白三烯類成分在動脈粥樣硬化中的研究進(jìn)展
2019-11-22 04:54:58黃家欣都文文黃榮增宋成武金姝娜
中國醫(yī)藥導(dǎo)報(bào) 2019年29期
關(guān)鍵詞:途徑
黃家欣 田 稷 倪 維 都文文 黃榮增 宋成武 謝 云 金姝娜
1.湖北中醫(yī)藥大學(xué)藥學(xué)院,湖北武漢 430065;2.湖北省中醫(yī)院檢驗(yàn)科,湖北武漢 430060;3.河南省焦作市人民醫(yī)院藥劑科,河南焦作 454000;4.華中科技大學(xué)公共衛(wèi)生學(xué)院,湖北武漢 430030
Feldberg 和Kellaway[1]于1938~1940 年首次發(fā)現(xiàn)白三烯C 化合物(leukotriene C,LTC),稱其為“緩慢的平滑肌刺激物質(zhì)”。隨后由瑞典學(xué)家Bengt Samuelsson[2]于1979 年命名為“白三烯”,其名稱來源于“白血球”和“三烯”。以往的綜述僅基于LTs 某一類化合物或相關(guān)聯(lián)的某一種疾病進(jìn)行講解,現(xiàn)通過PubMed 和Web of Science 等網(wǎng)上資料庫評估現(xiàn)有動物實(shí)驗(yàn)和人體臨床研究文獻(xiàn)中LTs 和該類疾病之間的相關(guān)性,總結(jié)在相應(yīng)部位測量白三烯含量可用于相關(guān)疾病的病情監(jiān)測及治療方案的調(diào)整,同時確定白三烯拮抗劑和白三烯受體拮抗劑在相關(guān)疾病治療中的關(guān)系,為基于LTs 疾病生物標(biāo)志物的篩選及其應(yīng)用于臨床靶向新型治療方案的研究提供參考,提高未來藥物治療的優(yōu)質(zhì)選擇。
1 LTs 存在形式及代謝途徑
1.1 存在形式
白三烯類成分(LTs)以一條具有四對雙鍵的二十碳長鏈為結(jié)構(gòu)特征,其中三對共軛。該類成分由炎癥細(xì)胞分泌,如中性粒細(xì)胞、巨噬細(xì)胞等,在體內(nèi)主要有A4、B4、C4、D4、E4、F4 六種主要存在形式。LTs 半衰期短,在體內(nèi)代謝快,主要通過膽汁、尿液排出。其中LTA4 是白三烯體系中最不穩(wěn)定的成分,常被LTA4 水解酶(leukotriene A4 hydrolase,LTA4H)、LTC4 合成酶(leukotriene C4 synthetase,LTC4S)迅速氧化成LTB4、LTC4,研究較少。LTB4 具有較強(qiáng)的白細(xì)胞趨化作用及免疫調(diào)節(jié)作用,屬于白三烯二羥酸類。LTC4、LTD4、LTE4、LTF4 屬于半胱氨酰白三烯類(Cys-LTs)[3],其特征是化學(xué)結(jié)構(gòu)C6 位上均有一個硫醚連接的肽,需要通過與激活的半胱氨酰白三烯受體(cysteinyl leukotrienes receptor,Cys-LTR)結(jié)合產(chǎn)生活性,能誘導(dǎo)正常外周血液中白細(xì)胞;LTC4 與LTE4 結(jié)構(gòu)相似,由β1 和β2 整合素與內(nèi)皮細(xì)胞表面結(jié)合,刺激嗜酸性粒細(xì)胞分泌而來;LTD4 