飼料中發酵芝麻粕替代菜粕對草魚生長性能、腸道形態和微生物及小肽轉運相關基因表達的影響
2019-11-26 02:17:04曹申平唐建洲羅文婕鼎鄧岳松
水生生物學報 2019年6期
關鍵詞:實驗
宋 鵬 曹申平 唐建洲 彭 亮 羅文婕 熊 鼎鄧岳松 賀 喜 劉 臻
(1. 湖南農業大學動物科技學院, 長沙 410128; 2. 長沙學院生物與環境工程學院, 水生動物營養與品質調控湖南省重點實驗室,長沙 410022; 3. 唐人神集團股份有限公司, 株洲 412000)
芝麻粕是指芝麻經壓榨取油后的殘粕, 再經粉碎所得的副產品。芝麻粕中主要成分為蛋白質, 粗蛋白質含量45%以上, 其所具有的幾種主要必需氨基酸, 如蛋氨酸、胱氨酸、精氨酸以及色氨酸, 含量均高于菜粕和豆粕[1]。然而, 芝麻粕中的賴氨酸含量較低, 而且還含有草酸、植酸及少量的單寧, 這些抗營養因子會阻礙或者破壞營養物質的消化利用, 并對動物健康和生長性能產生不良影響, 從而制約了芝麻粕在畜禽和水產中的應用[1—3]。Yamauchi等[4]研究發現, 飼喂蛋雞(Gallus gallus domesticus)芝麻粕后, 能顯著抑制其生長; Mamputu等[5]同樣發現蛋雞在飼喂芝麻粕飼料后, 其飼料轉化率和增重率均低于對照組。微生物發酵是指在適宜的條件下, 利用微生物的代謝作用, 將原料中部分多糖、脂肪和蛋白質等大分子降解為有機酸、可溶性多肽等小分子的過程。微生物發酵法可以有效地減少芝麻粕中植酸等抗營養因子的含量, 同時對芝麻粕中營養物質破壞較小, 還能夠提高其粗蛋白和小肽等有益成分的含量[2,5]。因此, 微生物發酵消除芝麻粕中抗營養因子對于其在畜禽和水產飼料中的應用前景尤為重要。
菜粕是油菜籽經壓榨或浸提取油后的副產品,具有蛋白質豐富、低廉易得等優點。如今, 菜粕已成為我國水產和畜禽養殖飼料中一種重要的蛋白源, 此外, 菜粕中氨基酸組成不平衡, 且具有硫代葡萄糖甙、芥酸和植酸等抗營養因子, 嚴重制約了水產動物對其利用效率[6]。隨著我國養殖行業的快速發展, 對于菜粕等蛋白資源的需求也與日俱增, 菜粕資源也面臨短缺, 從而菜粕價格不斷上漲[1,6]。因此, 尋求菜粕的優質替代蛋白源也成為亟待解決的科學問題。
