唐古特白刺果實花色苷改善高脂飲食誘導的小鼠非酒精性脂肪肝損傷
2019-10-30 05:32:42李翠芹
食品科學 2019年19期
李 冰,劉 康,張 錦,李翠芹,2,*
(1.陜西師范大學 西北瀕危藥材資源開發國家工程實驗室,藥用植物資源與天然藥物化學教育部重點實驗室,陜西 西安 710119;2.陜西師范大學生命科學學院,陜西 西安 710119)
非酒精性脂肪肝(nonalcoholic fatty liver disease,NAFLD)是代謝綜合征在肝臟中的表現,是臨床上十分常見的肝臟疾病[1]。NAFLD很容易發展為非酒精性脂肪性肝炎、肝纖維化和肝硬化[2]。長期攝入高脂飲食會導致肥胖和NAFLD等疾病[3]。來源于天然植物中的黃酮類成分可預防或輔助治療高脂飲食誘發的肥胖和肝損傷[4]。花色苷是一種常見的黃酮類天然色素和膳食營養因子,廣泛分布于水果、蔬菜和農作物中[5]。由于具有較高的藥用價值和亮麗的色澤,花色苷已經作為一種功能性的天然色素和營養物質廣泛應用于食品和藥品等領域[6-7]。
唐古特白刺(Nitraria tangutorun Bobr.)為蒺藜科(Zygophyllaceae)白刺屬(Nitraria)植物,廣泛分布于青藏高原沙漠地區,為中國特有品種[8]。《本草拾遺》記載“東廧(白刺古名),味甘平、無毒,益氣輕身(減肥),久服不饑,堅筋骨”,唐古特白刺果實經常被當地人用來治療脾胃虛弱、消化不良、神經衰弱和感冒[9]。現代研究表明,唐古特白刺果實富含花色苷、多酚和黃酮等多種生物活性成分[10]。唐古特白刺是花色苷的優質來源[11],然而有關唐古特白刺果實花色苷防治高脂飲食誘發的肥胖及NAFLD損傷的研究鮮見報道。
本研究提取唐古特白刺果實花色苷,經過長期高脂飲食建立小鼠NAFLD模型。在此基礎上研究唐古特白刺果實花色苷對NAFLD的預防與緩解作用,及其作用機制。以期為唐古特白刺果實花色苷防治NAFLD提供數據支撐,并對唐古特白刺果實的資源開發提供理論依據。
1 材料與方法
1.1 動物、材料與試劑
30 只SPF級C57BL/6J雄性小鼠,體質量(20±2)g,購自第四軍醫大學動物實驗中心(生產許可證號:SCXK(陜)2014-002)。
唐古特白刺干果購于青海西寧市城東區中藥批發市場。
XDA-6大孔樹脂 西安藍曉新材料有限公司;總膽固醇(total cholesterol,TC)、總甘油三酯(total triglyceride,TG)、高密度脂蛋白(high density lipoprotein,HDL)和低密度脂蛋白(low density lipoprotein,LDL)測定試劑盒 長春匯力生物技術有限公司;谷草轉氨酶(aspartate aminotransferase,AST)、谷丙轉氨酶(alanine aminotransferase,ALT)、丙二醛(malondialdehyde,MDA)、超氧化物歧化酶(superoxide dismutase,SOD)和谷胱甘肽過氧化物酶(glutathione peroxidase,GSH-Px)測定試劑盒、過氧化物酶增殖體激活受體-α(peroxisome proliferative activated receptor-α,PPAR-α)、肝X受體α(liver X receptor-α,LXR-α)、脂肪酸轉位酶(fatty acid translocase,CD36)和乙酰輔酶A羧化酶(acetyl coenzyme A carboxylase,ACC)酶聯免疫吸附測定(enzyme-linked immunosorbent assay,ELISA)試劑盒 南京建成生物工程研究所;無水甲醇、無水乙醇和多聚甲醛等試劑均為國產分析純;超純水為自制。