由外周血嗜酸性粒細(xì)胞經(jīng)白三烯D4 合成酶(leukotriene D4 synthetase,LTD4S)或γ-谷酰基轉(zhuǎn)移酶(γ-glutamyl transpeptidase,γ-GT)代謝生產(chǎn),和LTF4 一樣都是LTC4 的代謝產(chǎn)物,其中LTD4 脫掉的為谷胱甘肽的谷氨酸殘基,而LTF4 結(jié)構(gòu)中缺失的是谷胱甘肽的甘氨酸殘基[4]。6 種白三烯化學(xué)結(jié)構(gòu)見圖1。
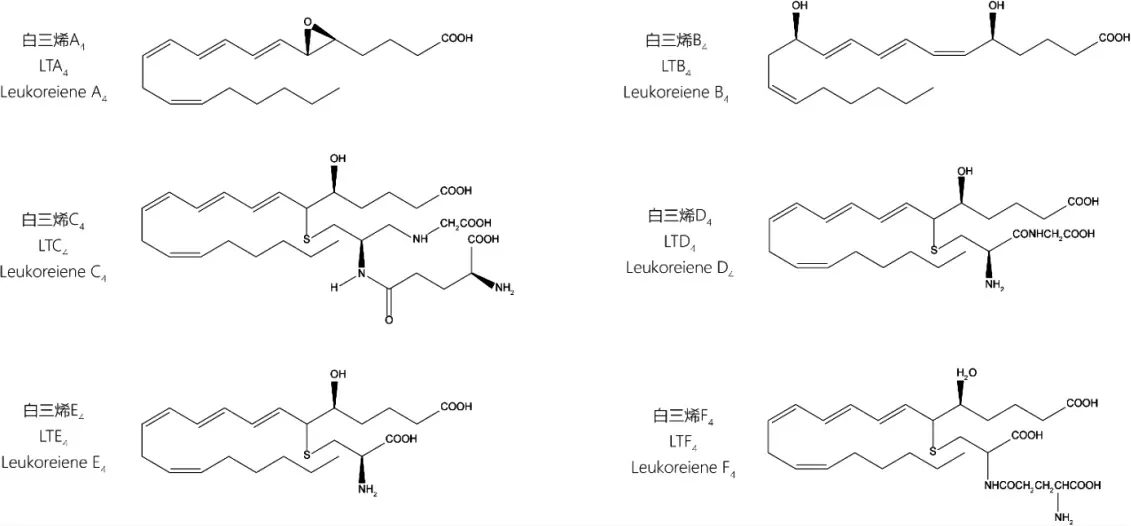
圖1 白三烯類成分LTA4、LTB4、LTC4、LTD4、LTE4、LTF4 化學(xué)結(jié)構(gòu)式
1.2 代謝途徑
LTs 生物合成啟動需要抗原與肥大細(xì)胞表面IgE受體相互作用,免疫反應(yīng)過程中,接觸變應(yīng)原后產(chǎn)生的炎性反應(yīng)能刺激并激活嗜曙紅細(xì)胞等炎癥細(xì)胞細(xì)胞膜、核膜磷脂雙分子層上的膜磷脂或甘油二酯經(jīng)磷脂酶A2(phospholipase A2,PLA2)釋放AA[5-7]。AA 存在兩條代謝途徑:①環(huán)氧化酶(cyclooxygenase,COX)途徑,生成前列腺素類[8];②脂氧合酶途徑,5-LO 代謝生成5-氫化氧化二十碳四烯酸和LTA4[9]。之后,部分LTA4 被LTA4H(有環(huán)氧化物水解酶、氨肽酶雙重活性[10],是水解LTA4 產(chǎn)生LTB4 的關(guān)鍵酶[11])轉(zhuǎn)化為LTB4 和水,再通過LTC4S 和谷胱甘肽作用生成LTC4,再經(jīng)γ-GT 脫去谷氨酸生成LTD4,LTD4 在二肽酶參與轉(zhuǎn)換下生成LTE4[12-13]。其代謝途徑見圖2。
2 LTs 受體
LTs 受體分為兩大亞型:LTB4 受體(BLT)和Cys-LTR,可再細(xì)分成BLT1、BLT2、Cys-LT1R 和Cys-LT2R四個小亞型[14]。其中LTB4 的受體BLT1 與其親和力較高,主要介導(dǎo)化學(xué)誘導(dǎo)和炎性反應(yīng),常在白細(xì)胞中表達(dá);而BLT2 與LTB4 親和力低,幾乎遍布于全身各個組織,作用尚不明確[15]。Cys-LT1R 主要分布于血管壁,可識別LTC4、LTD4、LTE4(與受體結(jié)合能力的順序?yàn)椋篖TD4>LTC4>LTE4),因其廣泛分布于巨噬細(xì)胞、嗜酸性粒細(xì)胞、肥大細(xì)胞等[16-17],成為治療LTs 疾病藥物的首要作用靶點(diǎn);Cys-LT2R 也可識別LTC4、LTD4、LTE4(與受體結(jié)合能力的順序?yàn)椋篖TD4=LTC4>LTE4),在炎癥、血管通透性、組織纖維化中起重要作用。
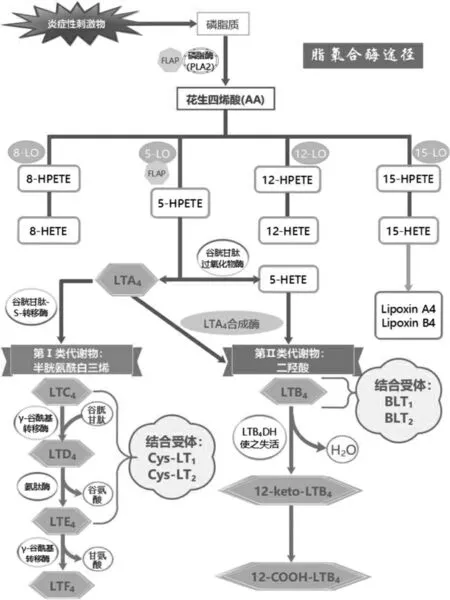
圖2 白三烯脂氧合酶途徑代謝過程及其產(chǎn)物
3 LTs 與AS 的相關(guān)性
3.1 AS 簡介