近年來, 我國水產品的產量呈逐年遞增趨勢,其中草魚(Ctenopharyngodon idellus)的產量最高,2016年, 我國草魚產量達5.8988×109kg[7]。草魚, 隸屬鯉形目鯉科草魚屬, 具有蛋白需求低、生長速度快、肉質鮮美等特點[8]。本實驗利用芽孢桿菌、酵母菌和乳酸桿菌協同發酵芝麻粕, 研究其替代菜粕蛋白添加到飼料中對草魚生長性能、腸道形態和微生物及尾型同源盒基因2 (Caudal-related homeobox 2,CDX2)、特異性蛋白1 (Specificity protein 1,Sp1)和小肽轉運載體(Oligopeptide transporter 1,PepT1)三個小肽轉運相關基因的影響, 為發酵芝麻粕在魚類飼料中的應用提供理論支撐。
1 材料與方法
1.1 實驗飼料
本實驗對照組飼料配方參照唐人神集團股份有限公司駱駝牌913飼料配方進行設計, 實驗用芝麻粕購買于河南正康糧油有限公司, 發酵芝麻粕(粗蛋白質45.52%、水分9.36%、鈣2.25%、總磷1.12%)的制備方法參照唐建洲CN107183356A發明專利[9]。實驗以5%、10%和15%發酵芝麻粕蛋白分別替代對照組飼料中11.8%、23.5%和35.1%的菜粕蛋白, 共設計了四組等氮(28.5%)等脂(5%)的飼料。實驗以豆油為主要脂肪源。具體飼料配方如表 1所示。各實驗原料經粉碎機粉碎后, 過40目篩, 以小型平模制粒機(陳氏機械廠, 浙江新昌, KL-105A型,2.5 mm)制成飼料顆粒, 自然風干后存于-4℃冰箱備用。
1.2 養殖系統和養殖條件
實驗在唐人神集團股份有限公司古大橋實驗基地(湖南, 株洲)室內循環水養殖系統中進行。循環水以6 L/min的速度進入每缸(體積400 L/缸)。在非投喂期間使用充氣頭連續充氣增氧。采用節能燈作為光源, 每天光照12h (8:00—20:00)。每天記錄水溫和水體pH, 每周監測循環水中氨氮、亞硝酸鹽和溶解氧含量。實驗期間水溫為18—26℃, pH為6.5—7.5, 氨氮<0.1 mg/L, 亞硝酸鹽<0.01 mg/L, 溶解氧>6.5 mg/L。
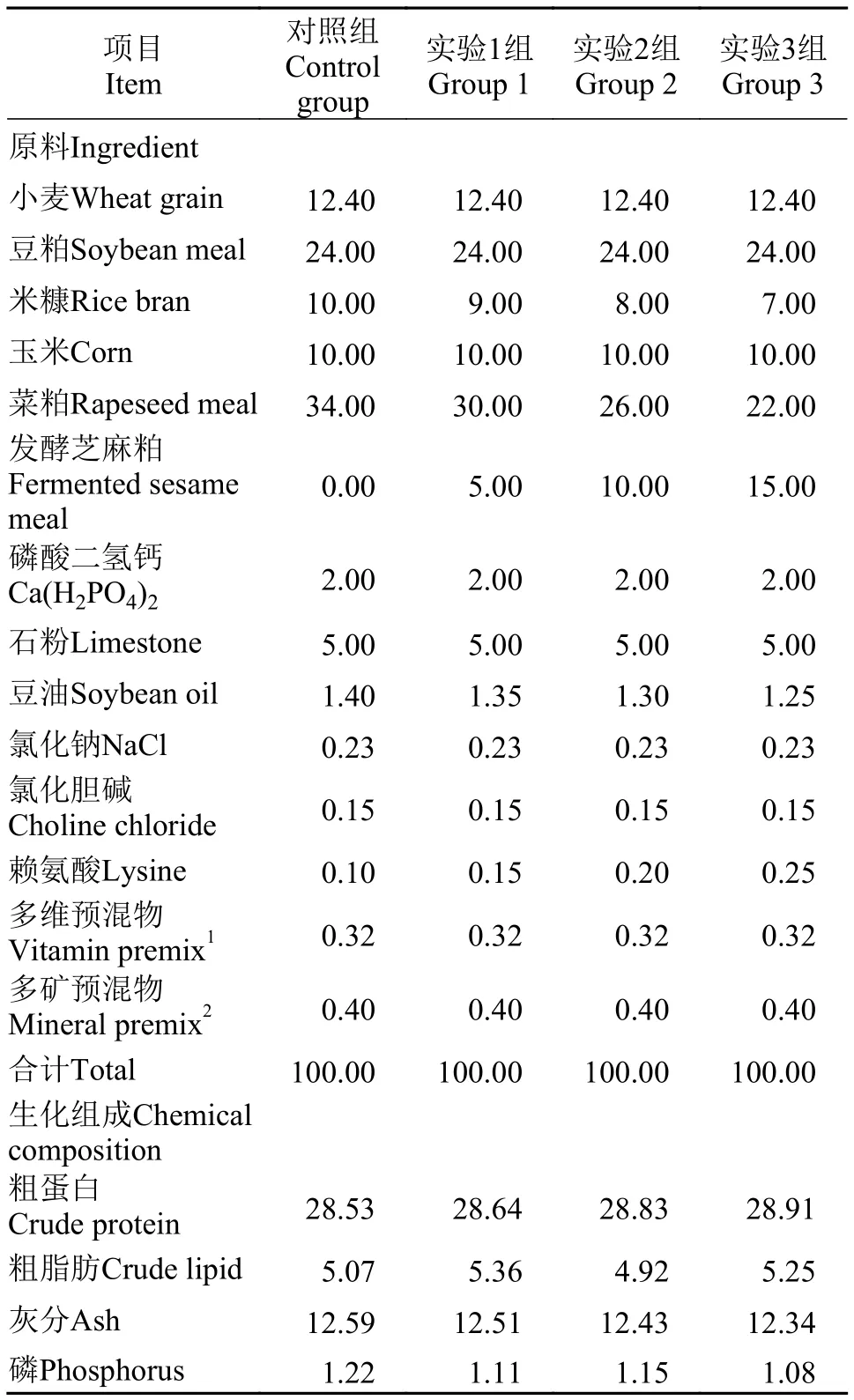
表1 飼料配方及生化組成(%干物質)Tab. 1 Formulation and Chemical composition of experimental diets (% dry matter)
1.3 實驗魚及飼養
實驗用草魚由湖南省水產科學研究所(湖南,長沙)提供, 在正式養殖實驗開始之前, 用對照組飼料馴養草魚2周。馴養期間, 每天飽食投喂2次(9:00和15:00)。養殖實驗開始前1周, 用4種實驗飼料混合后飽食投喂草魚, 使其適應實驗飼料。實驗前1d不進行投喂, 選取規格均勻、體格健壯的草魚[初重(99.98±0.69) g], 隨機放入12個養殖缸中, 每缸20尾。實驗共4個處理, 每處理3重復。生長實驗持續45d。實驗期間, 每天定時飽食投喂2次(9:00和15:00)。