1.2 儀器與設備
DFT-50粉碎機 溫嶺市林大機械有限公司;TCX-600S超聲波清洗機 濟寧天宇超聲機械設備有限公司;1510型全波長酶標儀 美國Thermo Scientific公司;H1850型離心機 湖南湘儀實驗室儀器開發有限公司。
1.3 方法
1.3.1 唐古特白刺果實花色苷的制備
按照本實驗室建立的唐古特白刺總花色苷的分離純化方法[11]制備唐古特白刺總花色苷樣品。具體步驟如下:用粉碎機粉碎唐古特白刺干果,粉末用0.1%(體積分數,下同)HCl-70%甲醇溶液以料液比1∶10(m/V)、70 ℃提取32 min,提取液用旋轉蒸發儀除去甲醇,然后用XDA-6大孔樹脂分離純化,得到純化后的唐古特白刺果實花色苷。將唐古特白刺果實花色苷溶液冷凍干燥保存于-20 ℃冰箱待用。采用高效液相色譜面積歸一化法測得唐古特白刺果實花色苷中酰化花色苷占總花色苷的77.76%。
1.3.2 實驗動物分組與喂養
30 只小鼠適應性喂養1 周后,隨機分為3 組(每組10 只):對照組(N)、模型組(M)和唐古特白刺果實花色苷組(TA)。飼養條件:溫度(22±2)℃,相對濕度(55±5)%,12 h光照/12 h黑暗飼養,自由進食與飲水。普通嚙齒動物飼料為40%(質量分數,下同)玉米粉、26%小麥粉、10%麩皮、10%魚粉、10%豆餅、2%礦物質、1%粗粉和1%維生素,高脂飼料為普通飼料與牛油以質量比3∶1制得,添加唐古特白刺果實花色苷的高脂飼料為普通飼料+1%花色苷粉末與牛油(質量比3∶1)[12],小鼠花色苷粉末攝入量為479.62 mg/kg。所有實驗均遵守中國第四軍醫大學動物倫理委員會頒布的《實驗動物管理條例》。對照組喂食普通飼料;模型組喂食高脂飼料;TA組喂食添加唐古特白刺果實花色苷的高脂飼料,持續飼養14 周,監測小鼠每天的進食量及形態、行動特征,每周末稱量小鼠體質量一次,最后1 d禁食不禁水12 h,摘眼球取血,測量小鼠體長(鼻尖至肛門的長度),并收集肝組織、腹部脂肪和內臟脂肪。所有實驗均按照國家科學技術委員會發布的實驗動物管理指南(ISBN-100-309-15396-4)進行。
1.3.3 脂體指數的測定
小鼠處死后分離附睪周圍脂肪并稱質量,按式(1)計算脂體指數。

1.3.4 Lee’s指數的測定
小鼠處死前,測量每組小鼠的體質量和體長,按式(2)計算Lee’s指數。

1.3.5 血清AST和ALT活力、TC和TG質量濃度、HDL和LDL濃度的測定
收集的血液樣本于3 000×g離心15 min,取上層血清于1.5 mL離心管中4 ℃貯存備用。血清AST和ALT活力、TC和TG質量濃度、HDL和LDL濃度均依據相應的試劑盒說明書進行測定。
1.3.6 肝勻漿中MDA含量、SOD和GSH-Px活力、PPAR-α、LXR-α、CD36和ACC質量濃度的測定
稱取0.5 g肝臟組織,加入4.5 mL生理鹽水后用手持勻漿機3 000 r/min充分勻漿,肝勻漿于3 000×g離心10 min,取上清液于離心管中貯存備用。MDA含量、SOD和GSH-Px活力、PPAR-α、LXR-α、CD36和ACC質量濃度均按相應試劑盒說明進行測定。
1.4 數據統計與分析
實驗數據以 ±s表示,用GraphPad Prism 5.01軟件中單因素方法分析和Tukey’s Multiple Comparison Test進行分析并作圖,P<0.05為有顯著性差異。
2 結果與分析