高脂血癥(HL)是脂代謝異常最為相關(guān)的疾病,作為AS 早期癥狀,臨床表現(xiàn)為血清總膽固醇、三酰甘油、低密度脂蛋白的異常升高。降血脂藥物他汀類通常用于治療HL,使用后可使大多數(shù)人血清總膽固醇降低約50%[18],但對于有HL 病癥而無其他健康問題的患者來說是否有效,存在著爭議。因此,與脂質(zhì)代謝密切相關(guān)的AA 代謝途徑及其相關(guān)代謝產(chǎn)物成為新的研究重點(diǎn)之一。研究表明,高膽固醇血癥可引起中性粒細(xì)胞數(shù)量增多,隨即產(chǎn)生脂質(zhì)介質(zhì),如強(qiáng)化學(xué)趨化劑LTB4,通過自分泌方式增加中性粒細(xì)胞的動能和數(shù)量,其合成取決于其產(chǎn)生途徑的游離花生四烯酸含量和可用性及5-LO 亞細(xì)胞定位:中性粒細(xì)胞休眠時,5-LO 位于細(xì)胞質(zhì)中,一旦被激活,5-LO 會重新分配到核膜上,啟動LTB4 的生物合成,由其C-末端催化區(qū)域(p-5-LO)通過細(xì)胞膜外信號調(diào)節(jié)激酶(extracellular signal regulated kinases,ERKs)介導(dǎo)進(jìn)行[19]。HL 早期ERKs 增加中性粒細(xì)胞中LTB4 的生成,中后期將引起器官功能性褪變,加重血管負(fù)擔(dān),從而誘發(fā)AS[20]。
3.2 LTs 參與AS 的途徑
由近幾年文獻(xiàn),此文總結(jié)出四大途徑,具體途徑及其參與物質(zhì)見圖3。

圖3 LTs 參與AS 的四條途徑
3.2.1 FLAP、LTA4H、LTC4S LTs 通過提高白細(xì)胞趨藥性,加速心血管炎癥及鈣化,其合成路線上的酶類與AS 的發(fā)病機(jī)制密不可分[21]。FLAP 激活蛋白位于人類ALOX5AP 基因染色體上,其中遺傳變異體rs4769874能增加基因產(chǎn)物FLAP 表達(dá),從而增加冠狀動脈疾病(coronary artery disease,CAD)風(fēng)險(xiǎn),放大LTs 途徑的炎性反應(yīng)和導(dǎo)致AS 的產(chǎn)生;rs9579646 則減少ALOX5AP的表達(dá)和其基因產(chǎn)物FLAP,降低CAD 風(fēng)險(xiǎn)[22]。動物體內(nèi)實(shí)驗(yàn)表明[23],ALOX5 基因敲除的小鼠能增加環(huán)氧合酶途徑COX-2 和PPAR-γ 的表達(dá),其激活的PON1基因能減少LDL 的氧化,還能水解ox-LDL,減少單核細(xì)胞的激活,阻止他們黏附在內(nèi)皮細(xì)胞上,從而阻止AS 的進(jìn)程。LTA4H 是LTB4 合成過程中的關(guān)鍵蛋白,涉及AA 的新陳代謝。LTC4S 是LTC4 合成酶,與谷胱甘肽共同組成LTC4 這種血管收縮劑。有實(shí)驗(yàn)表明[23],與健康對照組比較,在AS 斑塊中這三種酶的mRNA 水平有顯著增加趨勢,且FLAP、LTA4H 蛋白在AS 病灶的巨噬細(xì)胞中大量聚集。此外,F(xiàn)LAP、LTA4H、LTC4S 代謝通路上產(chǎn)生重要的炎癥介質(zhì),其豐富的表達(dá)可提高持續(xù)性AS 患者中斑塊的不穩(wěn)定。實(shí)驗(yàn)證明在給阿曲留通(VIA-2291,5-LO 活性抑制劑)24 周后,通過冠狀計(jì)算機(jī)斷層掃描,抑制劑組的34 例患者AS 斑塊量相對于安慰劑組大幅度下降[24],由此可知這條途徑中關(guān)鍵酶類的拮抗劑可成為藥物干預(yù)的潛在靶標(biāo)[25]。