1.4 樣品采集與指標測定
生長指標測定在實驗結束時, 草魚停止攝食24h后, 分別稱取每缸草魚總體重并記錄尾數, 結合攝食情況和飼料的基本組成統計增重率(Weight gain rate,WGR)、特定生長率(Special growth rate,SGR)、成活率(Survival rate,SR)、餌料系數(Feed conversion ratio,FCR)和蛋白質效率(Protein efficiency ratio,PER)。各組實驗魚的生長指標根據以下公式計算求得:

飼料常規營養成分測定實驗飼料中的粗蛋白、粗脂肪和灰分的測定參照AOAC(2003)標準方法: 采用凱氏定氮儀(2300, Kjeltec Analyzer Unit)測定粗蛋白; 粗脂肪采用索氏抽提儀(Soxtec System HT6, Tecator, Hoganas, Sweden)進行抽提測定; 灰分在馬弗爐中550℃煅燒3h, 采用失重法測定。
腸道黏膜形態測定每缸隨機撈取3尾魚、每組9尾, 置于冰上解剖后分離腸道, 用0.1 mol/L PBS溶液輕輕沖洗內容物后, 再以4%多聚甲醛溶液將前腸部分固定于1.5 mL無RNase EP管中, 并密封保存于4℃冰箱, 用于制作腸道光學顯微鏡切片。腸道黏膜形態樣品參照孫浪等[10]石蠟切片的制作程序(脫水、石蠟包埋、切片和H-E染色)制出切片, 使用光學顯微鏡, 放大40倍觀察并拍照。每張切片隨機選數根平整完好的絨毛, 測量絨毛高度與隱窩深度的比值, 最后把各指標的平均數作為測定數值。
腸道微生物測定每缸隨機撈取2尾魚、每組6尾, 置于冰上解剖后分離腸道, 用0.1 mol/L PBS溶液輕輕沖洗, 收集食糜, 用封口膜封好放入液氮中用于腸道微生物分析。腸道微生物樣品先用DNA提取試劑盒(北京天根生化科技有限公司)提取草魚腸道食糜中DNA。然后將DNA樣品送去人和未來生物科技有限公司(湖南, 長沙)進行16S rDNA測序分析。16S rDNA測序包括細菌基因組DNA提取、16S rDNA特異引物PCR擴增、擴增產物純化、DNA測序、序列比對等步驟, 本實驗基于Illumina Miseq測序平臺對構建的小片段文庫進行雙末端測序。
