Noble, E. P., Gottschalk, L. A., Fallon, J. H., Ritchie, T. L., & Wu, J. C. (1997). D2 dopamine receptor polymorphism and brain regional glucose metabolism.(2), 162–166.
Olino, T. M., McMakin, D. L., Nicely, T. A., Forbes, E. E., Dahl, R. E., & Silk, J. S. (2016). Maternal depression, parenting, and youth depressive symptoms: Mediation and moderation in a short-term longitudinal study.(3), 279–290.
Opmeer, E. M., Kortekaas, R., & Aleman, A. (2010). Depression and the role of genes involved in dopamine metabolism and signalling.(2), 112–133.
Pearson-Fuhrhop, K. M., Dunn, E. C., Mortero, S., Devan, W. J., Falcone, G. J., Lee, P., ... Cramer, S. C. (2014). Dopamine genetic risk score predicts depressive symptoms in healthy adults and adults with depression.(5), e93772.
Petersen, I. T., Bates, J. E., Goodnight, J. A., Dodge, K. A., Lansford, J. E., Pettit, G. S., ... Dick, D. M. (2012). Interaction between serotonin transporter polymorphism (5-HTTLPR) and stressful life events in adolescents' trajectories of anxious/depressed symptoms.(5), 1463–1475.
Peyrot, W. J., Milaneschi, Y., Abdellaoui, A., Sullivan, P. F., Hottenga, J. J., Boomsma, D. I., & Penninx, B. W. J. H. (2014). Effect of polygenic risk scores on depression in childhood trauma.(2), 113–119.
Pinsonneault, J. K., Han, D. D., Burdick, K. E., Kataki, M., Bertolino, A., Malhotra, A. K., ... Sadee, W. (2011). Dopamine transporter gene variant affecting expression in human brain is associated with bipolar disorder.(8), 1644–1655.
Plomin, R., DeFries, J. C., McClearn, G. E., & McGuffin, P. (2001).(Wen, N., Wang, X. H., Yang, Y. P., & Liu, X. L. Trans.). Shanghai, China: Huadong Normal University Press. (Original work published 2001).
[Plomin, R., DeFries, J. C., McClearn, G. E., & McGuffin, P. (2001)..(溫暖, 王小惠, 楊彥平, 劉曉陵譯). 上海: 華東師范大學出版社.]
Quach, A. S., Epstein, N. B., Riley, P. J., Falconier, M. K., & Fang, X. (2015). Effects of parental warmth and academic pressure on anxiety and depression symptoms in Chinese adolescents.(1), 106–116.
Roisman, G. I., Newman, D. A., Fraley, R. C., Haltigan, J. D., Groh, A. M., & Haydon, K. C. (2012). Distinguishing differential susceptibility from diathesis–stress: Recommendations for evaluating interaction effects.(2), 389–409.
Smokowski, P. R., Bacallao, M. L., Cotter, K. L., & Evans, C. B. R. (2015). The effects of positive and negative parenting practices on adolescent mental health outcomes in a multicultural sample of rural youth.(3), 333–345.
Stocker, C. M., Masarik, A. S., Widaman, K. F., Reeb, B. T., Boardman, J. D., Smolen, A., ... Conger, K. J. (2017). Parenting and adolescents’ psychological adjustment: Longitudinal moderation by adolescents’ genetic sensitivity.(4), 1289–1304.
Vrshek-Schallhorn, S., Stroud, C. B., Mineka, S., Zinbarg, R. E., Adam, E. K., Redei, E. E., ... Craske, M. G. (2015). Additive genetic risk from five serotonin system polymorphisms interacts with interpersonal stress to predict depression.(4), 776–790.
Wang, C. X., Xia, Y., Li, W. Z., Wilson, S. M., Bush, K., & Peterson, G. (2016). Parenting behaviors, adolescent depressive symptoms, and problem behavior: The role of self-esteem and school adjustment difficulties among Chinese adolescents.(4), 520–542.
Wang, M. P. (2015). Theory models on gene-environment interaction., 1852–1858.
[王美萍. (2015). 基因–環境交互作用理論模型及其檢驗方法., 1852–1858.]
Widaman, K. F., Helm, J. L., Castro-Schilo, L., Pluess, M., Stallings, M. C., & Belsky, J. (2012). Distinguishing ordinal and disordinal interactions.(4), 615–622.