2.1 唐古特白刺果實花色苷對高脂飲食小鼠體質量的影響
整個實驗過程中,對照組小鼠食欲正常、毛色光潔且行動靈活,未出現異常特征。與對照組比較,模型組小鼠毛色油亮、行動遲緩且體型肥胖。與模型組比較,TA組小鼠毛色和體型得到很好地改善,毛色光潔、體態均勻靈活。實驗過程中各組小鼠均未死亡。
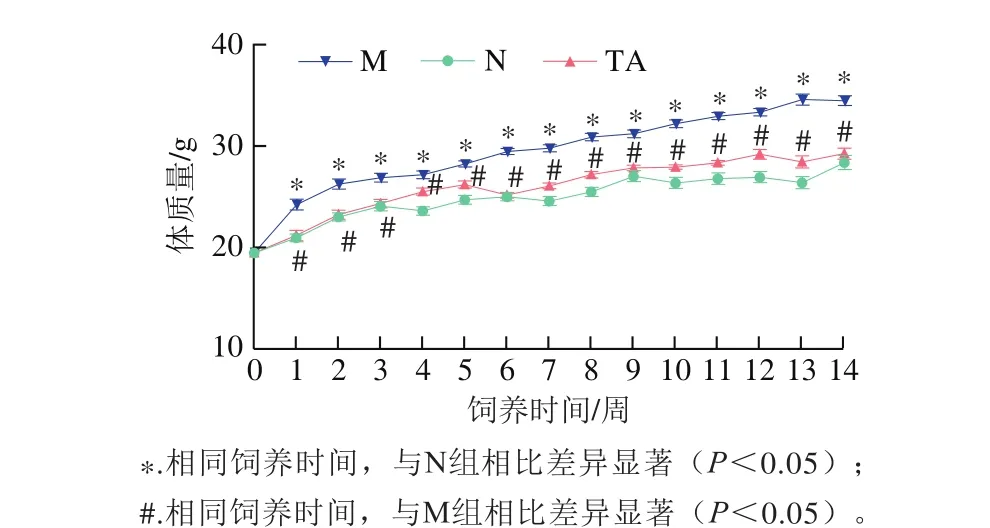
圖1 唐古特白刺果實花色苷對高脂飲食小鼠體質量的影響Fig. 1 Effects of anthocyanins from N. tangutorun Bobr. on HFD-induced body mass gain in C57BL/6J mice
體質量異常是長期高脂飲食引起的主要健康問題之一[13]。如圖1所示,從第2周開始,模型組小鼠體質量顯著高于對照組,TA組小鼠體質量顯著低于模型組,對照組和TA組小鼠體質量沒有顯著性差異;第9周后,模型組小鼠體質量增長速率并未減慢,對照組和TA組小鼠體質量增長緩慢。與模型組相比,TA組小鼠體質量增長情況得到改善。
2.2 唐古特白刺果實花色苷對高脂飲食小鼠常規生理指標的影響

表1 唐古特白刺果實花色苷對C57BL/6J小鼠常規生理指標的影響Table 1 Effects of anthocyanins from N. tangutorun Bobr. on food intake, abdominal fat mass, body fat index, visceral fat mass, body length and Lee’s index in HFD-fed C57BL/6J mice
表1結果顯示,對照組、模型組和TA組小鼠進食量無明顯差異。連續14 周攝入高脂飲食導致模型組小鼠的腹部脂肪質量、脂體指數、內臟脂肪質量以及Lee’s指數與對照組小鼠比較分別顯著增加了263.64%、192.16%、90.91%和8.93%(P<0.05)。而與模型組比較,TA組小鼠腹部脂肪質量、脂體指數、內臟脂肪質量以及Lee’s指數顯著降低,且Lee’s指數與對照組相比無顯著差異。
2.3 唐古特白刺果實花色苷對高脂飲食小鼠肝臟損傷的影響
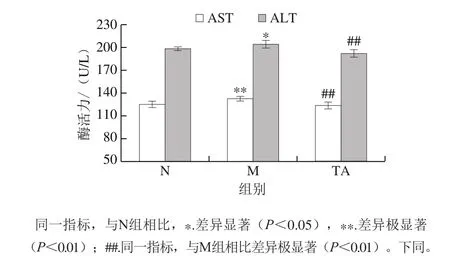
圖2 唐古特白刺果實花色苷對C57BL/6J小鼠血清中AST和ALT活力的影響Fig. 2 Effects of anthocyanins from N. tangutorun Bobr. on AST and ALT activity in serum of C57BL/6J mice
血清AST和ALT活力是評價肝損傷程度的經典指標,肝損傷發生時,血清AST和ALT活力會顯著升高[14]。如圖2所示,與對照組比較,模型組小鼠血清中AST和ALT活力顯著增加,表明長期攝入高脂飲食導致了明顯的肝損傷。而與模型組比較,唐古特白刺果實花色苷極顯著降低了小鼠血清中AST和ALT活力(P<0.01)。
2.4 唐古特白刺果實花色苷對高脂飲食小鼠肝臟脂肪代謝的影響
長期攝入高脂飲食在導致體質量異常增加的同時,通常伴隨著脂質代謝紊亂的發生,而脂質代謝紊亂是NAFLD的主要誘因[15]。肝臟發生脂肪性病變時,脂肪代謝能力下降,導致血液中TC和TG質量濃度上升。LDL和HDL與膽固醇在體內的運輸相關,其濃度會隨著血液中TC和TG質量濃度的變化而變化[16]。如圖3所示,與對照組比較,模型組小鼠血清中TC、TG質量濃度和LDL濃度極顯著增加(P<0.01),而HDL濃度極顯著降低(P<0.01),表明長期攝入高脂飲食導致了明顯的肝臟脂肪代謝異常。而與模型組比較,唐古特白刺果實花色苷極顯著降低了小鼠血清中TC、TG質量濃度和LDL濃度(P<0.01),并極顯著提高了血清HDL濃度(P<0.01)。