3.2.2 MMPs、HNE 金屬蛋白酶類(MMPs)是一組能特異降解細(xì)胞外基質(zhì)成分的蛋白水解酶家族,位于泡沫細(xì)胞的胞質(zhì)中,增加MMPs 活性是介導(dǎo)穩(wěn)定型AS 斑塊向不穩(wěn)定表現(xiàn)型發(fā)展的必要物質(zhì)條件。有研究提出,在有癥狀的AS 斑塊中,刺激巨噬細(xì)胞而激活的5-LO 能通過LTB4-BLT1 途徑、CysLTs 途徑分別促進(jìn)血管平滑肌細(xì)胞(VSMC)、巨噬細(xì)胞中MMP-2、MMP-9的釋放[26],后者通過軟化細(xì)胞外基質(zhì),降解斑塊內(nèi)保護(hù)性纖維帽,從而促進(jìn)斑塊破裂[27]。在急性缺血綜合征的預(yù)防及穩(wěn)定保護(hù)糖尿病患者AS 斑塊的防治策略中,LTs 受體拮抗劑通過抑制LTs 受體,使LTs 及其衍生物產(chǎn)生減少,從而降低MMPs 的表達(dá)[28],預(yù)防和治療糖尿病患者急性AS 血栓形成,為該類新藥的研發(fā)提供了方向。
3.2.3 LTB4 研究發(fā)現(xiàn)肥胖組織中LTB4 明顯增加,且其高親和受體——BLT1,作為新的化學(xué)誘導(dǎo)物受體角色,通過LTB4/BLT1 途徑促進(jìn)脂肪組織中的單核細(xì)胞聚集,導(dǎo)致慢性炎癥。LTB4 通過上調(diào)細(xì)胞間黏附因子(ICAM-1)和血管細(xì)胞黏附分子(VCAM-1)分別與其在單核細(xì)胞/巨噬細(xì)胞表面的受體整合素β1、β2 的親和力,促進(jìn)單核細(xì)胞黏附和貼壁,啟動內(nèi)皮反應(yīng)。選擇性提高5-LO 基因表達(dá)的氧化型心磷脂,激活5-LO 生成其衍生物L(fēng)TB4,增加單核細(xì)胞/巨噬細(xì)胞內(nèi)游離Ca2+濃度,增加腫瘤壞死因子(TNF-α)誘導(dǎo)血管細(xì)胞間黏附因子表達(dá)[29],還能增加ICAM-1、VCAM-1水平,加速動脈粥樣硬化。LTB4 還能從I-κB 激酶β/NF-κB 通路促進(jìn)血管結(jié)合激活的BLT1 促使單核細(xì)胞中CCL2 的產(chǎn)生[30],促進(jìn)其在血管內(nèi)皮的聚集和黏附,增加血管通透性,使VSMC 增殖、遷移,增加單核趨化蛋白1(MCP-1)產(chǎn)生[31],參與動脈粥樣硬化早期進(jìn)程。粥樣硬化斑塊組織中,巨噬細(xì)胞產(chǎn)生脂肪酶、活性氧、自由基等使LDL 分子氧化,并通過清道夫受體攝取ox-LDL。LTB4 能使單核細(xì)胞/巨噬細(xì)胞膜表面ox-LDL 受體表達(dá)上調(diào),激活內(nèi)皮細(xì)胞、單核細(xì)胞、巨噬細(xì)胞、T 細(xì)胞,引起巨噬細(xì)胞吞噬膽固醇脂質(zhì),最終轉(zhuǎn)化成泡沫細(xì)胞,導(dǎo)致斑塊內(nèi)脂質(zhì)堆積,形成不穩(wěn)定斑塊。且溶血性卵磷脂(LPC)在ox-LDL 誘發(fā)的免疫活化中起重要作用,能促進(jìn)LDL 向ox-LDL 轉(zhuǎn)化。因此抑制巨噬細(xì)胞攝取ox-LDL,從而減少血管炎癥及脂質(zhì)堆積,也能成為一個新的治療AS 的方向。