小肽轉運相關基因表達水平測定每缸隨機撈取3尾魚、每組9尾, 置于冰上解剖后分離腸道,取前腸用剪刀剪碎裝入1.5 mL無RNase EP管中, 迅速投入液氮中后轉移至-80℃冰箱保存, 用于基因表達分析。取凍存的草魚腸道樣品液氮研磨后, 用RNA提取試劑(TaKaRa, RNAiso Plus, Cat. 9109)提取總RNA, 具體方法按說明書進行操作, 分別采用1%瓊脂糖凝膠電泳和全波長讀數儀(Beijing Free More bioscience)檢測RNA的完整性和純度。各取500 ng總RNA作為反轉錄模板, 用反轉錄試劑盒(TaKaRa, Cat.RR036A)進行第一鏈cDNA合成。設計熒光定量引物: 從GenBank中查詢并下載草魚的CDX2、Sp1和PepT1基因的特異序列, 用Primer Premier 5.0軟件設計實時熒光定量PCR (Real-time PCR)特異引物, 內參基因引物根據草魚管家基因βactin設計(表 2)。采用試劑盒(TaKaRa, Cat.RR104A)和BIO-RAD CFX96TM Real-time PCR儀進行Realtime PCR擴增。PCR反應體系為: SYBR Premix ExTaqTM(2×)10 μL, 正向引物(10 μmol/L) 0.5 μL, 反向引物(10 μmol/L) 0.5 μL, 模板0.5 μL, 雙蒸水8.5 μL,總體積為20 μL, 每個樣品重復3次。Real-time PCR反應在Eppendorf Master cycler gradient (Eppendorf, German)中進行, 反應程序: 95℃ 30s, 95℃3s、55℃ 25s、72℃ 11s共40個循環, 95℃ 1min、55℃ 30s、95℃ 30s終止, 4℃保存。Real-time PCR結束后, 從擴增曲線得出Ct值。
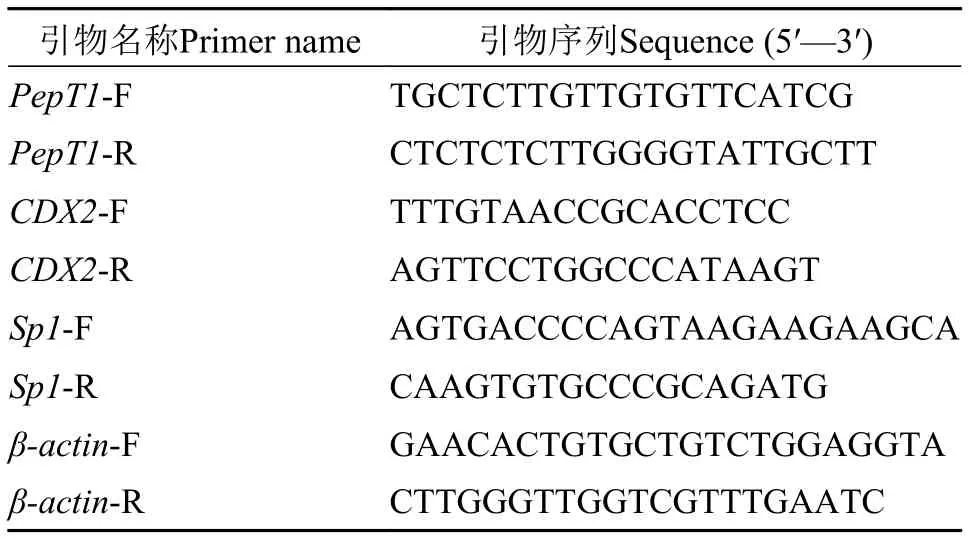
表2 熒光定量分析的引物參數Tab. 2 Primers used for real-time quantitative PCR
1.5 數據處理
實驗以Image-pro plus 6.0軟件分析腸道切片圖, 并計算出絨毛高度/隱窩深度的比值。Real-time PCR數據通過公式RQ=2-ΔΔCt[ΔΔCt=(Ct目的基因-Ct管家基因)-ΔCt校準樣]計算得出目的基因的相對表達量[11]。用Excel 2007進行數據統計, 以SPSS 19.0軟件進行單因素方差分析(One-way ANOVA),所有數據以平均值±標準誤(Means±SE)表示。采用Duncan檢驗進行多重比較, 檢驗不同組間的差異是否顯著, 以P<0.05為顯著性判斷標準。
2 結果
2.1 生長性能