圖3 唐古特白刺果實花色苷對C57BL/6J小鼠血清中TC和TG質量濃度(A)、LDL和HDL濃度(B)的影響Fig. 3 Effects of anthocyanins from N. tangutorun Bobr. on TC and TG (A), LDL and HDL (B) levels in serum of C57BL/6J mice
2.5 唐古特白刺果實花色苷對高脂飲食小鼠肝勻漿中MDA含量、SOD和GSH-Px活力的影響


圖4 唐古特白刺果實花色苷對C57BL/6J高脂飲食小鼠肝勻漿中MDA含量(A)、SOD活力(B)和GSH-Px活力(C)的影響Fig. 4 Effects of anthocyanins from N. tangutorun Bobr. on MDA (A),SOD (B) and GSH-Px (C) levels in liver homogenate of C57BL/6J mice
如圖4所示,與對照組比較,模型組小鼠肝臟MDA含量和SOD活力極顯著升高,GSH-Px活力極顯著降低(P<0.01)。與模型組比較,TA組小鼠肝臟MDA含量和SOD活力極顯著降低,GSH-Px活力極顯著升高(P<0.01)。
2.6 唐古特白刺果實花色苷對高脂飲食小鼠肝勻漿中PPAR-α、LXR-α、CD36和ACC質量濃度的影響
如圖5所示,與對照組比較,模型組小鼠肝勻漿中LXR-α、CD36和ACC質量濃度極顯著升高(P<0.01),而PPAR-α質量濃度極顯著降低(P<0.01)。與模型組比較,TA組小鼠肝勻漿中的LXR-α、CD36和ACC質量濃度極顯著降低,PPAR-α質量濃度極顯著升高(P<0.01)。

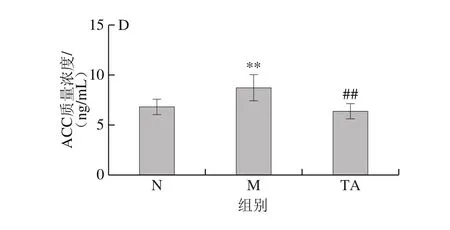
圖5 唐古特白刺果實花色苷對C57BL/6J小鼠肝勻漿中PPAR-α(A)、LXR-α(B)、CD36(C)和ACC(D)質量濃度的影響Fig. 5 Effects of anthocyanins from N. tangutorun Bobr. on PPAR-α (A),LXR-α (B), CD36 (C) and ACC (D) levels in liver homogenate of C57BL/6J mice
3 討 論
高脂飲食誘導的肥胖在動物模型中主要表現為體質量增加、體脂積累、肝臟損傷和血脂異常[17]。小鼠連續14 周攝入高脂飲食引起了其體質量、腹部脂肪質量、Lee’s指數、血清AST和ALT活力、TC和TG質量濃度顯著增加,與已有報道的結果[17-18]一致,進一步證實了長期攝入高脂飲食會導致肥胖、肝臟損傷和高甘油三酯血癥。高脂飲食誘導的血脂異常不僅與增加血清TC、TG質量濃度和LDL濃度有關,也與降低血清中的HDL濃度緊密相關[19]。與模型組相比,TA組小鼠體質量增長情況明顯改善,脂體指數、內臟脂肪質量以及Lee’s指數顯著降低,表示脂肪肝得到一定改善;血清中AST和ALT活力、TC和TG質量濃度、LDL濃度極顯著降低,HDL濃度極顯著升高,表明唐古特白刺果實花色苷能夠改善肝損傷和肝臟脂肪代謝。