Xia, L. W., & Yao, S. Q. (2015). The involvement of genes in adolescent depression: A systematic review., 9, 329.
Xing, Q. H., Qian, X. Q., Li, H. F., Wong, S. M., Wu, S. N., Feng, G. Y., ... He, L. (2007). The relationship between the therapeutic response to risperidone and the dopamine D2 receptor polymorphism in Chinese schizophrenia patients.(5), 631–637.
Zhang, J. P., & Malhotra, A. K. (2011). Pharmacogenetics and antipsychotics: therapeutic efficacy and side effects prediction.(1), 9-37.
Zhang, W. X., Cao, Y. M., Wang, M. P., Ji, L. Q., Chen, L., & Deater-Deckard, K. (2015). The dopamine D2 receptor Polymorphism (DRD2 TaqIA) interacts with maternal parenting in predicting early adolescent depressive symptoms: Evidence of differential susceptibility and age differences.(7), 1428–1440.
The influence of dopaminergic genetic variants and maternal parenting on adolescent depressive symptoms: A multilocus genetic study
CAO Yanmiao; ZHANG Wenxin
(School of Psychology, Shandong Normal University, Jinan 250014, China)
For decades, there is increasing evidence for the importance of single-gene by environment interactions (G × E) in understanding the etiology of depression. However, several concerns have been raised about the ignoring the polygenic traits of depression when conducting G × E research using single loci. Within this context, the multilocus genetic profile score (MGPS) have recently emerged as an approach of capturing polygenic nature across multiple genes. In line with the monoamine deficiency hypothesis, recent research has begun to show that the combined effects of multiple dopaminergic genetic variants are stronger than the influence of any single gene examined in isolation. Additionally, genes related to the functioning of the dopaminergic system, which coordinates individual’s response to stress. However, existing G × E research has largely focused on adverse family environments (i.e., maltreatment, maternal unresponsiveness) and to a lesser extent on positive environment, such as positive parenting. Therefore, the present study aimed to examine the interaction between dopaminergic genetic variants and maternal parenting on adolescent depressive symptoms, by adopting the approach of multilocus genetic profile score.
Participants were 1052 mother-offspring (adolescents mean age 12.31 ± 0.37 years old at the first time point, 50.2% females) dyads recruited from the community. Youth completed assessments twice with an interval of one year. Saliva samples, self-reported depressive symptoms and mother-reported parenting were collected. All measures showed good reliability. Genotyping in three dopaminergic genes were performed for each participant in real time with MassARRAY RT software version 3.0.0.4 and analyzed using the MassARRAY Typer software version 3.4 (Sequenom). To examine whether multilocus genetic profile score moderates the effects of parenting on adolescent depressive symptoms and whether this potential moderating effect act in a diathesis–stress or differential susceptibility manner, hierarchical regression analyses were conducted. We also tested above questions by recoding into categorical variables and re-conducted analyses.
The results found that multilocus genetic profile score was a significant risk factor of depression, with higher dopamine genetic risk scores (indicating lower dopaminergic neurotransmission) predicting higher levels of depression. After controlling for gender and prior depressive symptoms, the G × E effect with positive and negative parenting were also significant, suggesting that G × E interaction significantly predicted change in depression level between Time 1 and Time 2. Specifically, adolescents with higher MGPS exhibited higher risk for depression when encountered with lower levels of positive parenting and higher levels of negative parenting, compared to their counterparts with lower MGPS. The results support the diathesis-stress model and highlight the complex ways that genes and environment interact to influence development.
These finding underscores complex polygenic underpinnings of depression and lends support for the mulitlocus genetic profile scores-environment interactions implicated in the etiology of depressive symptoms.
adolescent depression, maternal parenting, dopamine, multilocus genetic profile score, gene- environment interaction
2019-01-31
* 國家自然科學基金項目(31671156; 31900776)和中國博士后科學基金(2017M622249)資助。
B844; B845
張文新, E-mail: zhangwenxin@sdnu.edu.cn
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
核科學與工程(2021年4期)2022-01-12 06:30:26
瘋狂英語·新讀寫(2021年7期)2021-10-13 06:35:44
今日農業(2020年19期)2020-12-14 14:16:52
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
中學物理·高中(2016年12期)2017-04-22 11:53:03
光學精密工程(2016年6期)2016-11-07 09:07:19
中國火炬(2014年4期)2014-07-24 14:22:19
中國火炬(2013年1期)2013-07-24 14:20:18