如表 3所示, 各組間草魚增重率、特定生長率和蛋白質效率均無顯著性差異, 實驗1組和2組略高于對照組, 實驗3組略低于對照組(P>0.05); 各組間餌料系數也無顯著性差異, 實驗1組和2組略低于對照組, 實驗3組略高于對照組(P>0.05); 在存活率方面, 四組均為100%。
2.2 腸道形態
用不同比例的發酵芝麻粕替代菜粕的四組草魚前腸黏膜形態在光學顯微鏡下的觀察比較情況如圖 1。表 4展示了發酵芝麻粕替代菜粕對草魚腸道形態的影響: 實驗組的草魚腸絨毛高度均顯著高于對照組(P<0.05)。隨著飼料中發酵芝麻粕替代比例的增加, 草魚腸絨毛高度也隨之增加, 在實驗2組(替代比例為23.5%)時達到最大值, 隨后顯著下降(P<0.05); 實驗組魚腸隱窩深度小于對照組, 但無顯著差異(P>0.05); 實驗組魚腸絨毛高度與隱窩深度的比值(V/C)也均顯著高于對照組(P<0.05), 其變化趨勢與絨毛高度變化趨勢相似。
2.3 腸道微生物
表5展示了發酵芝麻粕替代飼料中菜粕蛋白后, 大腸桿菌(Escherichia coli)、乳酸桿菌(Lactobacillus)、芽孢桿菌(Bacillus)和氣單胞菌(Aeromonas)在草魚腸道總菌群中所占比例的變化情況。與對照組相比, 實驗組大腸桿菌占比顯著下降(P<0.05),實驗組間無顯著差異; 實驗組乳酸桿菌和芽孢桿菌占比均顯著高于對照組(P<0.05), 乳酸桿菌占比隨發酵芝麻粕替代比例的增加而升高, 芽孢桿菌占比在實驗組間無顯著差異(P>0.05); 實驗2組和3組的氣單胞菌占比顯著低于對照組(P<0.05)。
2.4 小肽轉運相關基因mRNA表達量
圖2表示飼料中不同水平發酵芝麻粕對草魚腸道小肽轉運相關基因CDX2、Sp1和PepT1 mRNA相對表達量的影響。這3種基因mRNA的相對表達量均在實驗1組時達到最高, 顯著高于對照組(P<0.05)。如圖所示, 實驗1組的CDX2 mRNA表達水平顯著高于對照組(P<0.05), 其他實驗組也高于對照組, 但無顯著差異(P>0.05)。Sp1 mRNA表達量在實驗1組時顯著高于其他三組(P<0.05), 其余三組間無顯著差異。實驗1組PepT1 mRNA表達量顯著高于對照組和其他實驗組(P<0.05), 實驗2組PepT1 mRNA表達量顯著高于對照組和實驗3組(P<0.05)。
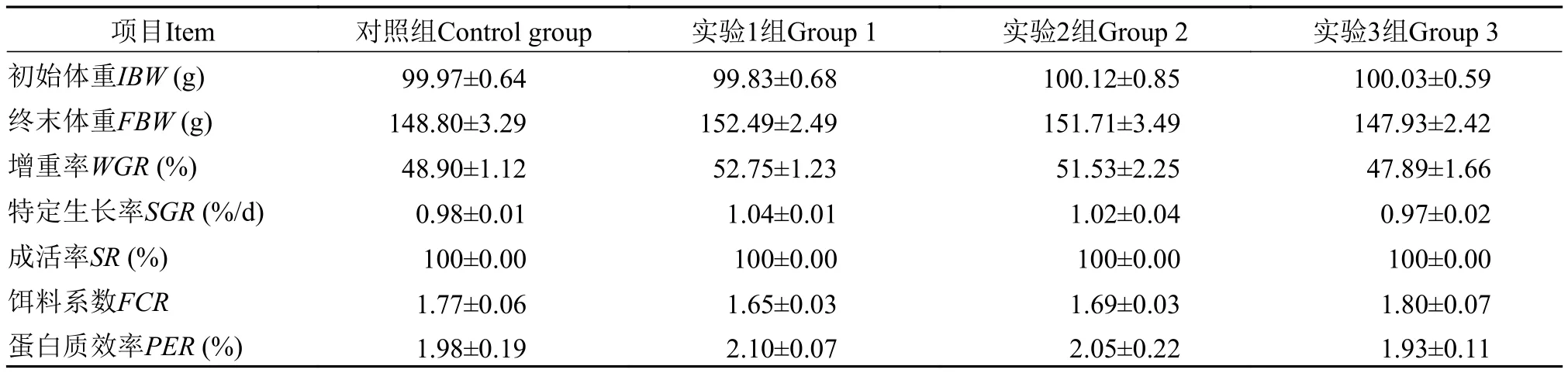
表3 發酵芝麻粕替代飼料中菜粕蛋白對草魚生長性能的影響Tab. 3 Effects of fermented sesame meal replacing dietary rapeseed meal on growth performance of grass carp