體質量異常與血脂代謝紊亂僅是高脂飲食誘導NAFLD的前期階段,在脂質過氧化和氧化應激的作用下NAFLD將進一步惡化[20]。活性氧具有與脂質反應的巨大潛力,特別是與多種不飽和脂肪酸發生反應,從而導致細胞膜損傷。MDA被廣泛用作脂質過氧化和氧化應激的標志物,其在機體內的水平會因為高脂飲食的攝入而顯著提高[21]。SOD是自由基天然清除劑,當機體受到感染、創傷并出現一個或多個臟器功能衰竭時,體內SOD活力增高[22]。本研究發現,模型組小鼠肝臟MDA含量和SOD活力顯著高于對照組,這表明高脂飲食引起了小鼠體內脂質過氧化程度的加深,且有肝臟功能衰竭跡象,與先前報道結果[22]一致。體內固有的抗氧化酶GSH-Px能通過清除活性氧和脂質過氧化產物而有效預防機體的氧化應激損傷[23],但高脂飲食的攝入降低了體內GSH-Px的活力,進而削弱機體的天然抗氧化防御系統。與模型組相比,TA組小鼠肝組織中MDA含量和SOD活力極顯著降低,GSH-Px活力極顯著升高,說明唐古特白刺果實花色苷通過調節肝臟氧化應激改善高脂飲食誘導的NAFLD損傷。
肝臟是脂肪合成和分解的重要場所[24]。脂肪酸合成起始于乙酰-CoA轉化成丙二酸單酰-CoA,這步反應是在ACC作用下實現的。在脂肪合成途徑中,ACC催化的反應不僅是第1步反應,同時也是限速步驟[25]。LXR-α激活后能增加脂肪酸合成相關基因的表達,提高血漿甘油三酯的水平[26]。LXR還能直接調節脂肪酸合成相關基因如脂肪酸合成酶、ACC和硬脂酰輔酶A去飽和酶1的表達[27]。Koonen等發現CD36主要通過促進TG的合成增加脂肪酸的攝取,從而加速NAFLD的形成[28]。本研究發現,模型組小鼠肝臟ACC、LXR-α和CD36質量濃度顯著高于對照組,與先前報道結果[28]一致。PPAR-α是脂質代謝過程的關鍵因子[29],PPAR-α水平的降低會引起一系列與脂質代謝相關蛋白質和酶基因轉錄水平的降低,使肝臟線粒體內脂肪酸的β氧化能力減弱,引發脂質堆積。脂肪沉積的肝細胞可發生脂性改變,誘發和加劇胰島素抵抗及炎癥反應,促進NAFLD的發生[30]。本研究結果表明,與模型組相比,TA組小鼠肝組織中ACC、LXR-α和CD36質量濃度極顯著降低,PPAR-α質量濃度極顯著提高,說明唐古特白刺果實花色苷通過調節肝臟脂質代謝改善高脂飲食誘導的小鼠NAFLD損傷。
綜上所述,唐古特白刺果實花色苷能明顯降低高脂飲食小鼠的體質量、腹部脂肪質量、脂體指數、內臟脂肪質量、體長以及Lee’s指數,降低高脂小鼠血清AST和ALT活力、TC和TG質量濃度、LDL濃度,提高HDL濃度,通過降低肝組織勻漿中MDA含量、SOD活力并提高GSH-Px活力緩解高脂飲食導致的肝臟氧化應激損傷,并通過降低脂肪合成相關酶和受體(ACC、LXR-α和CD36)質量濃度、上調脂肪分解相關的轉錄因子(PPAR-α)質量濃度調節肝臟脂質代謝,最終改善高脂飲食誘導的NAFLD損傷。因此,唐古特白刺果實花色苷作為潛在的功能食品在NAFLD的治療中具有重要的意義。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
學苑創造·A版(2020年9期)2020-10-13 09:41:02
中國生殖健康(2019年2期)2019-08-23 08:12:08
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
產品可靠性報告(2017年7期)2017-09-05 09:49:12
汽車觀察(2016年3期)2016-02-28 13:16:26
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
中國質量與標準導報(2014年1期)2014-02-28 22:21:28