3.2.4 LTC4、LTD4 LTC4、LTD4 能誘導(dǎo)氣管平滑肌收縮,使支氣管哮喘患者的白細(xì)胞產(chǎn)生強(qiáng)烈趨化性[32],在氣道黏液分泌、膠原合成中起著重要作用[33]。其小部分通過結(jié)合CysLT2R 誘導(dǎo)大量Ca2+進(jìn)入人臍靜脈內(nèi)皮細(xì)胞,增加TNF-α 誘導(dǎo)血管細(xì)胞間黏附因子表達(dá);還能從兩個受體的不同途徑——CysLT2R/Rho 激酶(Rho GTP 酶超家族的Rho 亞家族)途徑和CysLT1R/Erk(細(xì)胞外調(diào)節(jié)蛋白激酶)依賴途徑,誘導(dǎo)內(nèi)皮細(xì)胞收縮,加速動脈粥樣硬化,顯著誘導(dǎo)AS 患者冠狀動脈收縮。大部分與CysLT1R 結(jié)合,通過MAPK 和核轉(zhuǎn)錄因子-κB(NF-κB)途徑誘發(fā)人單核細(xì)胞/巨噬細(xì)胞中MCP-1 表達(dá)[34]。MAPK 屬于絲氨酸蘇氨酸激酶群體,在各種不同細(xì)胞功能的信號傳導(dǎo),哺乳動物細(xì)胞中有3 種主要MAPK 亞型:ERK1/2、JNK、p38 MAPK。在抗原呈遞細(xì)胞中,ERK 信號通路促進(jìn)最初的天然T 輔助細(xì)胞向Th2 表現(xiàn)型分化,從而激活Ras/ERK 通路,介導(dǎo)自身炎癥免疫反應(yīng);JNK 激活轉(zhuǎn)錄因子激活蛋白,能編碼各種不同種類的免疫調(diào)節(jié)基因。LTC4、LTD4 能顯著加速ERK1/2、JNK 途徑磷酸化過程,控制一些炎癥細(xì)胞激活過程,進(jìn)而導(dǎo)致斑塊趨于不穩(wěn)定狀態(tài)。NF-κB 是基因中普遍存在的轉(zhuǎn)錄因子,編碼前炎癥細(xì)胞活素、趨化因子、黏附因子等,LTD4 在人上皮細(xì)胞中刺激NF-κB 途徑活化;而THP-1 細(xì)胞和外周血CD14+中[34],LTC4、LTD4 通過結(jié)合其受體CysLT1R誘導(dǎo)NF-κB 途徑活化,且THP-1 細(xì)胞中由LTC4、LTD4 誘導(dǎo)產(chǎn)生的MCP-1 能被NF-κB 途徑抑制劑所阻斷[34],可證明NF-κB 途徑能介導(dǎo)人單核細(xì)胞/巨噬細(xì)胞中CysLTs 誘導(dǎo)MCP-1 產(chǎn)生,從而參加到AS 進(jìn)程中[33]。多藥耐藥相關(guān)蛋白-1(MRP1),作為谷胱甘肽、LTC4、雌激素等的轉(zhuǎn)運(yùn)體,能介導(dǎo)LTC4 轉(zhuǎn)運(yùn)、血管功能和粥樣斑塊的形成。