3 討論
3.1 發酵芝麻粕替代菜粕蛋白對草魚生長性能的影響
目前, 有關發酵芝麻粕對畜禽和水產動物生長性能影響的研究已有很多。Wu等[1]用發酵芝麻粕替代基礎日糧中豆粕來飼喂櫻桃谷肉鴨, 結果顯示各處理組肉鴨平均日增重和料增重比差異均不顯著, 而以4%—8%發酵芝麻粕替代飼料中豆粕時, 鴨胸肌率和屠宰率顯著高于對照組。Oluwagbenga等[12]研究發現, 在羅非魚(Oreochromis mossambicus)飼料中添加164.3 g/kg發酵芝麻粕時, 對照組與實驗組魚之間的生長性能無顯著性差異。本實驗結果表明, 發酵芝麻粕代替飼料中菜粕對草魚生長有一定的促進作用。發酵芝麻粕代替飼料中菜粕蛋白11.8%和23.5%時, 草魚增重率、特定生長率和蛋白質效率均高于對照組, 餌料系數低于對照組, 但差異不顯著。相似的是, 有研究報道表明, 飼料中使用發酵芝麻粉(未榨油)能促進魚體的生長性能。Mukhopadhyay等[3]發現, 以乳酸菌發酵芝麻粉, 能有效的消除芝麻粉中的主要抗營養因子植酸和單寧。同時, 露斯塔野鯪(Labeo rohita)在攝食含有40%該發酵芝麻粉日糧60d后, 其生長、飼料轉化率和蛋白質效率顯著提高[3]。同樣, Roy 等[13]用從成年露斯塔野鯪腸道中分離出的地衣芽孢桿菌LF1和LH1發酵芝麻粉, 并以30%此發酵芝麻粉飼喂野鯪幼魚, 能顯著提高其生長速度、飼料轉化率和蛋白效率。
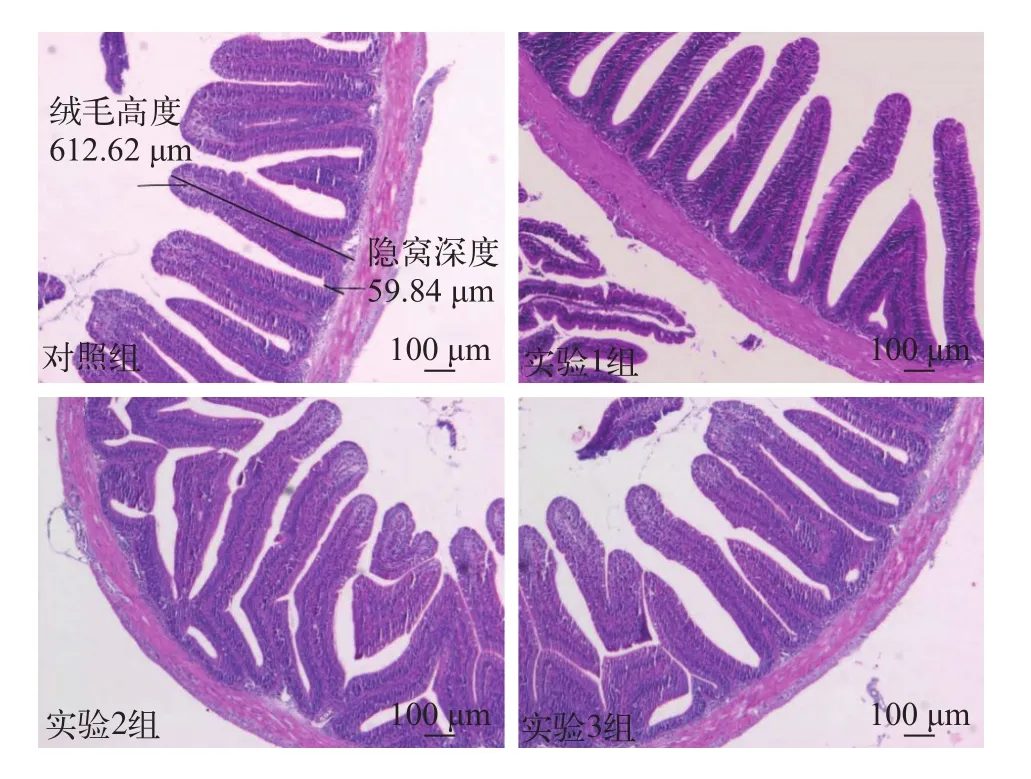
圖1 不同發酵芝麻粕替代組草魚前腸黏膜形態光鏡下比較(×100)Fig. 1 The foregut mucosa morphology of grass carp in different groups under optical microscope (×100)
3.2 發酵芝麻粕替代菜粕蛋白對草魚腸道形態的影響
腸道是動物機體消化吸收營養物質的主要部位, 絨毛高度、隱窩深度以及兩者的比值是反映小腸消化吸收功能的重要指標[14]。腸黏膜上有大量腸絨毛, 絨毛高度越高則腸道對營養物質的吸收面積越大, 消化吸收功能越強; 隱窩深度的高低反映了腸上皮細胞的生成率, 腸道隱窩的細胞不斷的向絨毛處分化, 替代補充脫落或損傷的絨毛細胞, 隱窩處細胞生成率下降將使隱窩深度變淺, 表明絨毛細胞成熟率上升, 小腸營養吸收功能增強; 腸絨毛高度與隱窩深度的比值(V/C)可以比較直觀的反映小腸的消化吸收能力, 絨毛高度與隱窩深度的比值越高則代表小腸消化吸收能力越強[14,15]。本實驗結果表明, 草魚前腸絨毛高度隨著發酵芝麻粕替代比例的增加呈先升高后下降趨勢, 當用發酵芝麻粕替代草魚飼料中23.5%的菜粕蛋白時, 草魚腸絨毛高度顯著高于對照組, 腸絨毛高度與隱窩深度的比值(V/C)也達最大值。這是因為發酵有效地降低了芝麻粕中抗營養因子的含量, 在對照組飼料中因豆粕和菜粕的存在, 從而單寧、植酸等抗營養因子含量過高而損害了草魚的腸道結構[16], 而實驗組中使用發酵芝麻粕替代菜粕, 相應減少了有關抗營養因子, 保護了魚腸道的形態結構[3,13]。

表4 發酵芝麻粕替代飼料中菜粕蛋白對草魚前腸形態的影響Tab. 4 Effects of fermented sesame meal replacing dietary rapeseed meal on foregut morphology of grass carp
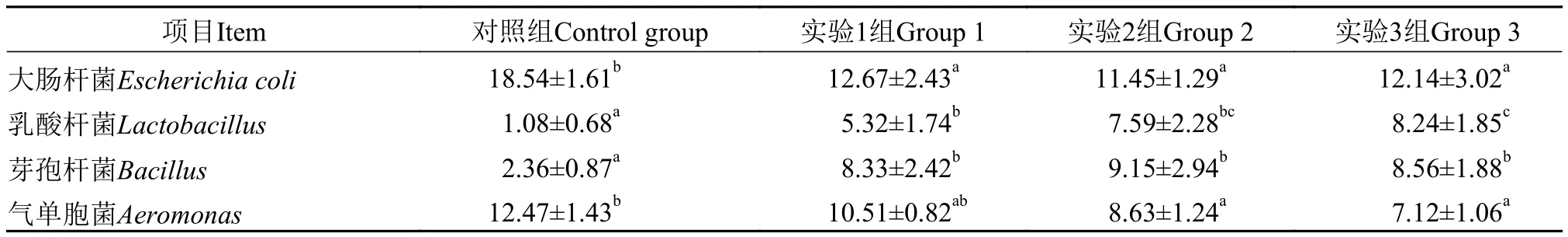
表5 飼料中發酵芝麻粕替代菜粕蛋白對草魚腸道微生物的影響Tab. 5 Effects of fermented sesame meal replacing dietary rapeseed meal on intestinal microflora of grass carp (‰)
3.3 發酵芝麻粕替代菜粕蛋白對草魚腸道微生物的影響
動物腸道微生物種群的穩態與宿主的腸道健康狀況及生長性能密切相關, 腸道微生物種群的變化可直接影響動物腸道對營養物質的吸收代謝[17,18]。飼喂不同餌料會在很大程度上影響腸道菌群結構[19]。本實驗中發酵芝麻粕替代菜粕蛋白增加了草魚腸道中乳酸桿菌和芽孢桿菌的占比, 減少了腸道內氣單胞菌和大腸桿菌的占比。這與夏素銀等[20]用發酵蛋白飼料替代豆粕飼喂肉仔雞, 導致腸道大腸桿菌數數量下降的結果相似。在一定程度上可以說明, 發酵芝麻粕能夠促進腸道中有益菌的增殖并抑制有害細菌的生長, 從而改善腸道微生態, 促進腸道健康。本實驗中腸道菌群的變化可能是由于飼喂芝麻粕經過微生物發酵后, 發酵產生的一些中間產物如肽類和氨基酸等促進了有益微生物混合菌的生長, 主要包括乳酸桿菌、酵母菌和芽孢桿菌,后兩者的存在正好為乳酸桿菌的生長繁殖創造了厭氧環境, 而乳酸桿菌大量繁殖產生的乳酸又降低了腸道中pH, 從而有效抑制了腸道中有害菌如大腸桿菌和氣單胞菌的繁殖, 保持了腸道健康的微生態環境[18, 21, 22]。
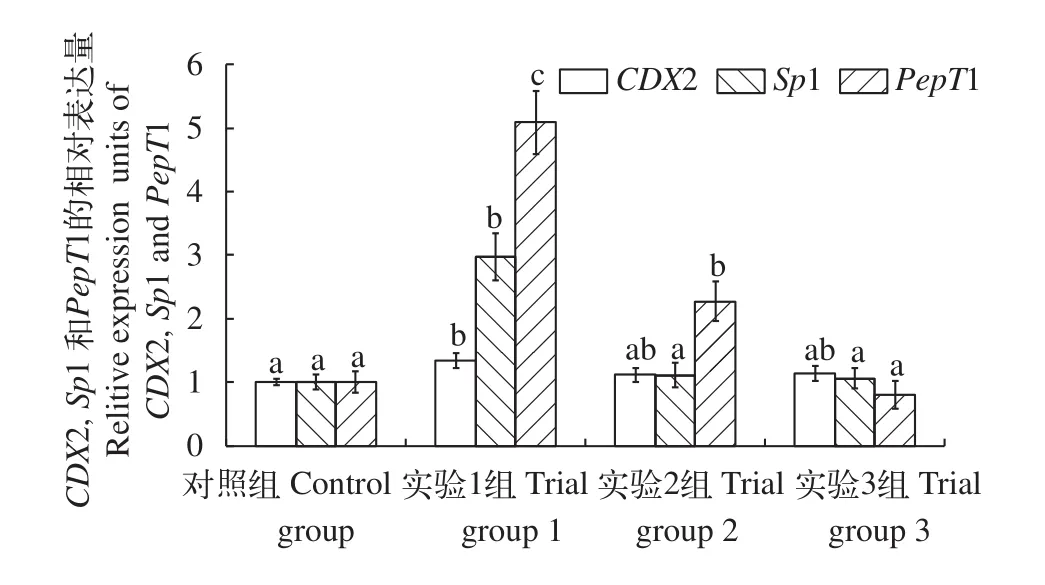