大量MRP1 存在于VSMCs中,對血管細(xì)胞中氧化應(yīng)激的管理具有重要作用,特別是在AS 早期,能通過血管緊張素Ⅱ刺激VSMCs 增加LTC4 生產(chǎn)和釋放,LTC4 的增加又導(dǎo)致活性氧含量上升,進(jìn)一步影響AS 發(fā)展。實(shí)驗(yàn)結(jié)果表明,這些釋放的活性氧也能被Cys-LT1R 拮抗劑孟魯司特所抑制,作為AS 治療作用的潛在方向[35]。由此可見,MRP1和Cys-LT1R 阻斷劑可能成為治療內(nèi)皮功能障礙和AS 的潛在作用靶點(diǎn)[36]。
4 小結(jié)
LTs 是一種強(qiáng)有力的細(xì)胞趨化因子,具有活化炎癥細(xì)胞的功能,且有較強(qiáng)的趨化作用及自身免疫調(diào)節(jié)作用,通過與其受體結(jié)合激活并聚集較多的炎癥細(xì)胞及免疫效應(yīng)細(xì)胞,參與全身性炎癥、過敏反應(yīng)。經(jīng)多種檢測手段,測量白三烯含量,可用于相關(guān)疾病的病情監(jiān)測及治療方案的調(diào)整。本文主要介紹了AS 及由各種途徑導(dǎo)致其斑塊趨于不穩(wěn)定甚至破裂誘發(fā)的疾病,已成為全球發(fā)病率和死亡率的新高,所以此文從白三烯入手,僅為新型藥物的開發(fā)及使用提供理論依據(jù),為相應(yīng)疾病的防治和治療開辟新道路新靶點(diǎn)。
猜你喜歡
語數(shù)外學(xué)習(xí)·高中版中旬(2023年2期)2023-05-10 13:26:53
語數(shù)外學(xué)習(xí)·高中版中旬(2022年5期)2022-07-13 20:47:51
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年10期)2019-11-25 07:33:58
中學(xué)生數(shù)理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中學(xué)生百科·大語文(2017年10期)2017-11-04 06:56:38
中國衛(wèi)生(2016年3期)2016-11-12 13:23:26
公民與法治(2016年22期)2016-05-17 04:20:13
中國衛(wèi)生(2014年12期)2014-11-12 13:12:52
癌變·畸變·突變(2014年6期)2014-02-27 06:15:03

- 中國醫(yī)藥導(dǎo)報(bào)的其它文章
- 白細(xì)胞介素-17A 水平及基因多態(tài)性與宮頸癌相關(guān)性
- 生育指數(shù)評分對子宮內(nèi)膜異位癥腹腔鏡術(shù)后妊娠方式的預(yù)測能效
- CD44V4 和nm23 在三陰性乳腺癌中的表達(dá)及其相關(guān)性
- 多學(xué)科護(hù)理照顧模式對腦膠質(zhì)瘤患者術(shù)后心理波動、家庭功能及生活質(zhì)量的影響
- 老年體外循環(huán)心臟手術(shù)患者術(shù)后認(rèn)知功能障礙的相關(guān)因素分析
- 基于患者體驗(yàn)為中心的醫(yī)療質(zhì)量關(guān)鍵指標(biāo)的構(gòu)建