圖2 發酵芝麻粕替代菜粕蛋白對草魚腸道CDX2、Sp1和PepT1 mRNA表達的影響Fig. 2 The effect of fermented sesame meal replacing dietary rapeseed meal on gene expression of CDX2, Sp1 and PepT1 mRNA in intestine of grass crap
3.4 發酵芝麻粕替代菜粕蛋白對草魚腸道小肽轉運相關基因的影響
很多研究都證明蛋白質并不是全部以氨基酸的形式被機體吸收, 有一部分是以小肽的形式被吸收且其吸收效果優于氨基酸的形式[23—25]。動物腸道上皮細胞是小肽吸收的主要場所, 而小肽的吸收是受到多種因素共同調控的[25]。小肽轉運載體PepT1是位于腸道上皮細胞上的一種轉運小肽的蛋白, 它負責將小肽從腸道細胞外轉運到細胞內, 并在動物腸道小肽的吸收過程中發揮關鍵性作用[26,27]。CDX2作為一種腸道上皮細胞特異性核轉錄因子,對營養物質在腸道內消化吸收起著關鍵性作用, 它調控了機體內糖代謝、脂肪代謝、蛋白質代謝和礦物元素代謝等相關功能基因的表達[28,29]。Sp1作為轉錄激活因子或抑制因子在細胞生長、分化、凋亡、染色質重構、纖維化和炎癥等方面起作用[30]。Shimakura等[31]在2005年報道, 轉錄因子CDX2和Sp1能相互作用來調控人類PepT1基因的表達。本實驗用發酵芝麻粕替代菜粕蛋白飼喂草魚, 結果發現小肽轉運相關基因(CDX2、Sp1和PepT1)的表達水平均在飼料中發酵芝麻粕替代11.8%菜粕蛋白時處于最高值, 替代比例為23.5%和35.1%時, 基因表達水平也高于對照組或與對照組沒有顯著差異。這可能是由于微生物發酵將芝麻粕中大量蛋白質降解為了腸道可吸收的小肽, 從而提高了機體小肽轉運相關基因的表達, 進而促進腸道對小肽分子的吸收[1,32]。CDX2、Sp1和PepT1基因表達水平的升高在一定程度上可以說明在飼料中用適量添加發酵芝麻粕替代菜粕有助于小肽的轉運以及氨基酸的消化吸收, 從而有利于草魚的健康和生長。
4 結論
本實驗結果表明, 使用發酵芝麻粕替代草魚飼料中11.8%—23.5%的菜粕蛋白時, 魚體的生長和飼料利用均有上升趨勢; 在飼料中添加適量發酵芝麻粕可改善草魚的腸道形態結構, 優化腸道菌群, 并上調魚腸道中小肽轉運相關基因CDX2、Sp1和PepT1的表達水平。綜上所述, 發酵芝麻粕可以部分替代菜粕用于草魚飼料中。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
